
The Loss-Function of KNL1 Causes Oligospermia and Asthenospermia in Mice by Affecting the Assembly and Separation of the Spindle through Flow Cytometry and Immunofluorescence

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Mice Culture
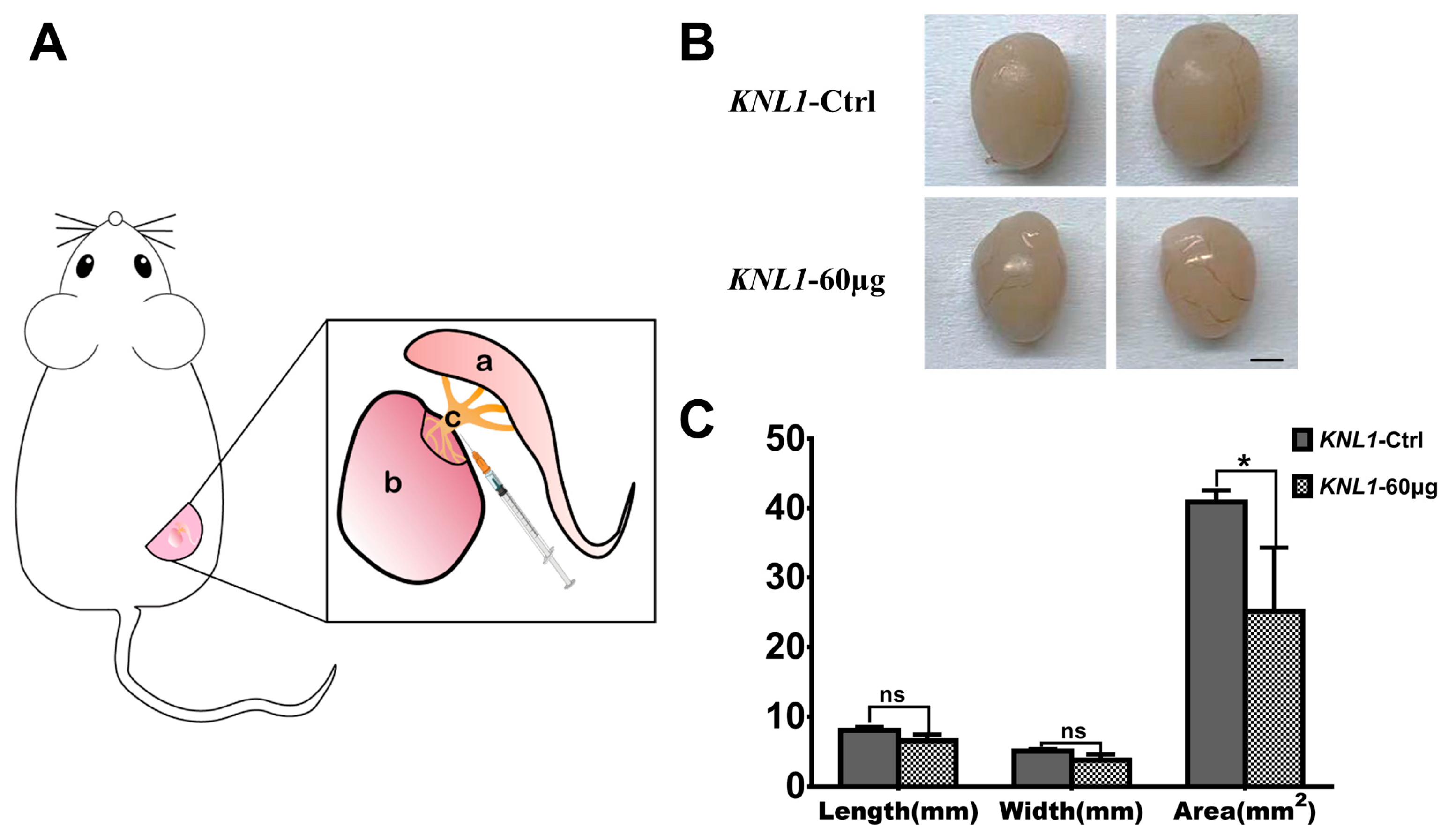
2.3. Seminiferous Tubule Injection Experiment
2.4. Sample Processing for HE Staining and CASA
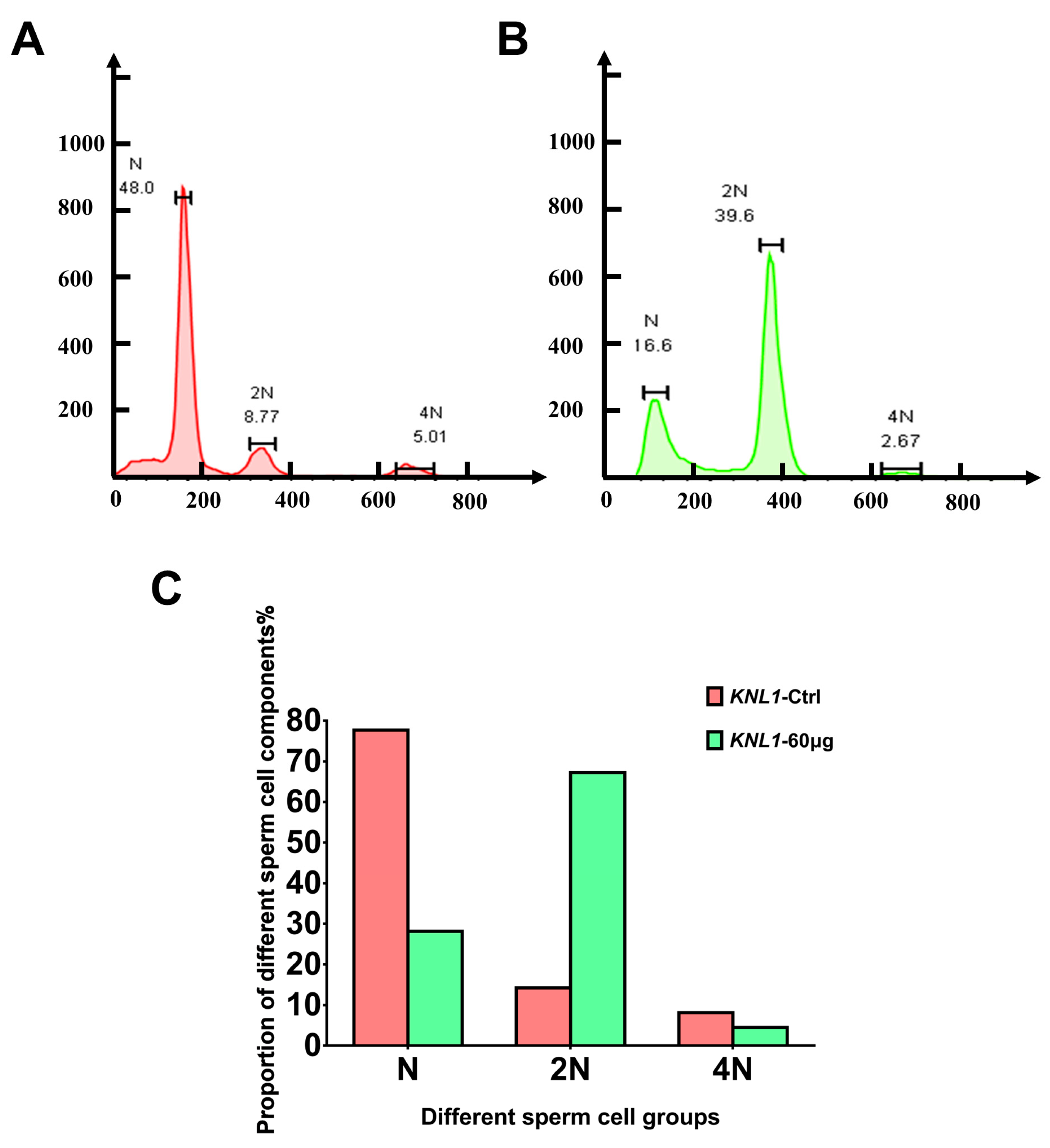
2.5. Flow Cytometry Experiment
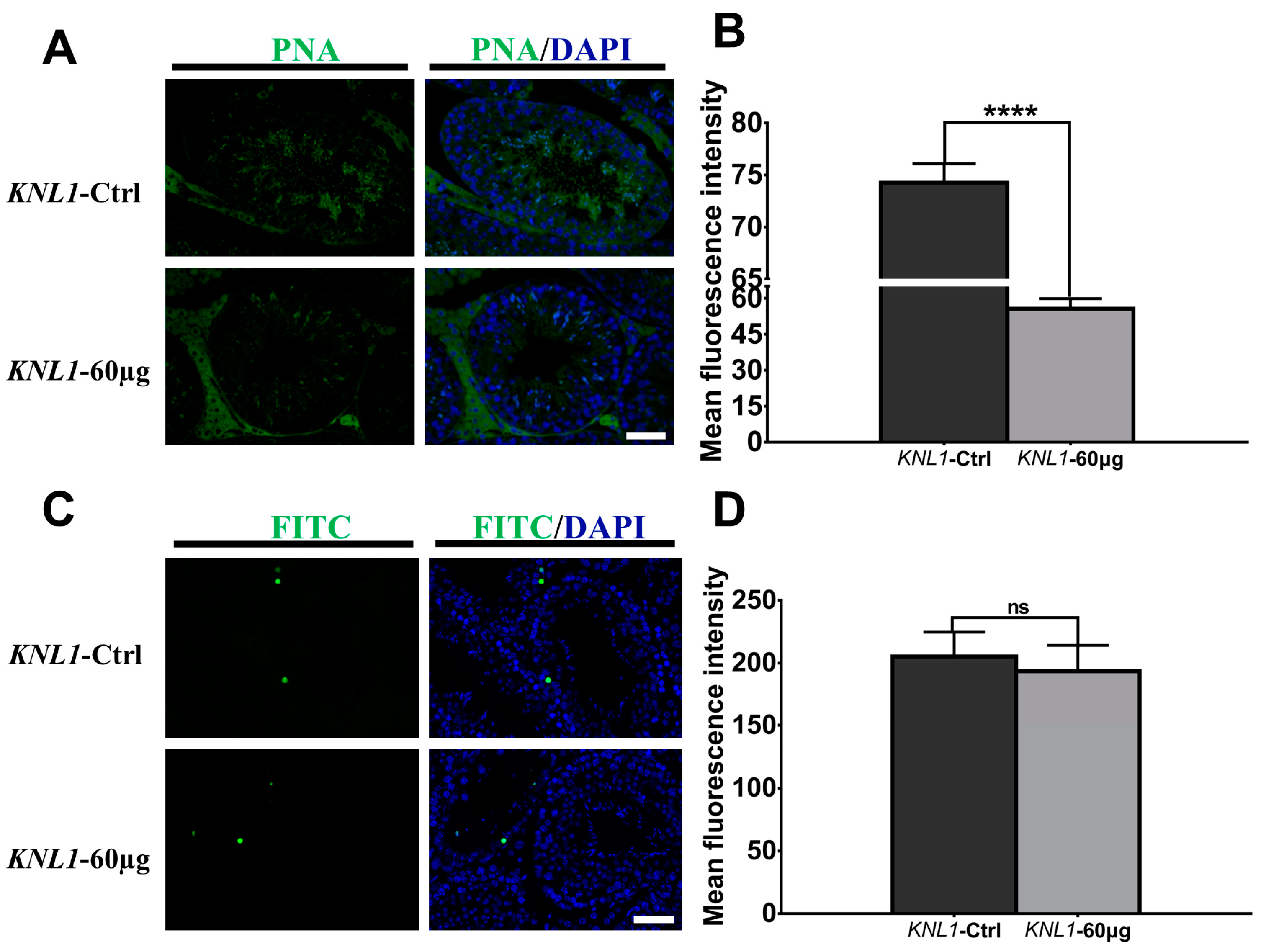
2.6. TUNEL and PNA Method
2.7. Immunofluorescence Staining
2.8. siRNA Transfection and Synchronization
2.9. siRNA Sequence and Immunofluorescence Antibody
3. Results
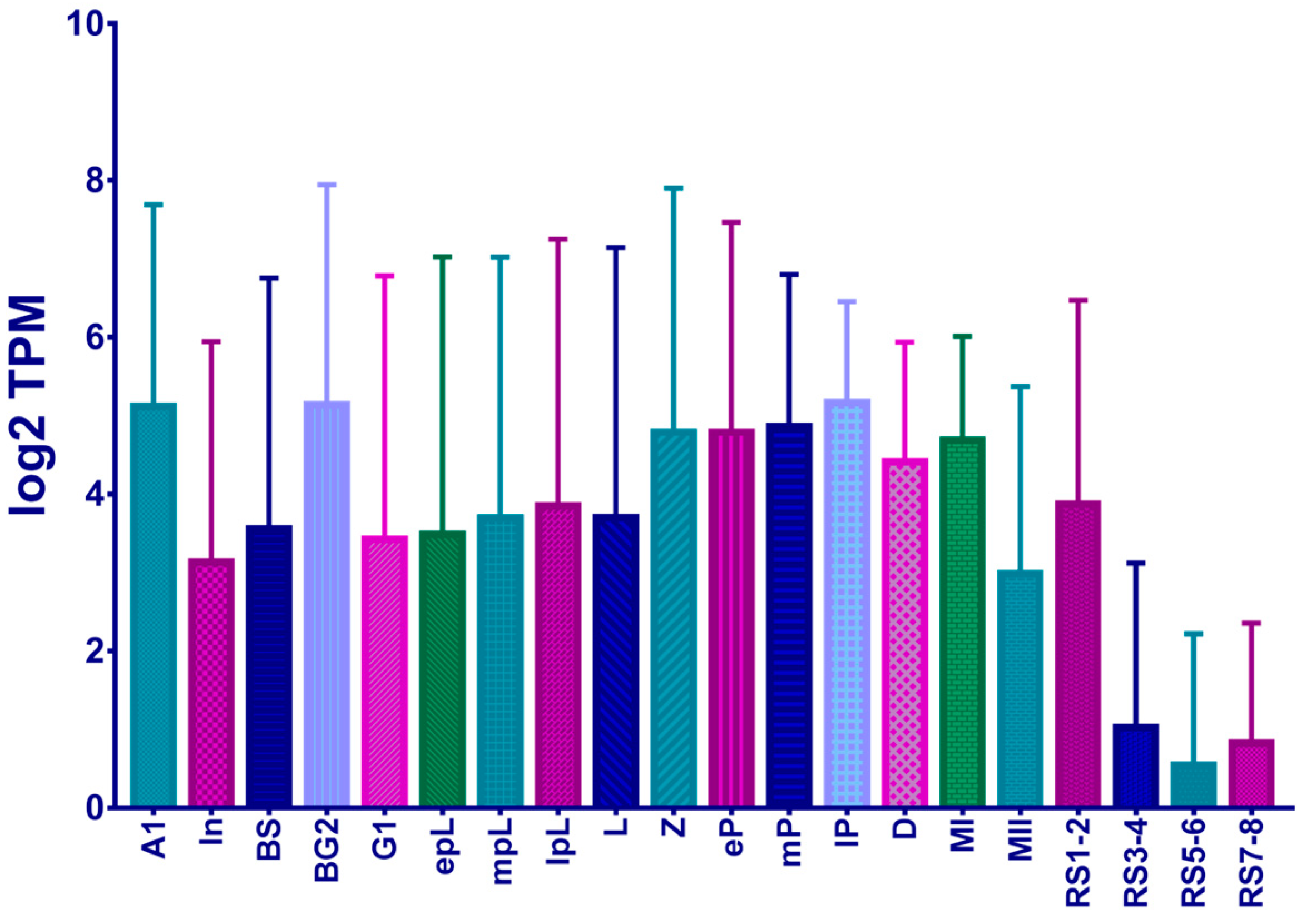
3.1. The Loss-Function of the KNL1 Gene Influences Male Reproductive Phenotype in Mice
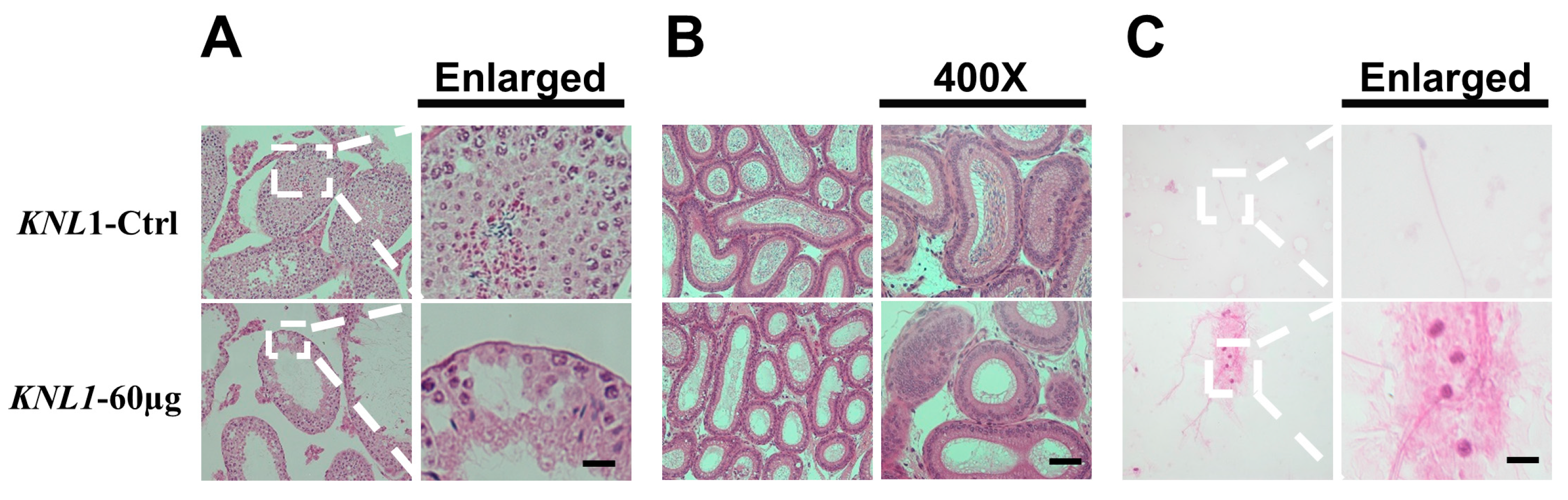
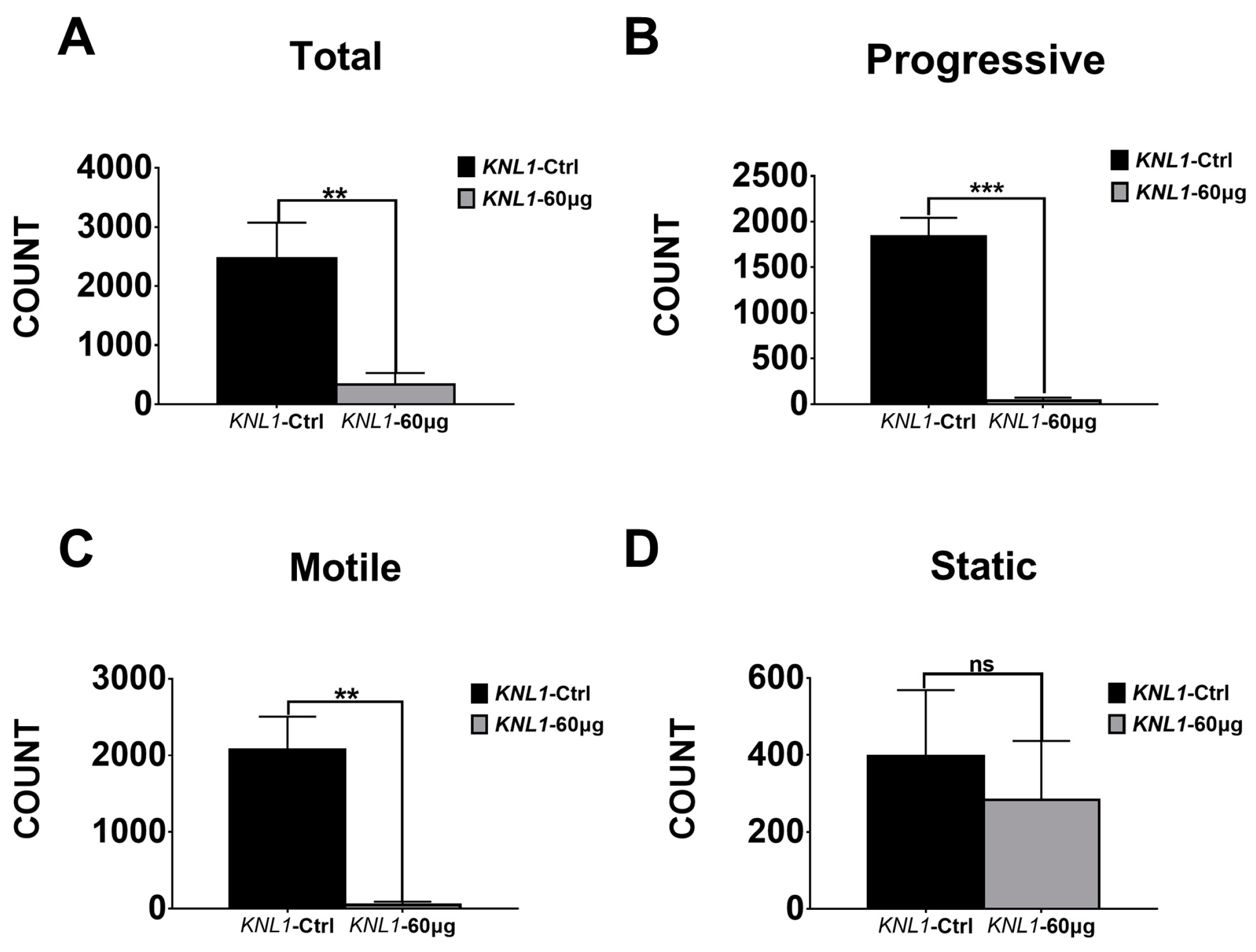
3.2. The Loss-Function of the KNL1 Gene Results in Oligospermia and Asthenospermia in Mice
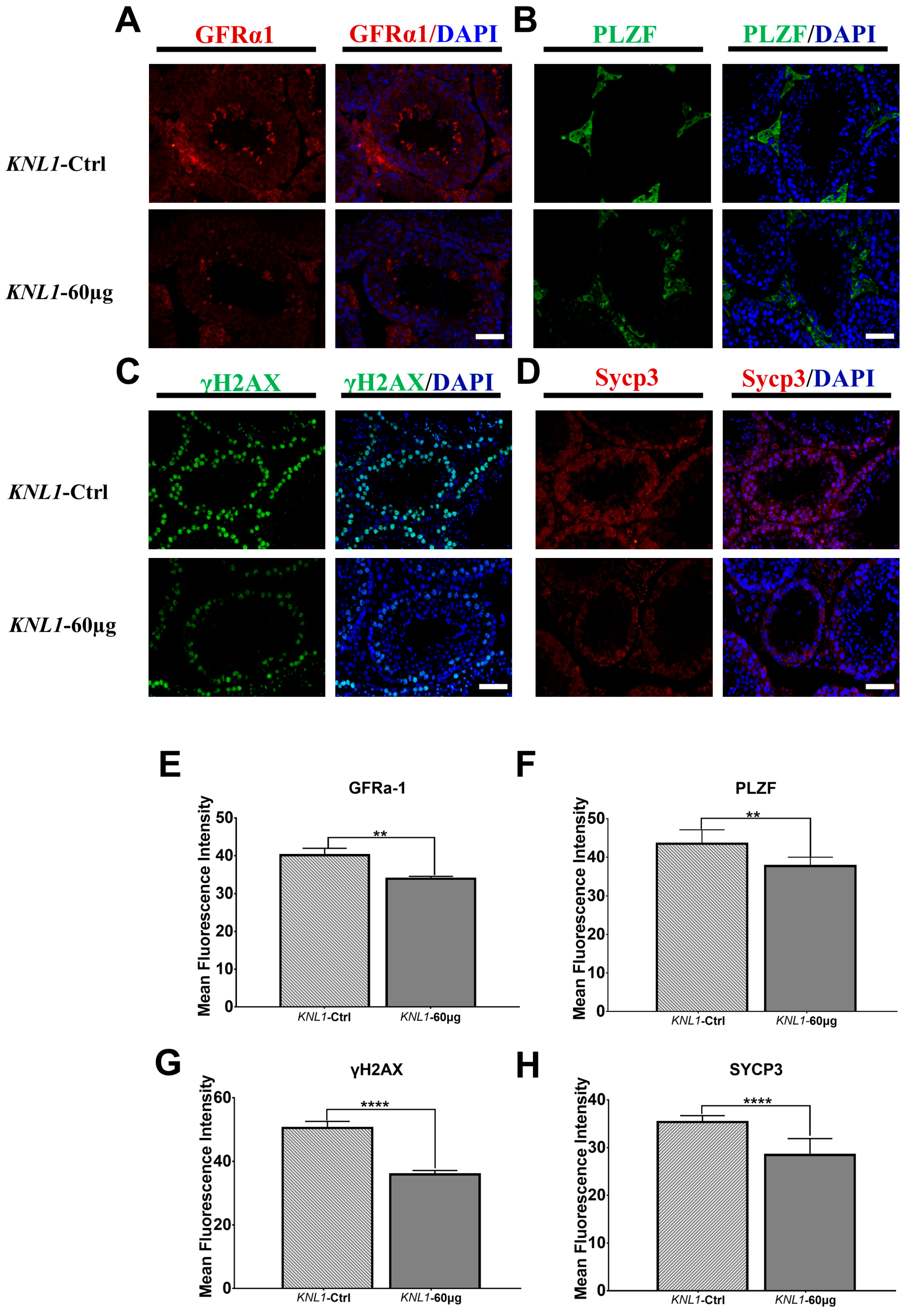
3.3. Loss-Function of KNL1 Causes Spermatocytes Arrest at the Meiotic Prophase I of Spermatogenesis
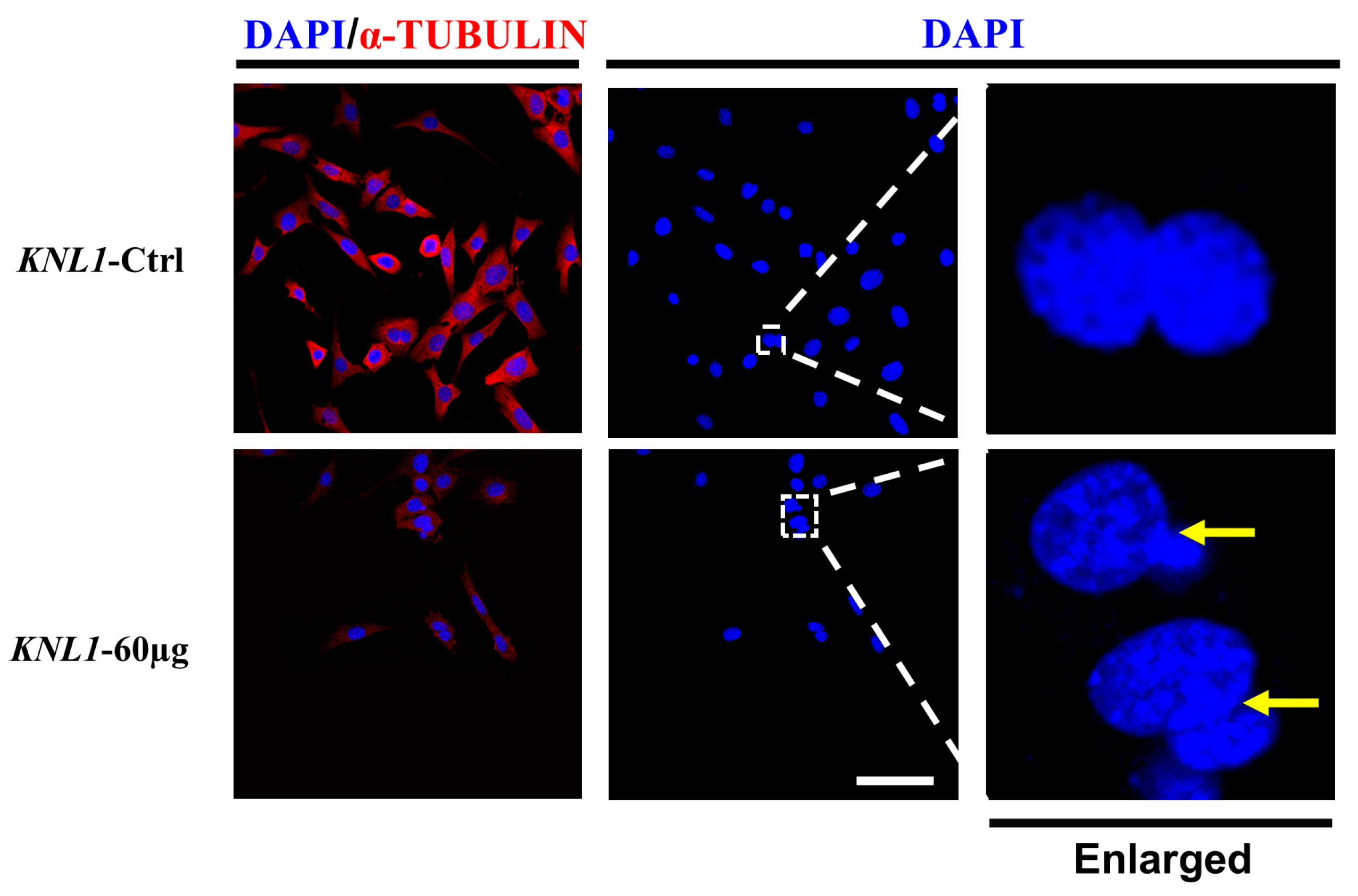
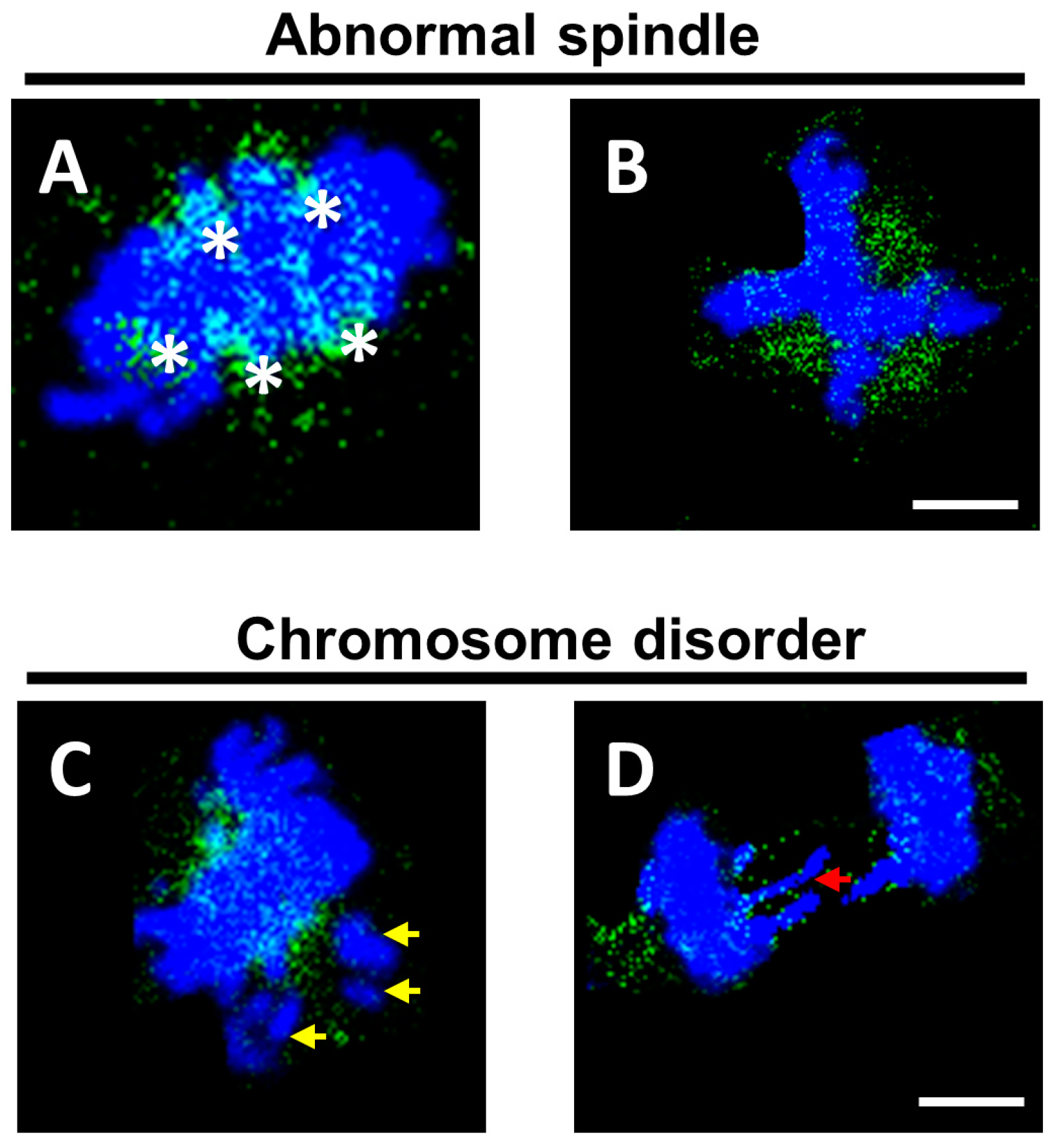
3.4. The Loss-Function of KNL1 Leads to Abnormal Assembly and Separation of the Spindle Resulting in Unequal Cell Segregation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mora-Bermúdez, F.; Kanis, P.; Macak, D.; Peters, J.; Naumann, R.; Xing, L.; Sarov, M.; Winkler, S.; Oegema, C.E.; Haffner, C.; et al. Longer metaphase and fewer chromosome segregation errors in modern human than Neanderthal brain development. Sci. Adv. 2022, 8, eabn7702. [Google Scholar] [CrossRef]
- Lara-Gonzalez, P.; Pines, J.; Desai, A. Spindle assembly checkpoint activation and silencing at kinetochores. Semin. Cell Dev. Biol. 2021, 117, 86–98. [Google Scholar] [CrossRef]
- Claeys Bouuaert, C.; Pu, S.; Wang, J.; Oger, C.; Daccache, D.; Xie, W.; Patel, D.J.; Keeney, S. DNA-driven condensation assembles the meiotic DNA break machinery. Nature 2021, 592, 144–149. [Google Scholar] [CrossRef]
- Stiff, T.; Echegaray-Iturra, F.R.; Pink, H.J.; Herbert, A.; Reyes-Aldasoro, C.C.; Hochegger, H. Prophase-Specific Perinuclear Actin Coordinates Centrosome Separation and Positioning to Ensure Accurate Chromosome Segregation. Cell Rep. 2020, 31, 107681. [Google Scholar] [CrossRef]
- Hinshaw, S.M.; Harrison, S.C. Kinetochore Function from the Bottom Up. Trends Cell Biol. 2018, 28, 22–33. [Google Scholar] [CrossRef]
- Ghodgaonkar-Steger, M.; Potocnjak, M.; Zimniak, T.; Fischböck-Halwachs, J.; Solis-Mezarino, V.; Singh, S.; Speljko, T.; Hagemann, G.; Drexler, D.J.; Witte, G.; et al. C-Terminal Motifs of the MTW1 Complex Cooperatively Stabilize Outer Kinetochore Assembly in Budding Yeast. Cell Rep. 2020, 32, 108190. [Google Scholar] [CrossRef]
- Musacchio, A.; Desai, A. A Molecular View of Kinetochore Assembly and Function. Biology 2017, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, I.M. The Kinetochore. Cold Spring Harb. Perspect. Biol. 2014, 6, a015826. [Google Scholar] [CrossRef]
- Vleugel, M.; Tromer, E.; Omerzu, M.; Groenewold, V.; Nijenhuis, W.; Snel, B.; Kops, G.J.P.L. Arrayed BUB recruitment modules in the kinetochore scaffold KNL1 promote accurate chromosome segregation. J. Cell Biol. 2013, 203, 943–955. [Google Scholar] [CrossRef]
- Hara, M.; Fukagawa, T. Dynamics of kinetochore structure and its regulations during mitotic progression. Cell Mol. Life Sci. 2020, 77, 2981–2995. [Google Scholar] [CrossRef]
- Cheeseman, I.M.; Chappie, J.S.; Wilson-Kubalek, E.M.; Desai, A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell 2006, 127, 983–997. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, I.M.; Hori, T.; Fukagawa, T.; Desai, A. KNL1 and the CENP-H/I/K complex coordinately direct kinetochore assembly in vertebrates. Mol. Biol. Cell 2008, 19, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Bock, L.J.; Pagliuca, C.; Kobayashi, N.; Grove, R.A.; Oku, Y.; Shrestha, K.; Alfieri, C.; Golfieri, C.; Oldani, A.; Dal Maschio, M.; et al. Cnn1 inhibits the interactions between the KMN complexes of the yeast kinetochore. Nat. Cell Biol. 2012, 14, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.W.; Jeyaprakash, A.A.; Nigg, E.A.; Santamaria, A. Aurora B controls kinetochore-microtubule attachments by inhibiting Ska complex-KMN network interaction. J. Cell Biol. 2012, 196, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Liu, Y.; Wang, C.; Liu, Y.; Feng, C.; Sun, Y.; Yuan, J.; Birchler, J.A.; Han, F. Knl1 participates in spindle assembly checkpoint signaling in maize. Proc. Natl. Acad. Sci. USA 2021, 118, e2022357118. [Google Scholar] [CrossRef]
- Caldas, G.V.; DeLuca, J.G. KNL1: Bringing order to the kinetochore. Chromosoma 2014, 123, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Krenn, V.; Overlack, K.; Primorac, I.; van Gerwen, S.; Musacchio, A. KI Motifs of Human Knl1 Enhance Assembly of Comprehensive Spindle Checkpoint Complexes around MELT Repeats. Curr. Biol. 2014, 24, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, A.; Mosalaganti, S.; Keller, J.; Mattiuzzo, M.; Overlack, K.; Krenn, V.; De Antoni, A.; Wohlgemuth, S.; Cecatiello, V.; Pasqualato, S.; et al. Modular Assembly of RWD Domains on the Mis12 Complex Underlies Outer Kinetochore Organization. Mol. Cell 2014, 53, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Silio, V.; McAinsh, A.D.; Millar, J.B. KNL1-Bubs and RZZ Provide Two Separable Pathways for Checkpoint Activation at Human Kinetochores. Dev. Cell 2015, 35, 600–613. [Google Scholar] [CrossRef] [Green Version]
- Caldas, G.V.; DeLuca, K.F.; DeLuca, J.G. KNL1 facilitates phosphorylation of outer kinetochore proteins by promoting Aurora B kinase activity. Mol. Biol. Cell 2013, 203, 957–969. [Google Scholar] [CrossRef]
- Petrovic, A.; Keller, J.; Liu, Y.; Overlack, K.; John, J.; Dimitrova, Y.N.; Jenni, S.; van Gerwen, S.; Stege, P.; Wohlgemuth, S.; et al. Structure of the MIS12 Complex and Molecular Basis of Its Interaction with CENP-C at Human Kinetochores. Cell 2016, 167, 1028–1040.e1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ouyang, X.; Su, X. The structure and function of kinetochore proteins KNL1 and its disease correlation. Chem. Life 2019, 39, 1147–1152. [Google Scholar]
- Bai, T.; Zhao, Y.; Liu, Y.; Cai, B.; Dong, N.; Li, B. Effect of KNL1 on the proliferation and apoptosis of colorectal cancer cells. Technol. Cancer Res. Treat. 2019, 18, 1533033819858668. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Qalieh, A.; Lam, M.M.; Keil, J.M.; Kwan, K.Y. Robust elimination of genome-damaged cells safeguards against brain somatic aneuploidy following Knl1 deletion. Nat. Commun. 2019, 10, 2588. [Google Scholar] [CrossRef] [Green Version]
- Saadi, A.; Verny, F.; Siquier-Pernet, K.; Bole-Feysot, C.; Nitschke, P.; Munnich, A.; Abada-Dendib, M.; Chaouch, M.; Abramowicz, M.; Colleaux, L. Refining the phenotype associated with CASC5 mutation. Neurogenetics 2016, 17, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Wang, Y.; Meng, T.G.; Zhang, H.Y.; Zhang, X.R.; Ouyang, Y.C.; Hou, Y.; Schatten, H.; Wang, Z.B.; Sun, Q.Y. Kinetochore scaffold 1 regulates SAC function during mouse oocyte meiotic maturation. FASEB J. 2022, 36, e22210. [Google Scholar] [CrossRef]
- Otsu, M.; Tanabe, Y.; Iwakiri, A.; Arima, K.; Uchiyama, A.; Yamamoto, M.; Ohtani, S.; Endo, H.; Komoto, M.; Miyazaki, K. A report on a modified protocol for flow cytometry-based assessment of blood group erythrocyte antigens potentially suitable for analysis of weak ABO subgroups. Transfusion 2023. [Google Scholar] [CrossRef]
- Ding, L.; Zheng, Q.; Lin, Y.; Wang, R.; Wang, H.; Luo, W.; Lu, Z.; Xie, H.; Ren, L.; Lu, H.; et al. Exosome-derived circTFDP2 promotes prostate cancer progression by preventing PARP1 from caspase-3-dependent cleavage. Clin. Transl. Med. 2023, 13, e1156. [Google Scholar] [CrossRef]
- Peng, B.Y.; Singh, A.K.; Chan, C.H.; Deng, Y.H.; Li, P.Y.; Su, C.W.; Wu, C.Y.; Deng, W.P. AGA induces sub-G1 cell cycle arrest and apoptosis in human colon cancer cells through p53-independent/p53-dependent pathway. BMC Cancer 2023, 23, 1. [Google Scholar] [CrossRef]
- Hara-Yokoyama, M.; Kurihara, H.; Ichinose, S.; Matsuda, H.; Ichinose, S.; Kurosawa, M.; Tada, N.; Iwahara, C.; Terasawa, K.; Podyma-Inoue, K.A.; et al. KIF11 as a Potential Marker of Spermatogenesis Within Mouse Seminiferous Tubule Cross-sections. J. Histochem. Cytochem. 2019, 67, 813–824. [Google Scholar] [CrossRef]
- Im, K.; Mareninov, S.; Diaz, M.F.P.; Yong, W.H. An Introduction to Performing Immunofluorescence Staining. Methods Mol. Biol. 2019, 1897, 299–311. [Google Scholar] [CrossRef]
- Xiong, X.; Zhang, Y.; Xiong, Y.; Yan, M.; Wu, J.; Hao, Z.; Zhang, H.; Li, J. Study on the establishment of a sorting system for yak X and Y sperm by flow cytometry. J. Anim. Sci. Vet. Med. 2021, 52, 399–407. [Google Scholar]
- Llavanera, M.; Mislei, B.; Blanco-Prieto, O.; Baldassarro, V.A.; Mateo-Otero, Y.; Spinaci, M.; Yeste, M.; Bucci, D. Assessment of sperm mitochondrial activity by flow cytometry and fluorescent microscopy: A comparative study of mitochondrial fluorescent probes in bovine spermatozoa. Reprod. Fertil. Dev. 2022, 34, 679–688. [Google Scholar] [CrossRef]
- Da Costa, R.; Redmann, K.; Schlatt, S. Simultaneous detection of sperm membrane integrity and DNA fragmentation by flow cytometry: A novel and rapid tool for sperm analysis. Andrology 2021, 9, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Purdy, P.H.; Graham, J.K.; Azevedo, H.C. Evaluation of boar and bull sperm capacitation and the acrosome reaction using flow cytometry. Anim. Reprod. Sci. 2022, 246, 106846. [Google Scholar] [CrossRef] [PubMed]
- Bulkeley, E.; Santistevan, A.C.; Varner, D.; Meyers, S. Imaging flow cytometry to characterize the relationship between abnormal sperm morphologies and reactive oxygen species in stallion sperm. Reprod. Domest. Anim. 2023, 58, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, Y.X.; Gao, Y.; Lin, Z.; Yang, S.M.; Wang, T.T.; Wang, Q.; Xie, N.N.; Hua, R.; Liu, M.X.; et al. Single-cell RNA-seq uncovers dynamic processes and critical regulators in mouse spermatogenesis. Cell Res. 2018, 28, 879–896. [Google Scholar] [CrossRef] [Green Version]
- Karna, K.K.; Soni, K.K.; You, J.H.; Choi, N.Y.; Kim, H.K.; Kim, C.Y.; Lee, S.W.; Shin, Y.S.; Park, J.K. MOTILIPERM Ameliorates Immobilization Stress-Induced Testicular Dysfunction via Inhibition of Oxidative Stress and Modulation of the Nrf2/HO-1 Pathway in SD Rats. Int. J. Mol. Sci. 2020, 21, 4750. [Google Scholar] [CrossRef]
- Li, G.; Zhang, P.; You, Y.; Chen, D.; Cai, J.; Ma, Z.; Huang, X.; Chang, D. Qiangjing Tablets Regulate Apoptosis and Oxidative Stress via Keap/Nrf2 Pathway to Improve the Reproductive Function in Asthenospermia Rats. Front. Pharmacol. 2021, 12, 714892. [Google Scholar] [CrossRef]
- Zhang, P.H.; Chen, D.A.; Dong, L.; Li, G.S.; Yin, J.; Qu, X.W.; You, Y.D.; Chang, D.G. Inhibitory effect of Qiangjing Tablets on the Fas/FasL pathway of cell apoptosis in male SD rats with infertility. Zhonghua Nan Ke Xue 2016, 22, 246–251. [Google Scholar]
- Hong, T.K.; Song, J.H.; Lee, S.B.; Do, J.T. Germ Cell Derivation from Pluripotent Stem Cells for Understanding In Vitro Gametogenesis. Cells 2021, 10, 1889. [Google Scholar] [CrossRef]
- Wu, J.X.; Xia, T.; She, L.P.; Lin, S.; Luo, X.M. Stem Cell Therapies for Human Infertility: Advantages and Challenges. Cell Transplant. 2022, 31, 9636897221083252. [Google Scholar] [CrossRef] [PubMed]
- Sheikhansari, G.; Aghebati-Maleki, L.; Nouri, M.; Jadidi-Niaragh, F.; Yousefi, M. Current approaches for the treatment of premature ovarian failure with stem cell therapy. Biomed. Pharmacother. 2018, 102, 254–262. [Google Scholar] [CrossRef]
- Leaver, R.B. Male infertility: An overview of causes and treatment options. Br. J. Nurs. 2016, 25, S35–S40. [Google Scholar] [CrossRef]
- Cheeseman, I.M.; Niessen, S.; Anderson, S.; Hyndman, F.; Yates, J.R., 3rd; Oegema, K.; Desai, A. A conserved protein network controls assembly of the outer kinetochore and its ability to sustain tension. Genes Dev. 2004, 18, 2255–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Lischetti, T.; Nilsson, J. A minimal number of MELT repeats supports all the functions of KNL1 in chromosome segregation. J. Cell Sci. 2014, 127, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Krenn, V.; Wehenkel, A.; Li, X.; Santaguida, S.; Musacchio, A. Structural analysis reveals features of the spindle checkpoint kinase Bub1-kinetochore subunit Knl1 interaction. J. Cell Biol. 2012, 196, 451–467. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, A.; Pasqualato, S.; Dube, P.; Krenn, V.; Santaguida, S.; Cittaro, D.; Monzani, S.; Massimiliano, L.; Keller, J.; Tarricone, A.; et al. The MIS12 complex is a protein interaction hub for outer kinetochore assembly. J. Cell Biol. 2010, 190, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Pan, Q.-Y.; Wang, Y.-J.; Chen, Q.-F. Impact of KMN network genes on progression and prognosis of non-small cell lung cancer. Anti-Cancer Drugs 2022, 33, E398–E408. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, C.; Ma, S.; Guo, W.; Cao, W.; Guan, F. CASC5 is a potential tumour driving gene in lung adenocarcinoma. Cell Biochem. Funct. 2020, 38, 733–742. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Q.; Li, L.; Lu, J. Establishment of a Mouse Asthenospermia Model through Triggering D-Galactose Mediated Oxidative Stress Injury. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tu, C.; Wang, L.; Wu, H.; Houston, B.J.; Mastrorosa, F.K.; Zhang, W.; Shen, Y.; Wang, J.; Tian, S.; et al. Deleterious variants in X-linked CFAP47 induce asthenoteratozoospermia and primary male infertility. Am. J. Hum. Genet. 2021, 108, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Liu, H.; Zhang, M.; Zhang, S.; Hu, J.; Wu, G.; Yang, J. Taurine Increases Spermatozoa Quality and Function in Asthenospermia Rats Impaired by Ornidazole. Adv. Exp. Med. Biol. 2019, 1155, 507–520. [Google Scholar] [CrossRef]
- Sha, Y.; Liu, W.; Huang, X.; Li, Y.; Ji, Z.; Mei, L.; Lin, S.; Kong, S.; Lu, J.; Kong, L.; et al. EIF4G1 is a novel candidate gene associated with severe asthenozoospermia. Mol. Genet. Genom. Med. 2019, 7, e807. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Researchers | Year | Methods | Machine Type | Detection |
|---|---|---|---|---|
| Miki Hara-Yokoyama et al. [30] | 2019 | FCM + PI | FACSCalibur | The proportion of haploid, diploid, and tetraploid cells during spermatogenesis |
| Xianrong Xiong et al. [32] | 2021 | FCM + Hoechst33342 + Allura Red | MoFlo XDP | Sperm X and Y were sorted out |
| Marc et al. [33] | 2022 | FCM + Simultaneous co-staining (CMA3 and Yo-Pro-1) | CytoFLEX cytometer | Sperm quality parameters (morphology, viability, total and progressive motility) |
| Raul et al. [34] | 2021 | The co-staining LD + AO | CytoFLEX S | Sperm membrane integrity and DNA fragmentation |
| Phillip et al. [35] | 2022 | FCM + a variety of fluorescent dye (PTYR, PDK, FITC-PNA, PI, M540 and Yo-Pro-1) | CYAN-ADP | Sperm capacitation and functions |
| Evelyn et al. [36] | 2023 | FCM + co-staining SytoxGreen™ and dihydroethidium (DHE) | Amnis® ImageStream® | Certain morphologic abnormalities and ROS |
| Count | KNL1-Ctrl Group | KNL1 Group | ||||
|---|---|---|---|---|---|---|
| Total | 3164 | 2071 | 2180 | 109 | 419 | 471 |
| Motile | 2569 | 1776 | 1878 | 0 | 74 | 75 |
| Progressive | 2067 | 1676 | 1771 | 0 | 62 | 54 |
| Rapid | 2496 | 1768 | 1875 | 0 | 72 | 75 |
| Slow | 73 | 8 | 3 | 0 | 2 | 0 |
| Static | 595 | 295 | 302 | 109 | 345 | 396 |
| Percentage | KNL1-Ctrl Group (%) | KNL1 Group (%) | ||||
|---|---|---|---|---|---|---|
| Motile | 81.2 | 85.8 | 86.1 | 0 | 17.7 | 15.9 |
| Progressive | 65.3 | 80.9 | 81.2 | 0 | 14.8 | 11.4 |
| Rapid | 78.9 | 85.4 | 86 | 0 | 17.2 | 15.9 |
| Slow | 2.3 | 0.3 | 0.1 | 0 | 0.4 | 0 |
| Static | 18.8 | 14.2 | 13.9 | 100 | 82.3 | 84.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yang, J.; Lu, D.; Zhu, Y.; Liao, K.; Tian, Y.; Yin, R. The Loss-Function of KNL1 Causes Oligospermia and Asthenospermia in Mice by Affecting the Assembly and Separation of the Spindle through Flow Cytometry and Immunofluorescence. Sensors 2023, 23, 2571. https://doi.org/10.3390/s23052571
Zhao Y, Yang J, Lu D, Zhu Y, Liao K, Tian Y, Yin R. The Loss-Function of KNL1 Causes Oligospermia and Asthenospermia in Mice by Affecting the Assembly and Separation of the Spindle through Flow Cytometry and Immunofluorescence. Sensors. 2023; 23(5):2571. https://doi.org/10.3390/s23052571
Chicago/Turabian StyleZhao, Yuwei, Jingmin Yang, Daru Lu, Yijian Zhu, Kai Liao, Yafei Tian, and Rui Yin. 2023. "The Loss-Function of KNL1 Causes Oligospermia and Asthenospermia in Mice by Affecting the Assembly and Separation of the Spindle through Flow Cytometry and Immunofluorescence" Sensors 23, no. 5: 2571. https://doi.org/10.3390/s23052571
APA StyleZhao, Y., Yang, J., Lu, D., Zhu, Y., Liao, K., Tian, Y., & Yin, R. (2023). The Loss-Function of KNL1 Causes Oligospermia and Asthenospermia in Mice by Affecting the Assembly and Separation of the Spindle through Flow Cytometry and Immunofluorescence. Sensors, 23(5), 2571. https://doi.org/10.3390/s23052571