Sugar Matters: Improving In Vivo Clearance Rate of Highly Glycosylated Recombinant Plasma Proteins for Therapeutic Use

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
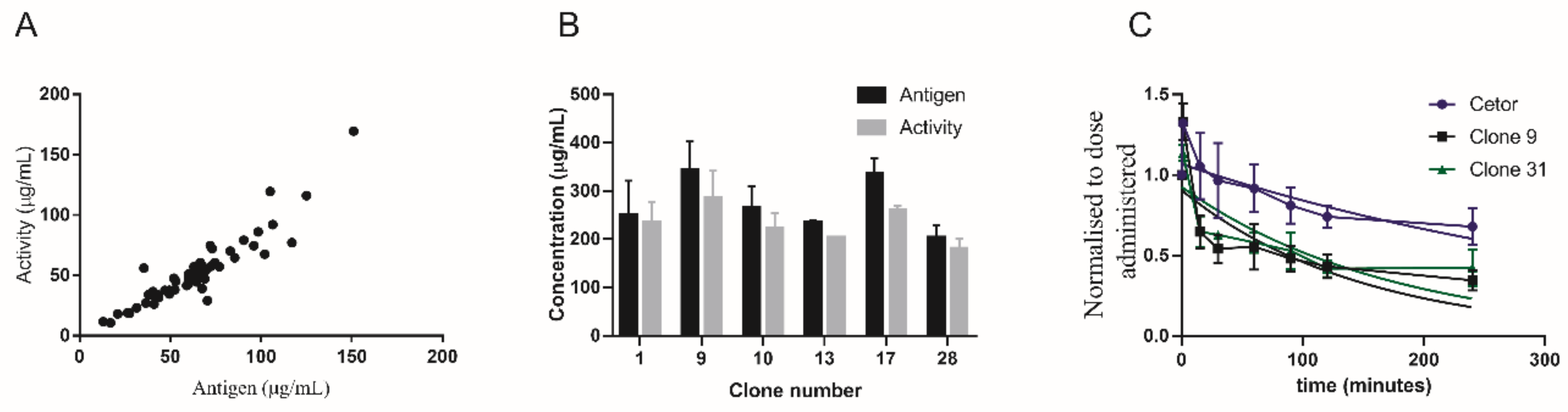
2.1. rC1-INH Is Cleared Rapidly in a Biphasic Manner In Vivo
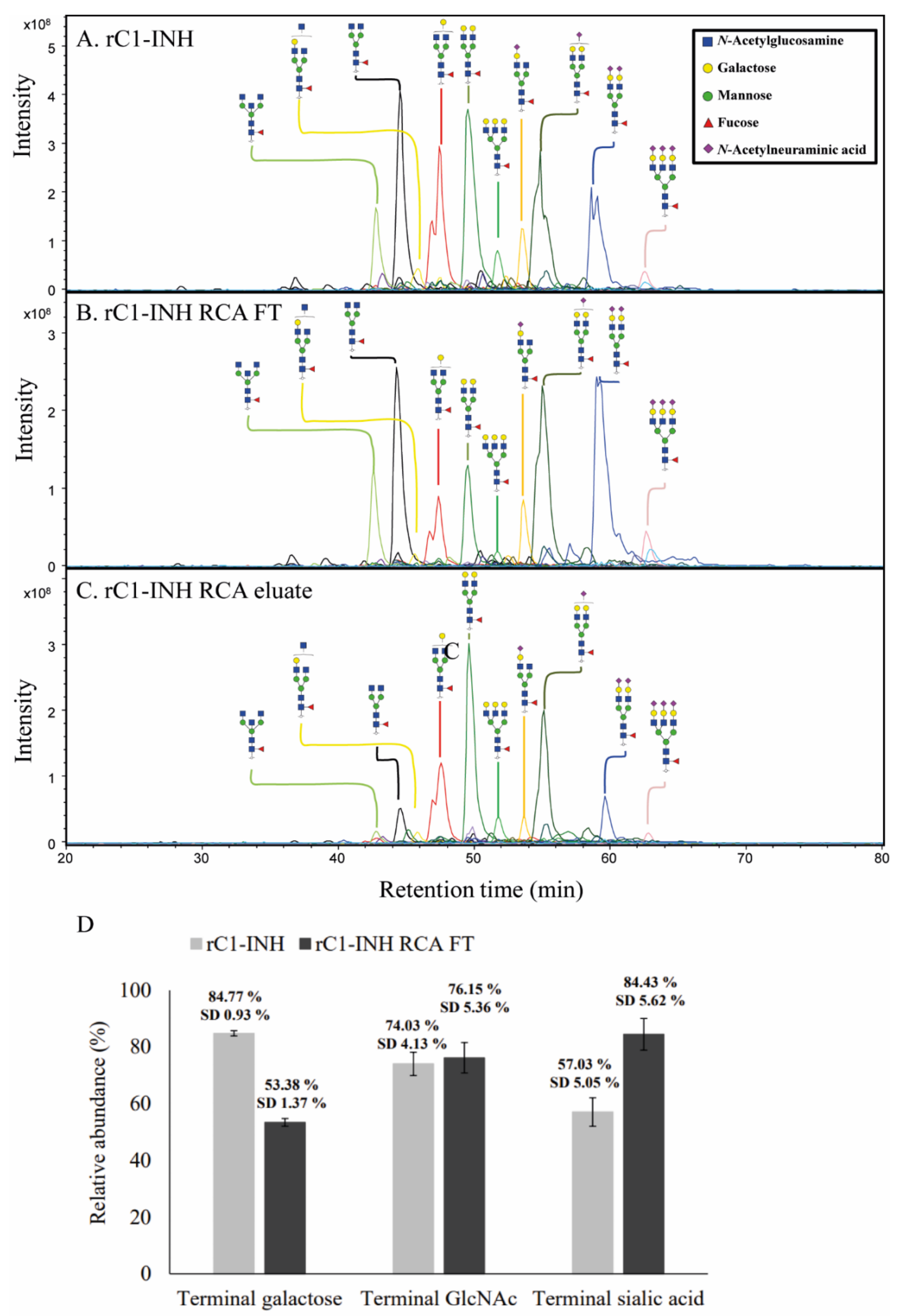
2.2. Terminal Galactose Screening for rC1-INH
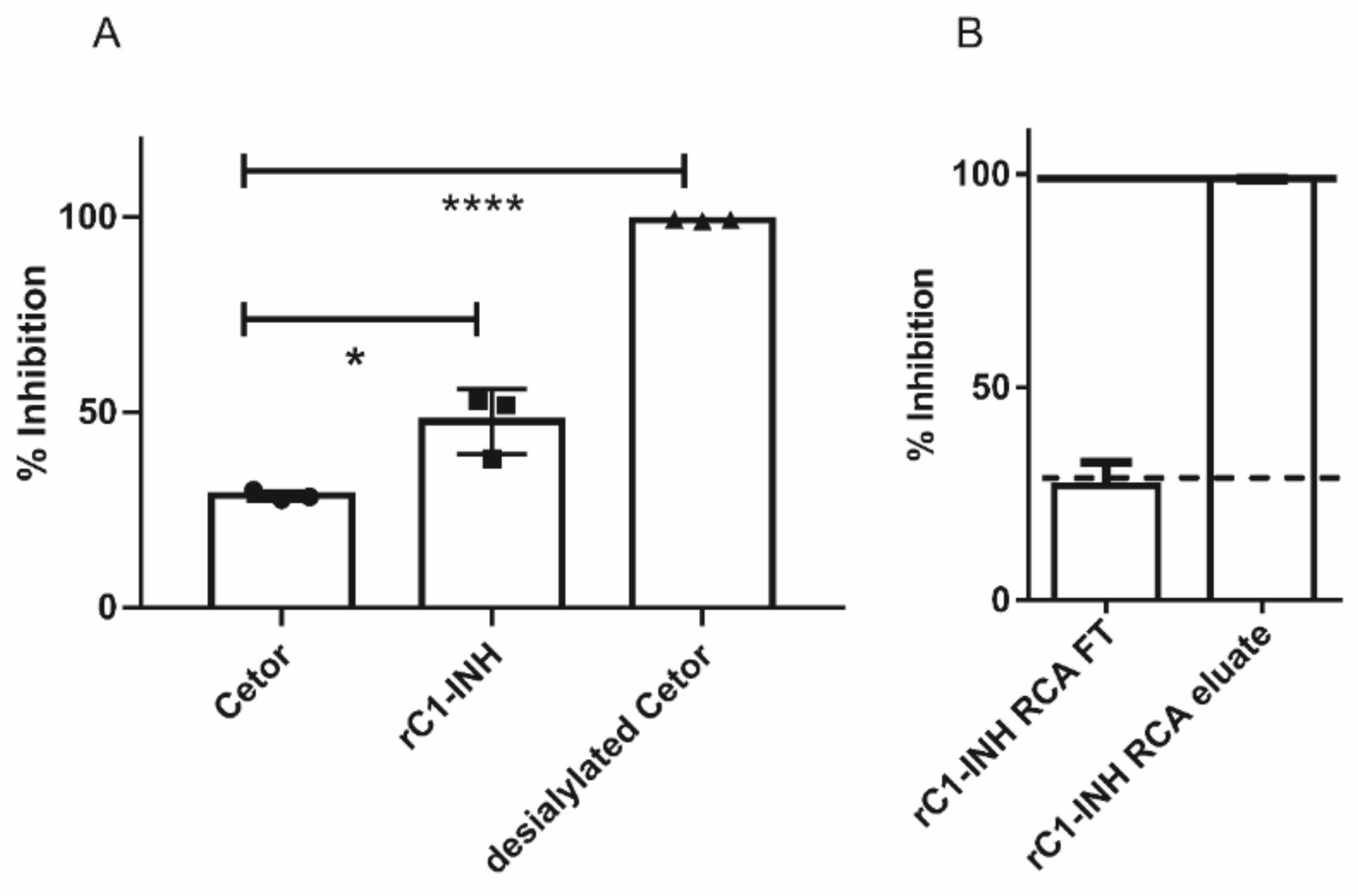
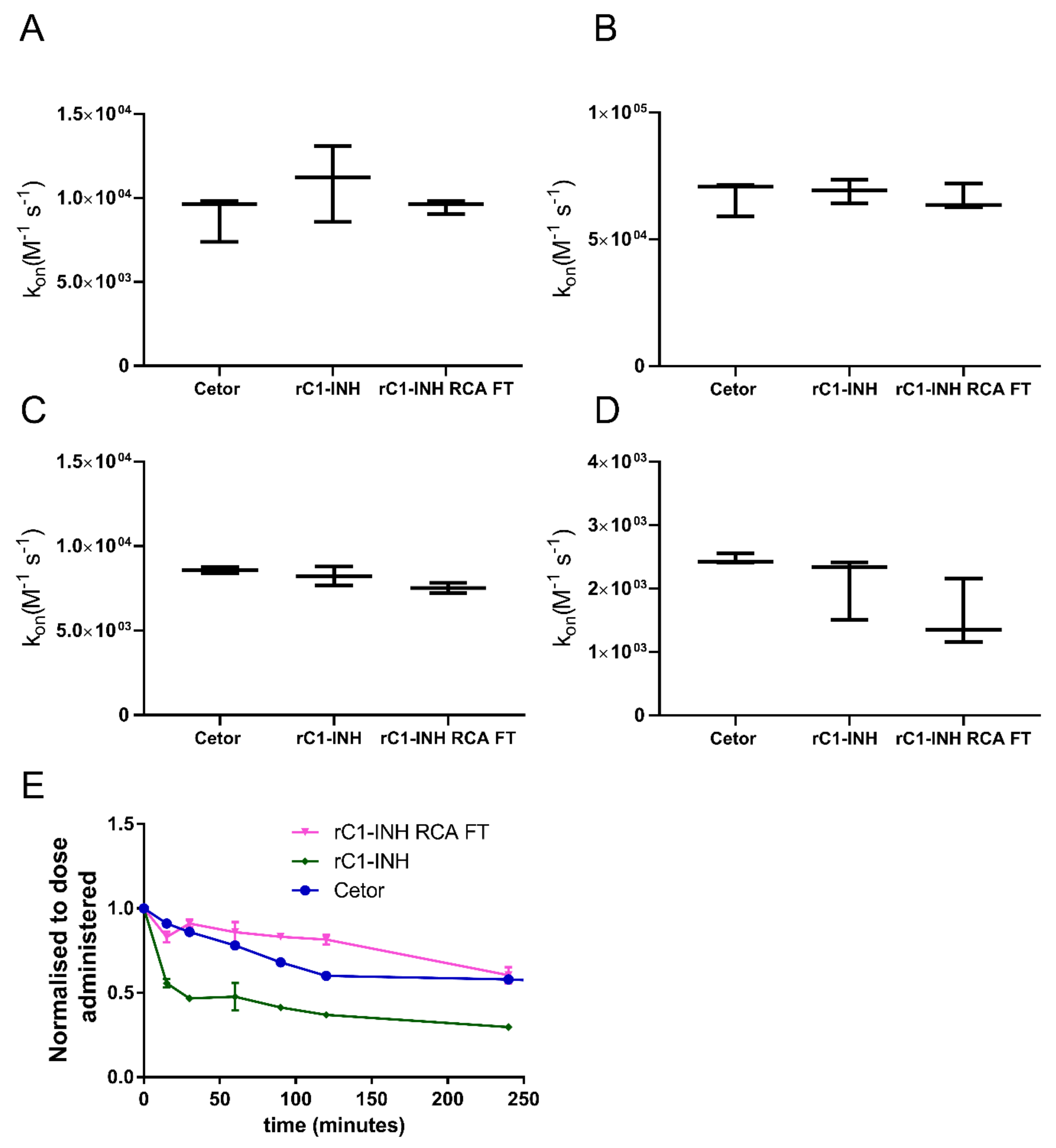
2.3. Removal of Terminal Galactose-Rich Fraction on RCA120 Column Does Not Impair C1-INH Function and Improves In Vivo Half-Life of rC1-INH
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Proteins, Antibodies and Substrates
4.3. Production and Purification of rC1-INH in CHO Cells
4.4. Antigen and Active C1-INH ELISA
4.5. SDS-PAGE
4.6. Kinetic Analyses of Protease Inhibition
4.7. Animal Experiments
4.8. Half-Life of rC1-INH in Rabbits
4.9. RCA120 Inhibition ELISA
4.10. Separation of Terminal Galactose-Rich Fraction on RCA120 Column
4.11. Preparation of N-Glycan Alditols Released from rC1-INH and RCA120 Depleted rC1-INH
4.12. Analysis of N-Glycan Alditols Released from rC1-INH and RCA120-Depleted rC1-INH Using PGC Nano-LC-ESI-MS/MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part i—Molecular mechanisms of activation and regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricklin, D.; Mastellos, D.C.; Reis, E.S.; Lambris, J.D. The renaissance of complement therapeutics. Nat. Rev. Nephrol. 2018, 14, 26–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricklin, D.; Lambris, J.D. Complement-targeted therapeutics. Nat. Biotechnol. 2007, 25, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Zeerleder, S. C1-inhibitor: More than a serine protease inhibitor. Semin. Thromb. Hemost. 2011, 37, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Nicola, S.; Rolla, G.; Brussino, L. Breakthroughs in hereditary angioedema management: A systematic review of approved drugs and those under research. Drugs Context 2019, 8, 212605. [Google Scholar]
- Stavenhagen, K.; Kayili, H.M.; Holst, S.; Koeleman, C.A.M.; Engel, R.; Wouters, D.; Zeerleder, S.; Salih, B.; Wuhrer, M. N- and O-glycosylation Analysis of Human C1-inhibitor Reveals Extensive Mucin-type O-Glycosylation. Mol. Cell. Proteom. 2018, 17, 1225–1238. [Google Scholar] [CrossRef] [Green Version]
- Lamark, T.; Ingebrigtsen, M.; Bjørnstad, C.; Melkko, T.; Mollnes, T.E.; Nielsen, E.W. Expression of Active Human C1 Inhibitor Serpin Domain in Escherichia coli. Protein Expr. Purif. 2001, 22, 349–358. [Google Scholar] [CrossRef]
- Bos, I.G.; de Bruin, E.C.; Karuntu, Y.A.; Modderman, P.W.; Eldering, E.; Hack, C.E. Recombinant human c1-inhibitor produced in pichia pastoris has the same inhibitory capacity as plasma c1-inhibitor. Biochim. Biophys. Acta 2003, 1648, 75–83. [Google Scholar] [CrossRef]
- Van Doorn, M.B.; Burggraaf, J.; Van Dam, T.; Eerenberg, A.; Levi, M.; Hack, C.E.; Schoemaker, R.C.; Cohen, A.F.; Nuijens, J. A phase I study of recombinant human C1 inhibitor in asymptomatic patients with hereditary angioedema. J. Allergy Clin. Immunol. 2005, 116, 876–883. [Google Scholar] [CrossRef]
- Hack, C.E.; Relan, A.; Van Amersfoort, E.S.; Cicardi, M. Target levels of functional C1-inhibitor in hereditary angioedema. Allergy 2012, 67, 123–130. [Google Scholar] [CrossRef]
- Riedl, M.A.; Grivcheva-Panovska, V.; Moldovan, D.; Baker, J.; Yang, W.H.; Giannetti, B.M.; Reshef, A.; Andrejevic, S.; Lockey, R.F.; Hakl, R.; et al. Recombinant human C1 esterase inhibitor for prophylaxis of hereditary angio-oedema: A phase 2, multicentre, randomised, double-blind, placebo-controlled crossover trial. Lancet 2017, 390, 1595–1602. [Google Scholar] [CrossRef]
- Zeerleder, S.; Caliezi, C.; Van Mierlo, G.; Eerenberg-Belmer, A.; Sulzer, I.; Hack, C.E.; Wuillemin, W.A. Administration of C1 Inhibitor Reduces Neutrophil Activation in Patients with Sepsis. Clin. Diagn. Lab. Immunol. 2003, 10, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Wouters, D.; Stephan, F.; Strengers, P.; De Haas, M.; Brouwer, C.; Hagenbeek, A.; Van Oers, M.H.; Zeerleder, S. C1-esterase inhibitor concentrate rescues erythrocytes from complement-mediated destruction in autoimmune hemolytic anemia. Blood 2013, 121, 1242–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, S.C.; Choi, J.; Aubert, O.; Haas, M.; Loupy, A.; Huang, E.; Peng, A.; Kim, I.; Louie, S.; Ammerman, N.; et al. A phase I/II, double-blind, placebo-controlled study assessing safety and efficacy of C1 esterase inhibitor for prevention of delayed graft function in deceased donor kidney transplant recipients. Am. J. Transplant. 2018, 18, 2955–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.Q.; Slingsby, F.C.; Richards, A.; Barlow, P.N. Production of biologically active complement factor H in therapeutically useful quantities. Protein Expr. Purif. 2011, 76, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhouri, F.; De Jorge, E.G.; Brune, F.; Azam, P.; Cook, H.T.; Pickering, M.C. Treatment with human complement factor H rapidly reverses renal complement deposition in factor H-deficient mice. Kidney Int. 2010, 78, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Walsh, G. Biopharmaceutical benchmarks 2014. Nat. Biotechnol. 2014, 32, 992–1000. [Google Scholar] [CrossRef]
- Kunschak, M.; Engl, W.; Maritsch, F.; Rosen, F.S.; Eder, G.; Zerlauth, G.; Schwarz, H.P. A randomized, controlled trial to study the efficacy and safety of C1 inhibitor concentrate in treating hereditary angioedema. Transfusion 1998, 38, 540–549. [Google Scholar] [CrossRef]
- Abdulkarim, A.; Craig, T.J. Hereditary Angioedema; Statpearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Levy, D.S.; Farkas, H.; Riedl, M.A.; Hsu, F.I.; Brooks, J.P.; Cicardi, M.; Feuersenger, H.; Pragst, I.; Reshef, A. Long-term efficacy and safety of subcutaneous C1-inhibitor in women with hereditary angioedema: Subgroup analysis from an open-label extension of a phase 3 trial. Allergy Asthma Clin. Immunol. 2020, 16, 8–9. [Google Scholar] [CrossRef]
- Mannucci, P.M. Hemophilia therapy: The future has begun. Haematologica 2020, 105, 545–553. [Google Scholar] [CrossRef]
- Cunningham-Rundles, C. Key aspects for successful immunoglobulin therapy of primary immunodeficiencies. Clin. Exp. Immunol. 2011, 164, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, T.; Khan, M.A.; Ijaz, B.; Ahmed, N.; Rahman, Z.U.; Latif, M.S.; Ali, Q.; Rana, M.A. Glycosylation of Recombinant Anticancer Therapeutics in Different Expression Systems with Emerging Technologies. Cancer Res. 2018, 78, 2787–2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higel, F.; Sandl, T.; Kao, C.-Y.; Pechinger, N.; Sörgel, F.; Friess, W.; Wolschin, F.; Seidl, A. N-glycans of complex glycosylated biopharmaceuticals and their impact on protein clearance. Eur. J. Pharm. Biopharm. 2019, 139, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Michelfelder, S.; Parsons, J.; Bohlender, L.L.; Hoernstein, S.N.W.; Niederkrüger, H.; Busch, A.; Krieghoff, N.; Koch, J.; Fode, B.; Schaaf, A.; et al. Moss-Produced, Glycosylation-Optimized Human Factor H for Therapeutic Application in Complement Disorders. J. Am. Soc. Nephrol. 2017, 28, 1462–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quastel, M.; Harrison, R.; Cicardi, M.; Alper, C.A.; Rosen, F.S. Behavior in vivo of normal and dysfunctional C1 inhibitor in normal subjects and patients with hereditary angioneurotic edema. J. Clin. Investig. 1983, 71, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Koles, K.; Van Berkel, P.H.; Pieper, F.R.; Nuijens, J.H.; Mannesse, M.L.; Vliegenthart, J.F.; Kamerling, J.P. N- and O-glycans of recombinant human C1 inhibitor expressed in the milk of transgenic rabbits. Glycobiology 2004, 14, 51–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaderi, D.; Zhang, M.; Hurtado-Ziola, N.; Varki, A. Production platforms for biotherapeutic glycoproteins. Occurrence, impact, and challenges of non-human sialylation. Biotechnol. Genet. Eng. Rev. 2012, 28, 147–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sola, R.J.; Griebenow, K. Glycosylation of therapeutic proteins: An effective strategy to optimize efficacy. BioDrugs 2010, 24, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Solá, R.J.; Griebenow, K. Effects of glycosylation on the stability of protein pharmaceuticals. J. Pharm. Sci. 2009, 98, 1223–1245. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Luo, S.; Zhang, B. The use of lectin microarray for assessing glycosylation of therapeutic proteins. mAbs 2016, 8, 524–535. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, S.; Halim, A.; Schulz, M.A.; Frodin, M.; Rahman, S.H.; Vester-Christensen, M.B.; Behrens, C.; Kristensen, C.; Vakhrushev, S.Y.; et al. Engineered CHO cells for production of diverse, homogeneous glycoproteins. Nat. Biotechnol. 2015, 33, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Amann, T.; Hansen, A.H.; Kol, S.; Hansen, H.G.; Arnsdorf, J.; Nallapareddy, S.; Voldborg, B.; Lee, G.M.; Andersen, M.R.; Kildegaard, H.F. Glyco-engineered CHO cell lines producing alpha-1-antitrypsin and C1 esterase inhibitor with fully humanized N-glycosylation profiles. Metab. Eng. 2019, 52, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büttner-Mainik, A.; Parsons, J.; Jérôme, H.; Hartmann, A.; Lamer, S.; Schaaf, A.; Schlosser, A.; Zipfel, P.F.; Reski, R.; Decker, E.L. Production of biologically active recombinant human factor H in Physcomitrella. Plant Biotechnol. J. 2011, 9, 373–383. [Google Scholar] [CrossRef]
- Graf, L. Extended Half-Life Factor VIII and Factor IX Preparations. Transfus. Med. Hemother. 2018, 45, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuijens, J.H.; Huijbregts, C.C.; Eerenberg-Belmer, A.J.; Abbink, J.J.; Strack van Schijndel, R.J.; Felt-Bersma, R.J.; Thijs, L.G.; Hack, C.E. Quantification of plasma factor xiia-cl(-)-inhibitor and kallikrein-cl(-)-inhibitor complexes in sepsis. Blood 1988, 72, 1841–1848. [Google Scholar] [CrossRef] [Green Version]
- Nuijens, J.H.; Huijbregts, C.C.; Van Mierlo, G.M.; Hack, C.E. Inactivation of C-1 inhibitor by proteases: Demonstration by a monoclonal antibody of a neodeterminant on inactivated, non-complexed C-1 inhibitor. Immunology 1987, 61, 387–389. [Google Scholar]
- Bos, I.G.; van Mierlo, G.J.; Bleeker, W.K.; Rigter, G.M.; te Velthuis, H.; Dickneite, G.; Hack, C.E. The potentiation of human c1-inhibitor by dextran sulphate is transient in vivo: Studies in a rat model. Int. Immunopharmacol. 2001, 1, 1583–1595. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, G.; Han, Z.; Yang, B.; Hu, Y.; Zhao, X.; Wu, J.; Lv, Y.; Chai, W. Specificities of Ricinus communis agglutinin 120 interaction with sulfated galactose. FEBS Lett. 2011, 585, 3927–3934. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Madunic, K.; Holst, S.; Zhang, J.; Jin, C.; Ten Dijke, P.; Karlsson, N.G.; Stavenhagen, K.; Wuhrer, M. Development of a 96-well plate sample preparation method for integrated N- and O-glycomics using porous graphitized carbon liquid chromatography-mass spectrometry. Mol. Omics 2020, 16, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Madunic, K.; Zhang, T.; Mayboroda, O.A.; Holst, S.; Stavenhagen, K.; Jin, C.; Karlsson, N.G.; Lageveen-Kammeijer, G.S.M.; Wuhrer, M. Colorectal cancer cell lines show striking diversity of their O-glycome reflecting the cellular differentiation phenotype. Cell. Mol. Life Sci. 2020, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, N.G.; Wilson, N.L.; Wirth, H.J.; Dawes, P.; Joshi, H.; Packer, N.H. Negative ion graphitised carbon nano-liquid chromatography/mass spectrometry increases sensitivity for glycoprotein oligosaccharide analysis. Rapid Commun. Mass Spectrom. 2004, 18, 2282–2292. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, N.G.; Schulz, B.L.; Packer, N.H. Structural determination of neutral O-linked oligosaccharide alditols by negative ion LC-electrospray-MSn. J. Am. Soc. Mass Spectrom. 2004, 15, 659–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anugraham, M.; Everest-Dass, A.V.; Jacob, F.; Packer, N.H. A platform for the structural characterization of glycans enzymatically released from glycosphingolipids extracted from tissue and cells. Rapid Commun. Mass Spectrom. 2015, 29, 545–561. [Google Scholar] [CrossRef]
- Ceroni, A.; Maass, K.; Geyer, H.; Geyer, R.; Dell, A.; Haslam, S.M. GlycoWorkbench: A Tool for the Computer-Assisted Annotation of Mass Spectra of Glycans. J. Proteome Res. 2008, 7, 1650–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, C.A.; Gasteiger, E.; Packer, N.H. Glycomod—A software tool for determining glycosylation compositions from mass spectrometric data. Proteomics 2001, 1, 340–349. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeerleder, S.; Engel, R.; Zhang, T.; Roem, D.; van Mierlo, G.; Wagenaar-Bos, I.; van Ham, S.M.; Wuhrer, M.; Wouters, D.; Jongerius, I. Sugar Matters: Improving In Vivo Clearance Rate of Highly Glycosylated Recombinant Plasma Proteins for Therapeutic Use. Pharmaceuticals 2021, 14, 54. https://doi.org/10.3390/ph14010054
Zeerleder S, Engel R, Zhang T, Roem D, van Mierlo G, Wagenaar-Bos I, van Ham SM, Wuhrer M, Wouters D, Jongerius I. Sugar Matters: Improving In Vivo Clearance Rate of Highly Glycosylated Recombinant Plasma Proteins for Therapeutic Use. Pharmaceuticals. 2021; 14(1):54. https://doi.org/10.3390/ph14010054
Chicago/Turabian StyleZeerleder, Sacha, Ruchira Engel, Tao Zhang, Dorina Roem, Gerard van Mierlo, Ineke Wagenaar-Bos, Sija Marieke van Ham, Manfred Wuhrer, Diana Wouters, and Ilse Jongerius. 2021. "Sugar Matters: Improving In Vivo Clearance Rate of Highly Glycosylated Recombinant Plasma Proteins for Therapeutic Use" Pharmaceuticals 14, no. 1: 54. https://doi.org/10.3390/ph14010054
APA StyleZeerleder, S., Engel, R., Zhang, T., Roem, D., van Mierlo, G., Wagenaar-Bos, I., van Ham, S. M., Wuhrer, M., Wouters, D., & Jongerius, I. (2021). Sugar Matters: Improving In Vivo Clearance Rate of Highly Glycosylated Recombinant Plasma Proteins for Therapeutic Use. Pharmaceuticals, 14(1), 54. https://doi.org/10.3390/ph14010054