
Cucumis sativus L. Seeds Ameliorate Muscular Spasm-Induced Gastrointestinal and Respiratory Disorders by Simultaneously Inhibiting Calcium Mediated Signaling Pathway

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
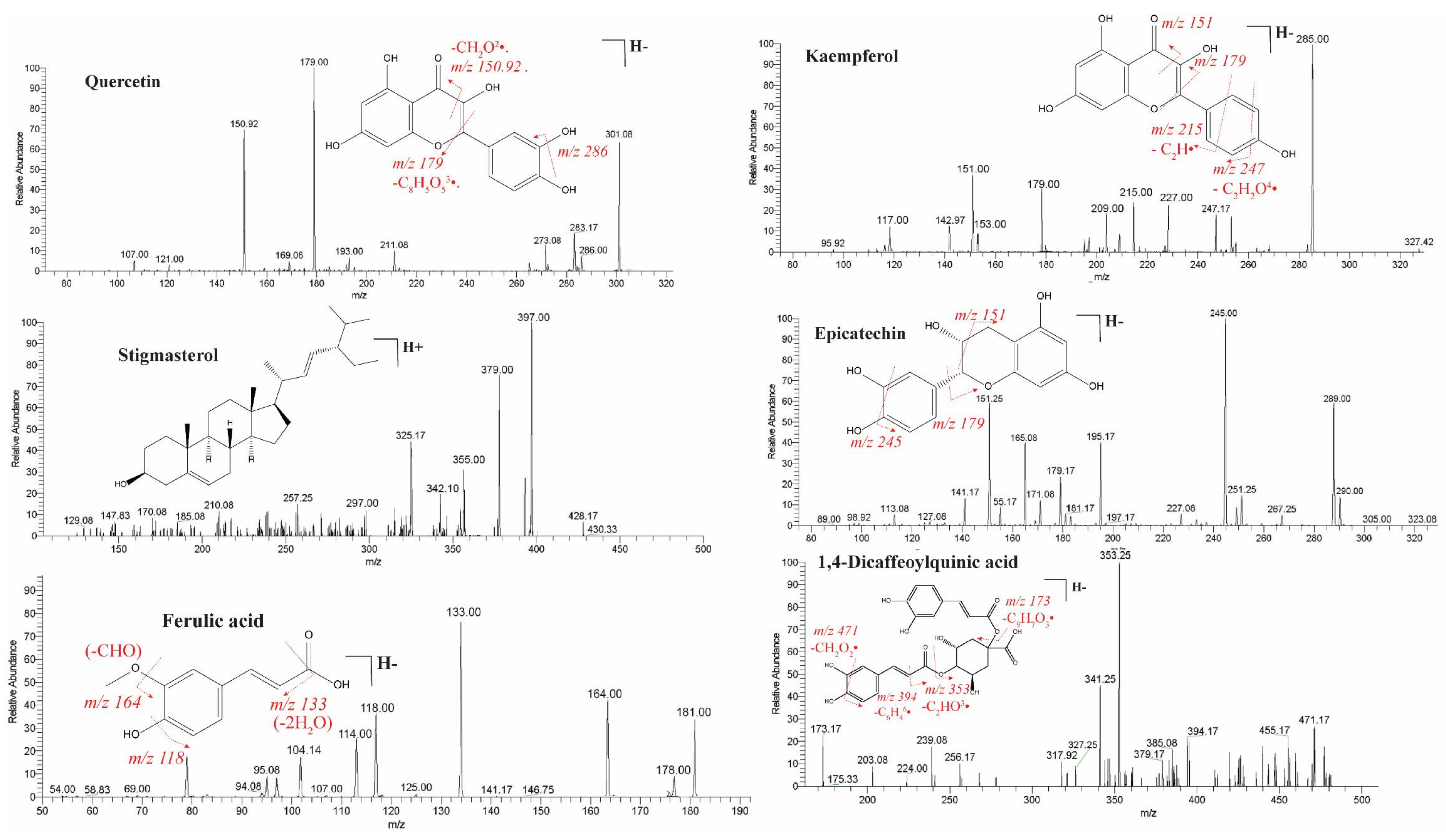
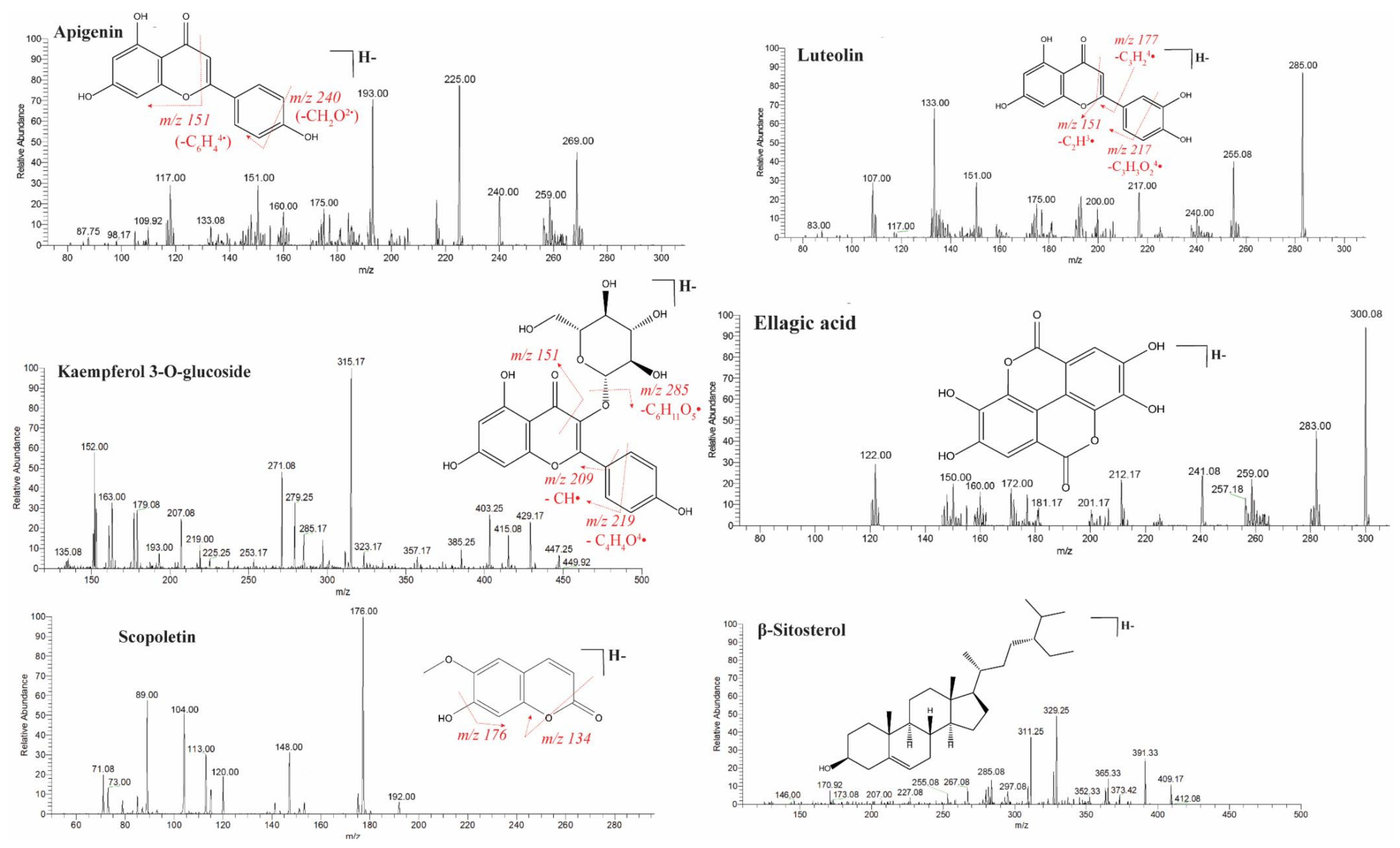
2.1. Identification of Bioactive Compounds by LC-ESI-MS/MS Analysis
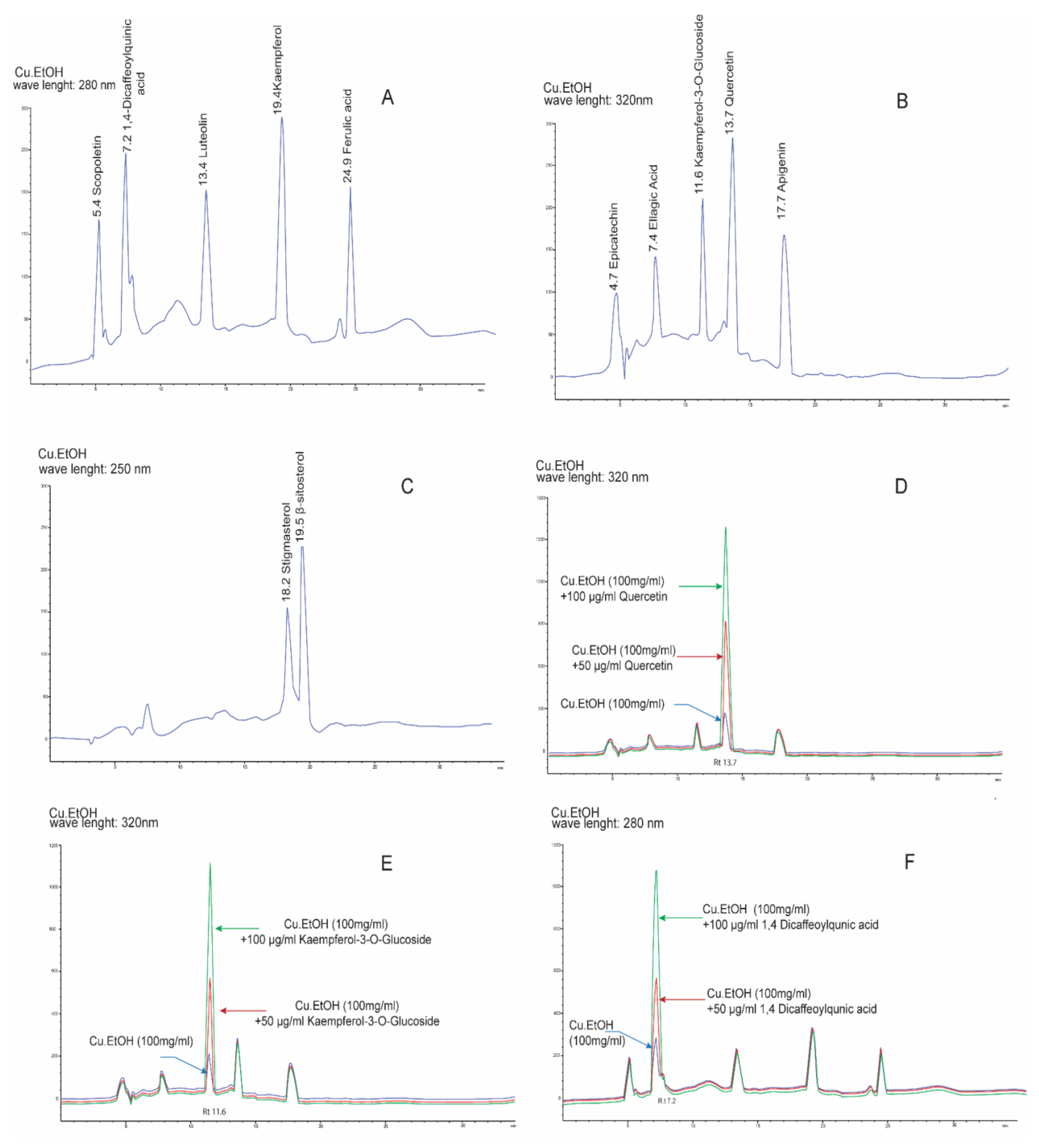
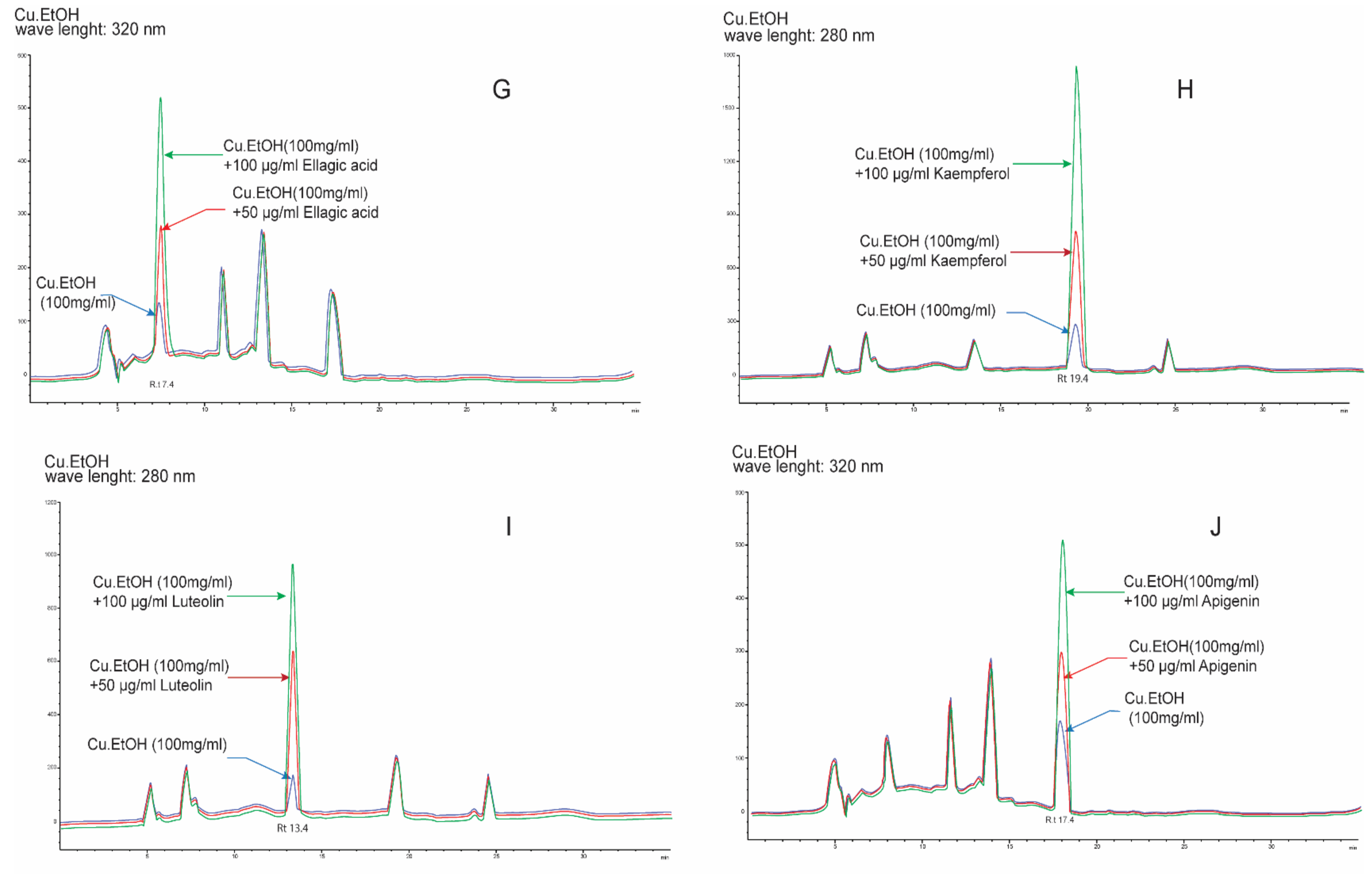
2.2. Optimization of HPLC Conditions and Method Validation
Quantification Analysis of Bioactive Compounds by HPLC
2.3. In Silico Studies
2.3.1. ADMET and Drug-Likeness
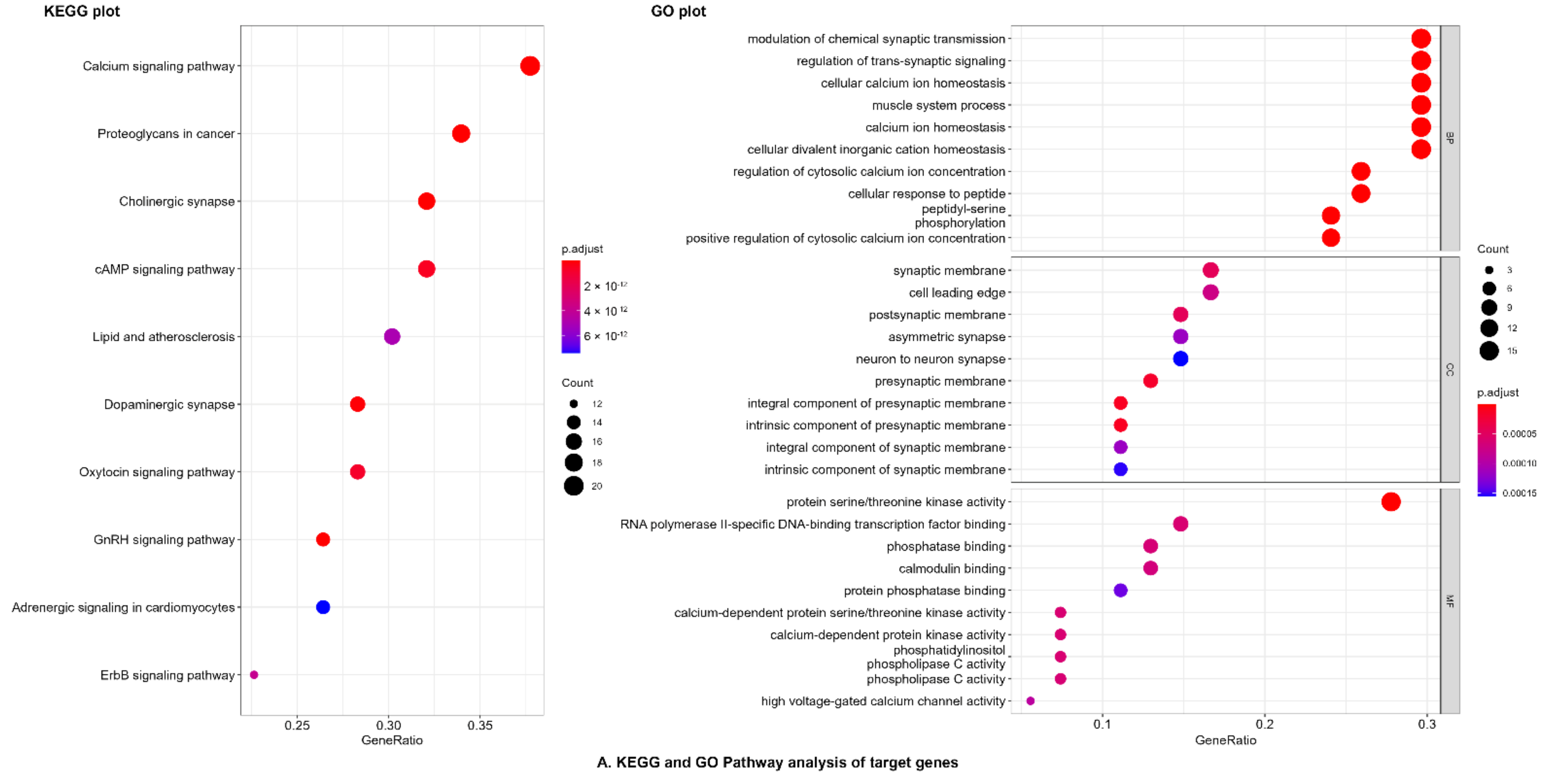
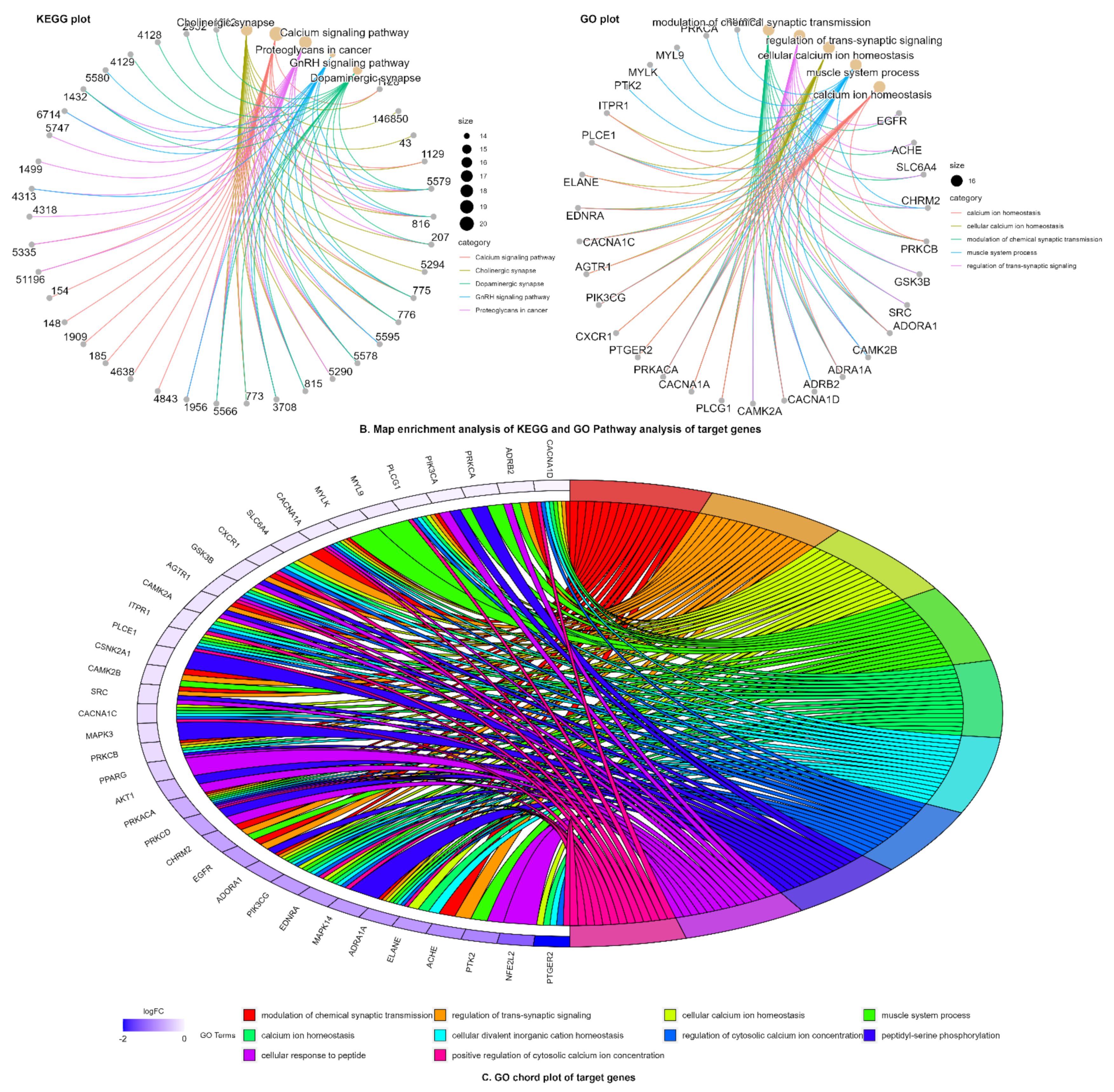
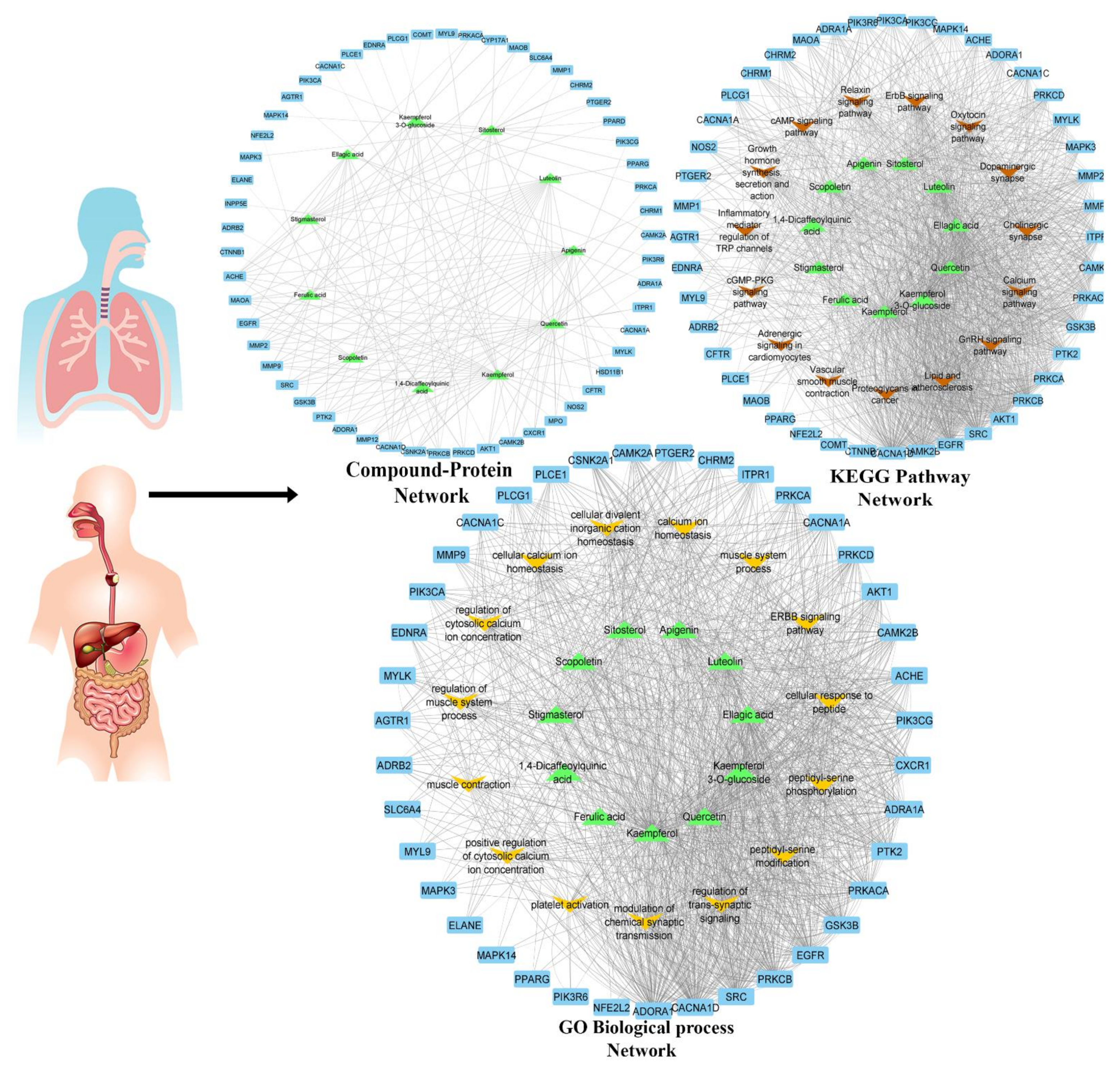
2.3.2. Network Pharmacology Analysis
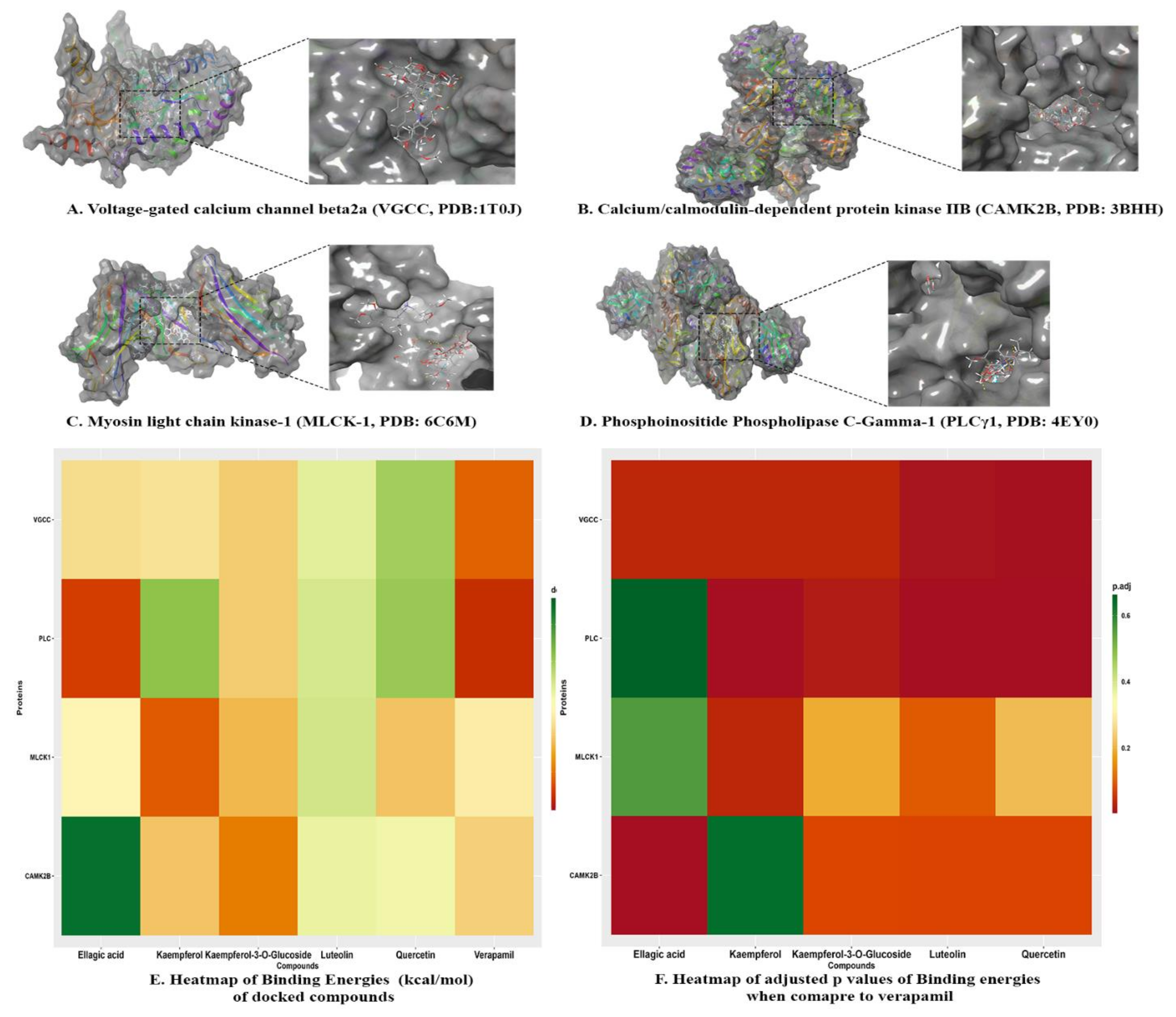
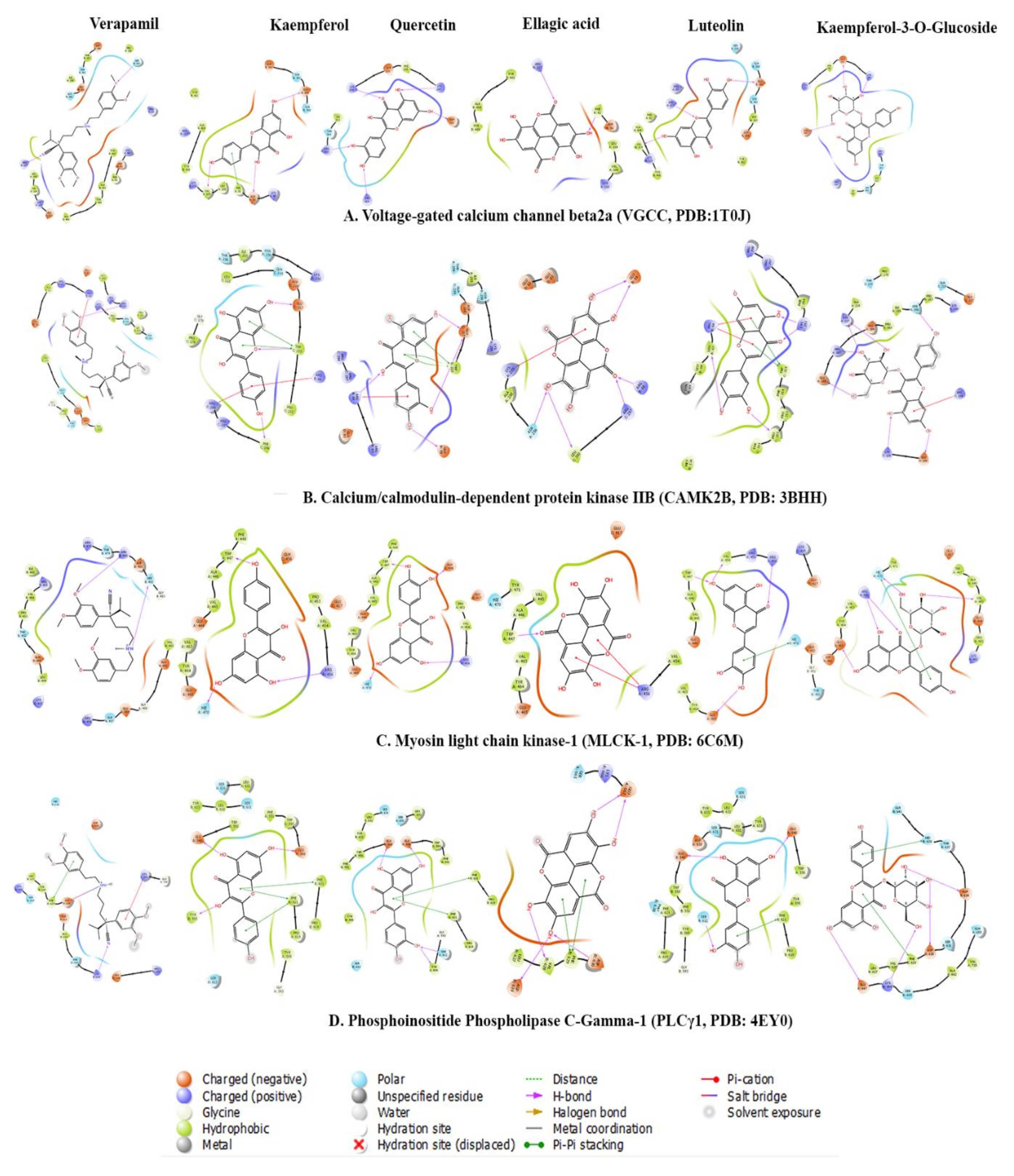
2.3.3. Molecular Docking
2.4. Isolated Tissue Experimentation
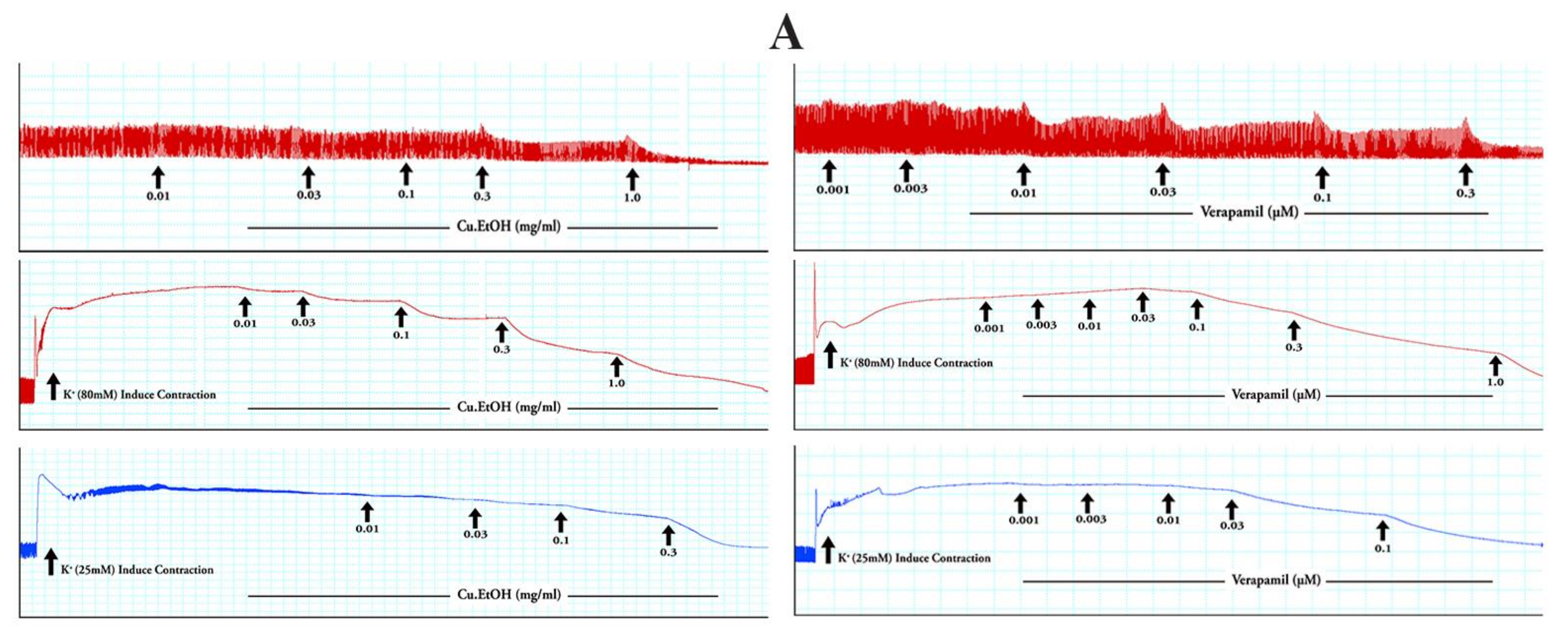
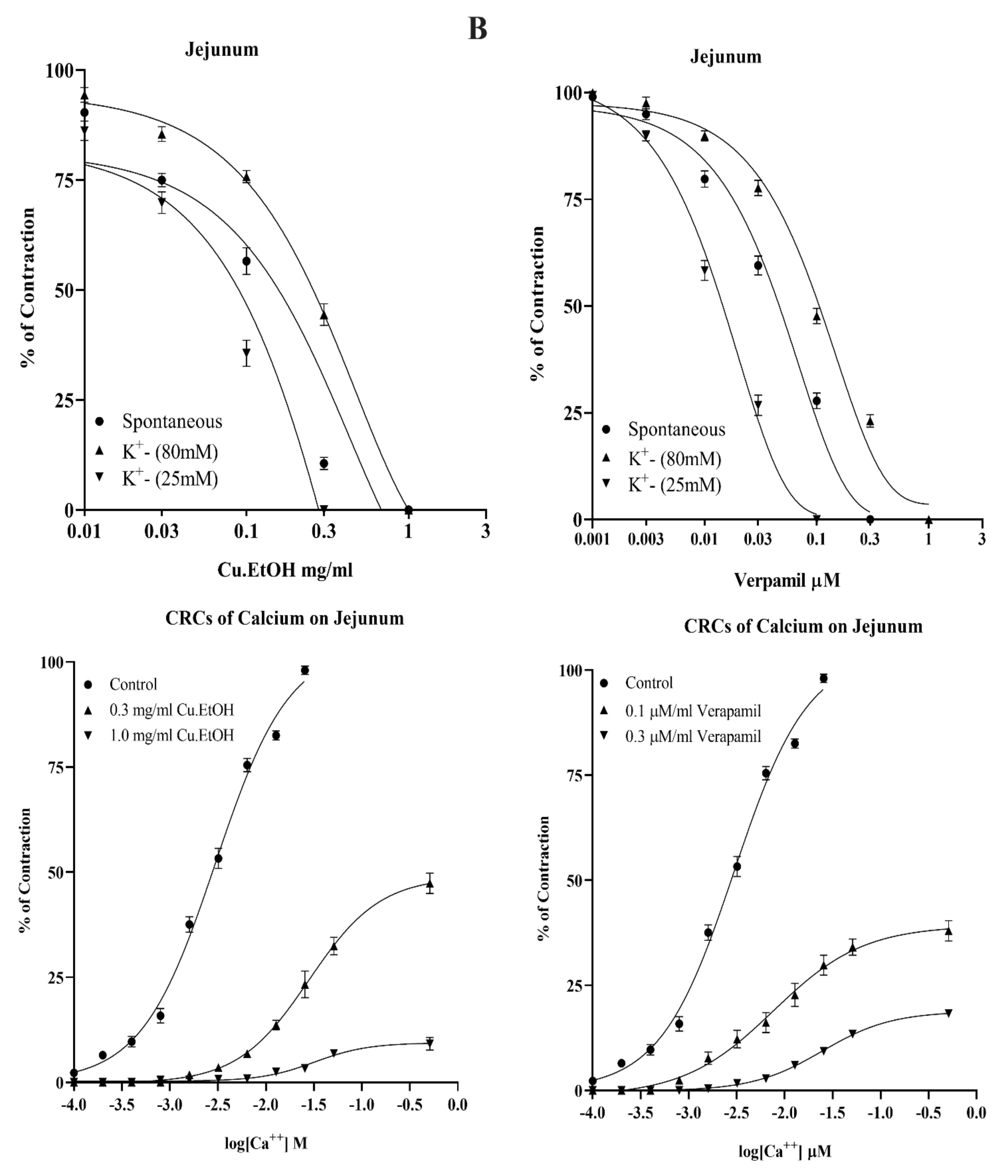
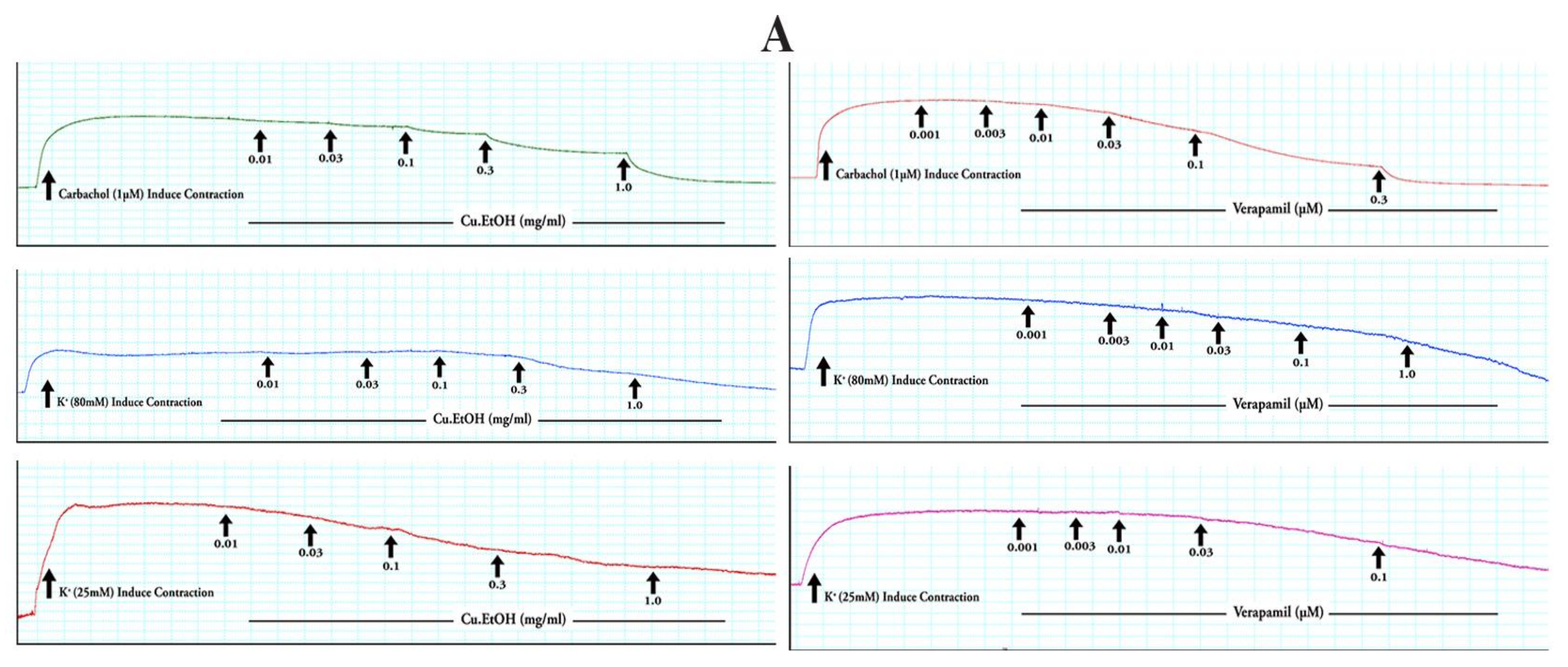
2.4.1. Effects on Isolated Rabbit Jejunum Preparation
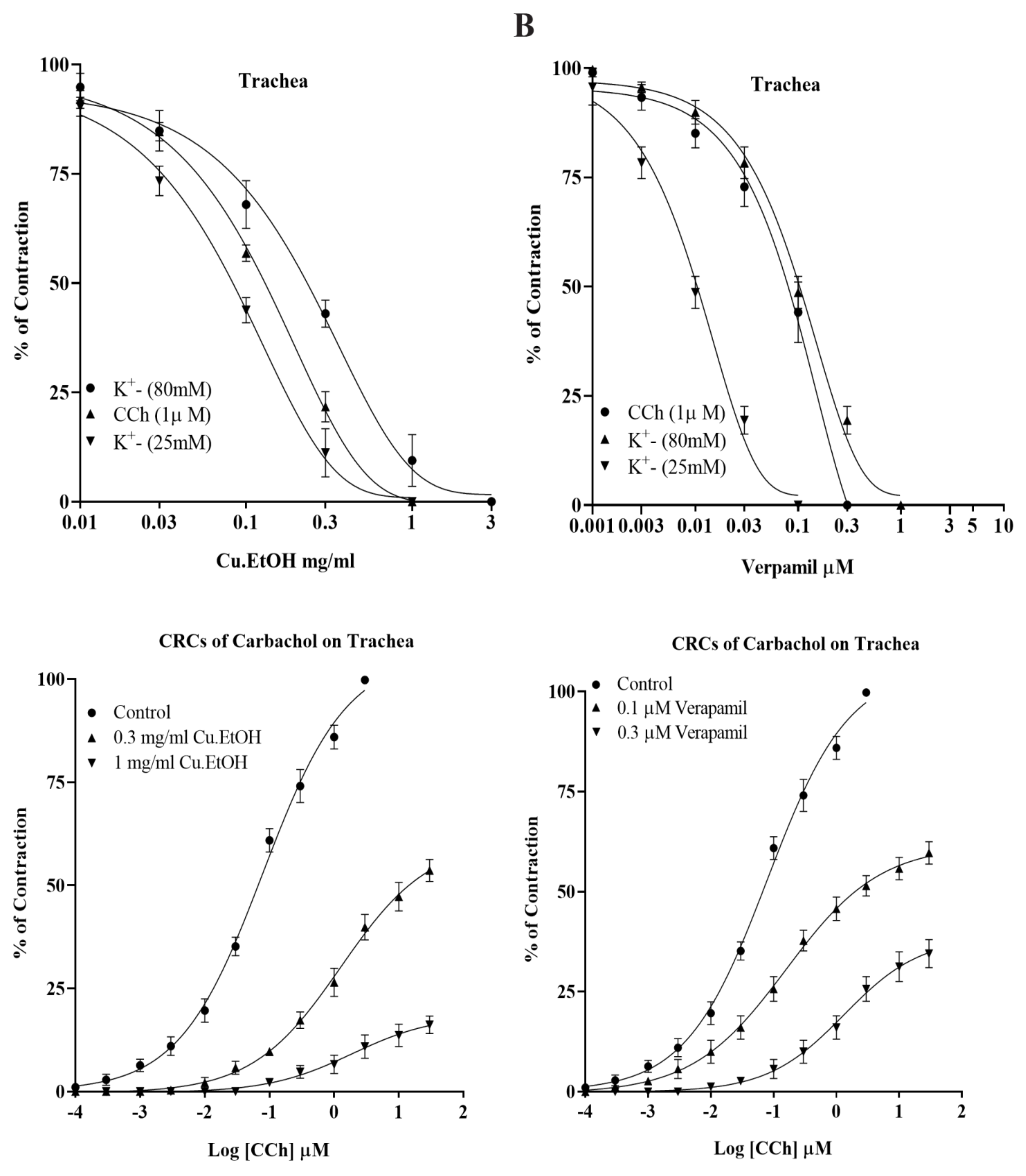
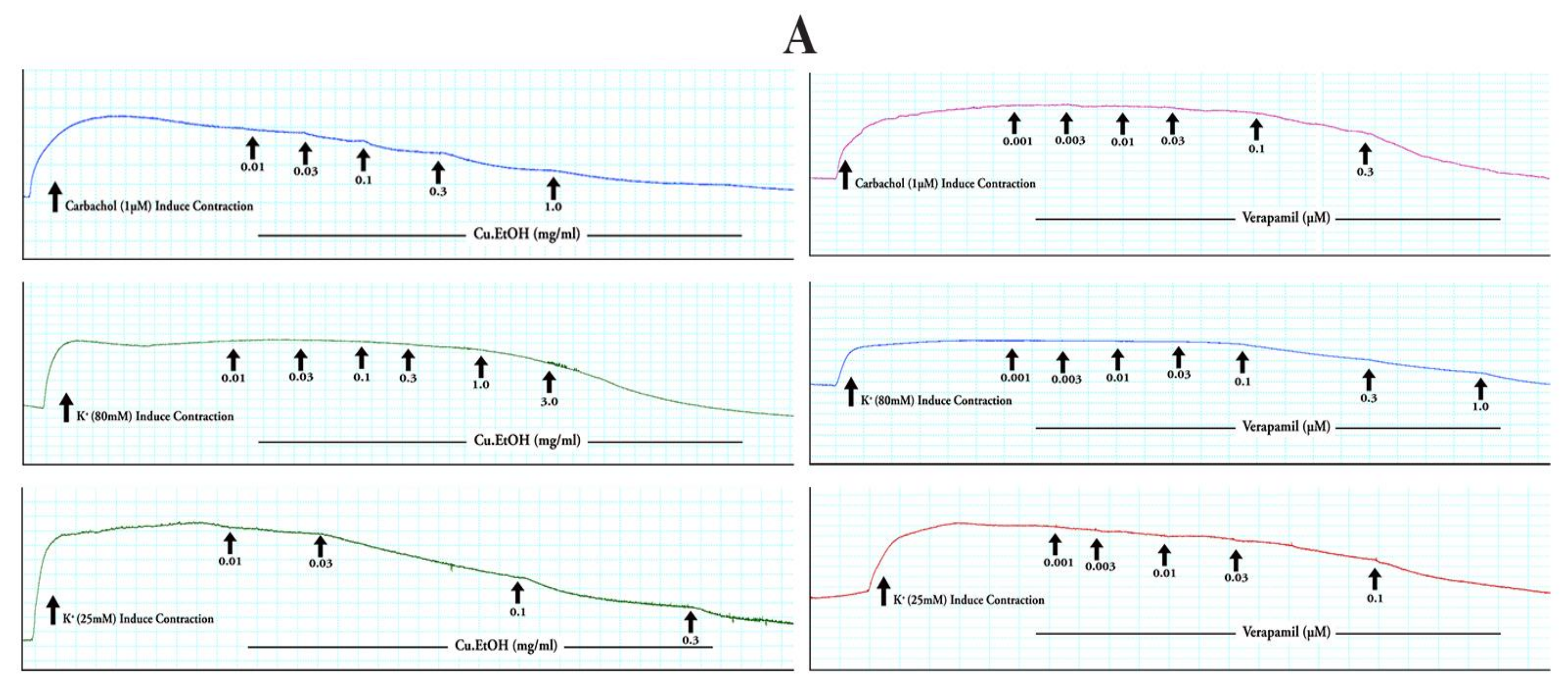
2.4.2. Effect on Isolated Rabbit Tracheal Preparations
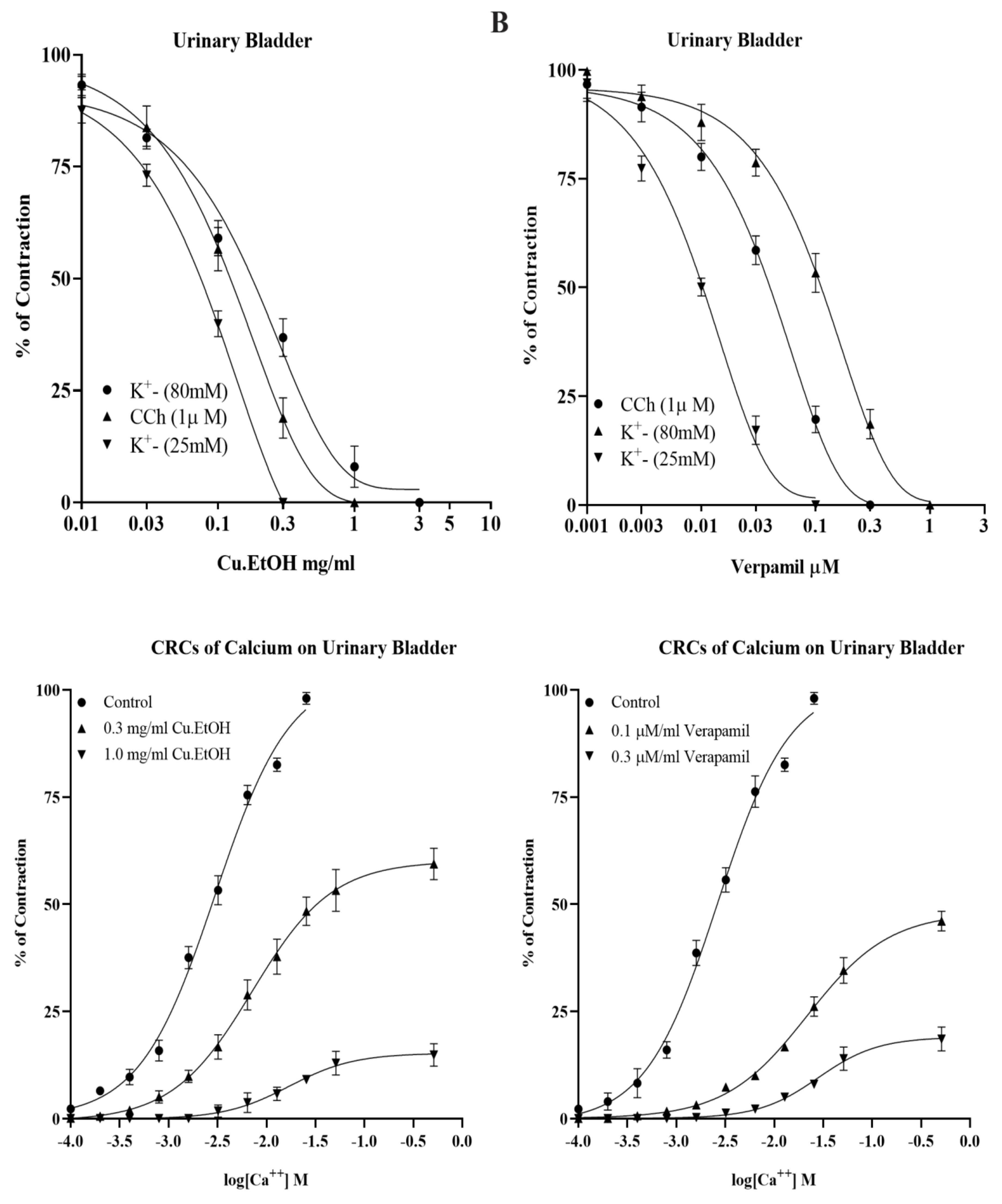
2.4.3. Effect on Isolated Rabbit Urinary Bladder Preparations
2.5. In Vivo Experiments
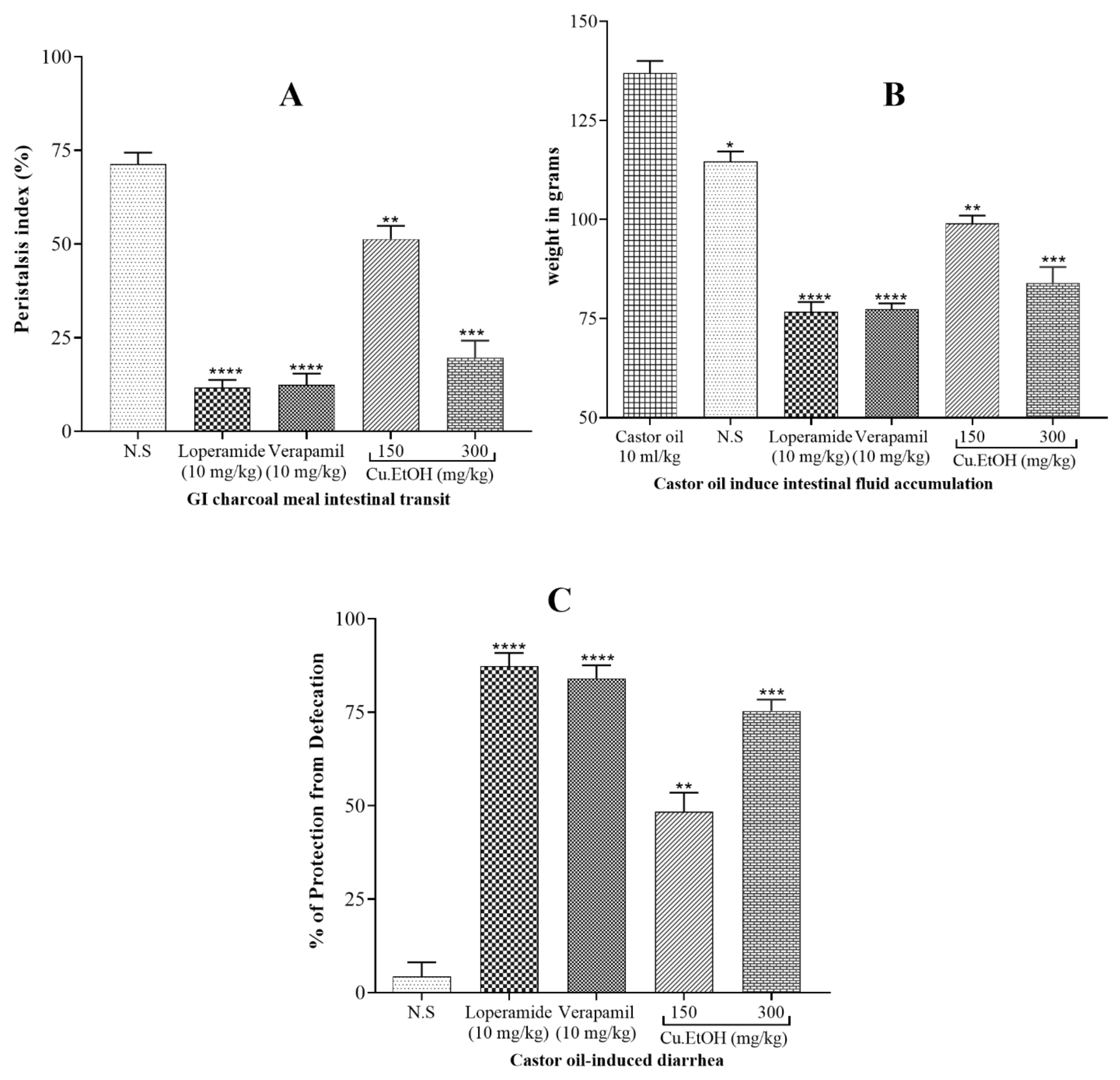
2.5.1. Effect on GI Charcoal Meal Intestinal Transit
2.5.2. Effect on Castor Oil-Induced Diarrhea
2.5.3. Effect on Intestinal Fluid Accumulation
3. Discussion
4. Materials and Methods
4.1. Preparation of Extract
4.2. Chemicals
4.3. Sample Preparation for HPLC and LC-ESI-MS/MS
4.4. LC ESI-MS/MS Analysis
4.5. Quantification of Bioactive Compounds by Using Analytical HPLC-DAD UV/Vis
4.5.1. HPLC Method Optimization
4.5.2. Validation of the Analytical Method
4.6. In Silico Approaches
4.6.1. ADMET and Drug-likeness
4.6.2. Network Pharmacology Analysis
4.6.3. Molecular Docking
4.7. Animals and Housing Conditions
4.8. Isolated Tissue Experimentation
4.8.1. Isolated Rabbit Jejunum Preparations
4.8.2. Isolated Rabbit Tracheal Preparations
4.8.3. Isolated Urinary Bladder Preparations
4.9. In Vivo Experimentation
4.9.1. Evaluation of Maximum Tolerated Dose
4.9.2. Charcoal Meal GI Transit Test
4.9.3. Castor Oil-Induced Diarrhea
4.9.4. Intestinal Fluid Accumulation
4.10. Statistical Analysis
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BP | Biological process |
| C-T-D | Compound and targeted disease genes network |
| C-T-P | Compound, targeted disease genes and pathway network |
| CAMK2B | Calcium/calmodulin-dependent protein kinase IIB |
| CCB | Calcium Channel blocker |
| CCh | Carbachol |
| CRC | Concentration-response curves |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| Log Ki | Logarithmic of Inhibition Constant (Ki) |
| MM-GBSA | molecular mechanics energies combined generalized Born and surface area |
| MLCK-1 | Myosin light chain kinase-1 |
| Cu.EtOH | C. sativus seeds hydroethanolic extract |
| VGCC | Voltage-gated calcium channel β2a |
References
- Mariod, A.A.; Mirghani, M.E.S.; Hussein, I. Cucumis sativus Cucumber. Unconv. Oilseeds Oil Sources 2017, 89–94. [Google Scholar] [CrossRef]
- Renner, S.S.; Pandey, A.K. The Cucurbitaceae of India: Accepted names, synonyms, geographic distribution, and information on images and DNA sequences. PhytoKeys 2013, 20, 53–118. [Google Scholar] [CrossRef] [PubMed]
- Hameed, S.; Hans, S.; Nandan, S.; Fatima, Z. Mechanistic insights into the antimycobacterial action of unani formulation, Qurs Sartan Kafoori. J. Tradit. Complement. Med. 2021. [Google Scholar] [CrossRef]
- Mosleh, G.; Badr, P.; Zaeri, M.; Mohagheghzadeh, A. Potentials of Antitussive Traditional Persian Functional Foods for COVID-19 Therapy†. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Afzal, M.; Khan, N.; Ghufran, A.; Iqbal, A.; Inamuddin, M. Diuretic and nephroprotective effect of Jawarish Zarooni Sada—A polyherbal unani formulation. J. Ethnopharmacol. 2004, 91, 219–223. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Medicinal Plants; Khare, C.P., Ed.; Springer: New York, NY, USA, 2007; ISBN 978-0-387-70637-5. [Google Scholar]
- Sharma, S.; Yadav, S.; Singh, G.; Paliwal, S.; Dwivedi, J. First report on laxative activity of Cucumis Sativus. Int. J. Pharm. Sci. Rev. Res. 2012, 12, 129–131. [Google Scholar]
- Saha, T.; Aoun, J.; Sarkar, P.; Bourdelais, A.J.; Baden, D.G.; Leblanc, N.; Hamlyn, J.M.; Woodward, O.M.; Hoque, K.M. Cucumis sativus extract elicits chloride secretion by stimulation of the intestinal TMEM16A ion channel. Pharm. Biol. 2021, 59, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.M.; Khan, M.; Ahmad, M.; Zafar, M.; Jahan, S.; Sultana, S. Ethnopharmacological application of medicinal plants to cure skin diseases and in folk cosmetics among the tribal communities of North-West Frontier Province, Pakistan. J. Ethnopharmacol. 2010, 128, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Dwivedi, J.; Paliwal, S. Evaluation of antacid and carminative properties of Cucumis sativus under simulated conditions. Sch. Res. Libr. Der Pharm. Lett. 2012, 4, 234–239. [Google Scholar]
- Rajasree, R.S.; Sibi, P.I.; Francis, F.; William, H. Phytochemicals of cucurbitaceae family—A review. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 113–123. [Google Scholar]
- Tuama, A.A.; Mohammed, A.A. Phytochemical screening and in vitro antibacterial and anticancer activities of the aqueous extract of Cucumis sativus. Saudi J. Biol. Sci. 2018, 26, 600–604. [Google Scholar] [CrossRef]
- Pertami, S.B.; Rahayu, D.Y.S.; Budiono, B. EFFECT OF CUCUMBER (CUCUMIS SATIVUS) JUICE ON LOWERING BLOOD PRESSURE IN ELDERLY. Public Health Indones. 2017, 3, 30–36. [Google Scholar] [CrossRef]
- Agatemor, U.; Nwodo, O.; Anosike, C. Anti-inflammatory Activity of Cucumis sativus L. Br. J. Pharm. Res. 2015, 8, 1–8. [Google Scholar] [CrossRef]
- Soltani, R.; Hashemi, M.; Farazmand, A.; Asghari, G.; Heshmat-Ghahdarijani, K.; Kharazmkia, A.; Ghanadian, S.M. Evaluation of the Effects ofCucumis sativusSeed Extract on Serum Lipids in Adult Hyperlipidemic Patients: A Randomized Double-Blind Placebo-Controlled Clinical Trial. J. Food Sci. 2016, 82, 214–218. [Google Scholar] [CrossRef]
- Minaiyan, M.; Zolfaghari, B.; Kamal, A. Effect of hydroalcoholic and buthanolic extract of Cucumis sativus seeds on blood glucose level of normal and streptozotocin-induced diabetic rats. Iran. J. Basic Med. Sci. 2011, 14, 436–442. [Google Scholar] [CrossRef]
- Vetriselvan, S.; Subasini, U.; Velmurugan, C.; Muthuramu, T.; Revathy, J. Anti-inflammatory activity of Cucumis sativus seed in carrageenan and xylene induced edema model using albino wistar rats. Int. J. Biopharm. 2013, 4, 34–37. [Google Scholar]
- Wahid, S.; Alqahtani, A.; Alam Khan, R. Analgesic and anti-inflammatory effects and safety profile of Cucurbita maxima and Cucumis sativus seeds. Saudi J. Biol. Sci. 2021, 28, 4334–4341. [Google Scholar] [CrossRef]
- Gill, N.S.; Garg, M.; Bansal, R.; Sood, S.; Muthuraman, A.; Bali, M.; Sharma, P.D. Evaluation of Antioxidant and Antiulcer Potential of Cucumis sativum L. Seed Extract in Rats. Asian J. Clin. Nutr. 2009, 1, 131–138. [Google Scholar] [CrossRef]
- Khan, M.S.; Tariq, S.S.H. Medicinal treatment of multiple renal calculi (Hisat-E-Kulyah) and bilateral ureteric calculi (Hisat-E-Halib) by Unani Pharmacopoeial formulations—A Case Study. World J. Adv. Res. Rev. 2021, 11, 183–189. [Google Scholar] [CrossRef]
- Anwar, S.; Khan, N.A.; Amin, K.M.Y.; Ahmad, G. Effects of Banadiq-al Buzoor in some renal disorders. Hamdard Med. 1999, 42, 31–36. [Google Scholar]
- Alam, S.; Khan, N.A.; Nasiruddin, M. The Effect of Qurs-e-Zarishk Sagheer (A Compound Unani Formulation) on Liver Enzymes in CCl4 Induced Hepatotoxicity in Rats. Hippocrat. J. Unani Med. 2014, 9, 1–12. [Google Scholar]
- Siddika, M.; Hasnat, R.; Bahar, E. Thrombolytic (in vitro) and analgesic (in vivo) effect of methanolic extract of Cucumis sativus. Pharma Innov. 2016, 3, 01–07. [Google Scholar]
- Rehman, S.; Imran, M. An ethno-botanical review of seeds of Cucumis sativa (Maghz-e-tukhme khiyarain) from Unani Medicine and its Pharmacological Updates. Int. J. Adv. Pharm. Med. Bioallied Sci. 2021, 9, 31–36. [Google Scholar]
- Babu, P.A.S.; Aafrin, B.V.; Archana, G.; Sabina, K.; Sudharsan, K.; Krishnan, K.R.; Babuskin, S.; Sivarajan, M.; Sukumar, M. Polyphenolic and phytochemical content of Cucumis sativus seeds and study on mechanism of preservation of nutritional and quality outcomes in enriched mayonnaise. Int. J. Food Sci. Technol. 2016, 51, 1417–1424. [Google Scholar] [CrossRef]
- Gilani, A.H.; Khan, A.-U.; Raoof, M.; Ghayur, M.N.; Siddiqui, B.; Vohra, W.; Begum, S. Gastrointestinal, selective airways and urinary bladder relaxant effects of Hyoscyamus niger are mediated through dual blockade of muscarinic receptors and Ca2+ channels. Fundam. Clin. Pharmacol. 2008, 22, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-Q.; Wilkinson, B. Drug discovery beyond the ‘rule-of-five’. Curr. Opin. Biotechnol. 2007, 18, 478–488. [Google Scholar] [CrossRef]
- Gajjar, N.D.; Dhameliya, T.M.; Shah, G.B. In search of RdRp and Mpro inhibitors against SARS CoV-2: Molecular docking, molecular dynamic simulations and ADMET analysis. J. Mol. Struct. 2021, 1239, 130488. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.-T.; Liu, S.-Y.; Kuang, Y.-J.; Jiang, Z.-M.; Lin, Y.; Xie, Z.-S.; Liu, E.-H. Network pharmacology analysis and experimental validation to explore the mechanism of sea buckthorn flavonoids on hyperlipidemia. J. Ethnopharmacol. 2020, 264, 113380. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, M.-H.; Wang, F.; Chen, P.-Y.; Ke, X.-G.; Yu, B.; Yang, Y.-F.; You, P.-T.; Wu, H.-Z. Network pharmacology and molecular docking reveal the mechanism of Scopoletin against non-small cell lung cancer. Life Sci. 2021, 270, 119105. [Google Scholar] [CrossRef]
- Sirous, H.; Chemi, G.; Campiani, G.; Brogi, S. An integrated in silico screening strategy for identifying promising disruptors of p53-MDM2 interaction. Comput. Biol. Chem. 2019, 83. [Google Scholar] [CrossRef]
- Kuhn, B.; Kollman, P.A. Binding of a Diverse Set of Ligands to Avidin and Streptavidin: An Accurate Quantitative Prediction of Their Relative Affinities by a Combination of Molecular Mechanics and Continuum Solvent Models. J. Med. Chem. 2000, 43, 3786–3791. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-B.; Li, Q.-Y.; Chen, Q.-L.; Su, S.-B. Network Pharmacology: A New Approach for Chinese Herbal Medicine Research. Evid. -Based Complement. Altern. Med. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Park, S.-Y.; Lee, H.-J.; Kim, C.-E. A Systems-Level Analysis of Mechanisms of Platycodon grandiflorum Based on A Network Pharmacological Approach. Molecules 2018, 23, 2841. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Sharkey, K.A. Pharmacotherapy of Gastric Acidity, Peptic Ulcers, and Gastroesophageal Reflux Disease. In Goodman & Gilman’s The Pharmacological Basis of Therapeutics; Brunton, L.L., Hilal-Dandan, R., Knollmann, B.C., Eds.; McGraw-Hill: New York, NY, USA, 2018; pp. 1309–1322. ISBN 978-0-07-176939-6. [Google Scholar]
- Saqib, F.; Janbaz, K.H. Rationalizing ethnopharmacological uses of Alternanthera sessilis: A folk medicinal plant of Pakistan to manage diarrhea, asthma and hypertension. J. Ethnopharmacol. 2016, 182, 110–121. [Google Scholar] [CrossRef]
- Bin-Jumah, M.; Alwakeel, S.S.; Moga, M.; Buvnariu, L.; Bigiu, N.; Zia-Ul-Haq, M. Application of Carotenoids in Cosmetics. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Sharkey, K.A.; MacNaughton, W.K. Gastrointestinal Motility and Water Flux, Emesis, and Biliary and Pancreatic Disease. In Goodman & Gilman’s The Pharmacological Basis of Therapeutics; Brunton, L.L., Hilal-Dandan, R., Knollmann, B.C., Eds.; McGraw-Hill: New York, NY, USA, 2018; pp. 921–944. ISBN 978-1-25-958474-9. [Google Scholar]
- Saqib, F.; Janbaz, K.H. Ethnopharmacological basis for folkloric claims of Anagallis arvensis Linn. (Scarlet Pimpernel) as prokinetic, spasmolytic and hypotensive in province of Punjab, Pakistan. J. Ethnopharmacol. 2020, 267, 113634. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M. Past, present and future of Carotenoids Research. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 827–854. [Google Scholar]
- Gohar, U.F.; Iqbal, I.; Shah, Z.; Mukhtar, H.; Zia-Ul-Haq, M. COVID-19: Recent Developments in Therapeutic Approaches. In Alternative Medicine Interventions for COVID-19; Zia-Ul-Haq, M., Bin-Jumah, M.N., Alothamn, S.I., Henidi, H.A., Eds.; Springer: Cham, Switzerland, 2021; pp. 249–274. [Google Scholar]
- Galati, E.M.; Lanuzza, F.; Occhiuto, F.; Monforte, M.T.; Tripodo, M.M.; D’Angelo, V. Antioxidant phytochemicals of Opuntia ficus-indica (L.) Mill. cladodes with potential anti-spasmodic activity. Pharmacogn. Mag. 2017, 13, 424–S429. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Riaz, M.; Ahmedah, H.T. Carotenoids and Periodontal Diseases. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 715–720. [Google Scholar]
- Zia-Ul-Haq, M. Historical and introductory aspects of carotenoids. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 1–42. [Google Scholar]
- Amira, S.; Rotondo, A.; Mulè, F. Relaxant effects of flavonoids on the mouse isolated stomach: Structure-activity relationships. Eur. J. Pharmacol. 2008, 599, 126–130. [Google Scholar] [CrossRef]
- Janbaz, K.H.; Jan, A.; Qadir, M.I.; Gilani, A.H. Spasmolytic, bronchodilator and vasorelaxant activity of methanolic extract of Tephrosia purpurea. Acta Pol. Pharm.—Drug Res. 2013, 70, 261–269. [Google Scholar]
- Ko, W.-C.; Liu, P.-Y.; Chen, J.-L.; Leu, I.-J.; Shih, C.-M. Relaxant Effects of Flavonoids in Isolated Guinea Pig Trachea and Their Structure-Activity Relationships. Planta Med. 2003, 69, 1086–1090. [Google Scholar] [CrossRef] [Green Version]
- Capasso, R.; Aviello, G.; Romano, B.; Atorino, G.; Pagano, E.; Borrelli, F. Inhibitory effect of quercetin on rat trachea contractility in vitro. J. Pharm. Pharmacol. 2009, 61, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Djelili, H.; Arrar, L.; Naline, E.; Devillier, P. Relaxant Effects of Quercetin and Rutin on Human Isolated Bronchus. Chin. Med. 2012, 03, 94–100. [Google Scholar] [CrossRef]
- Chan, H.J.; Ji, Y.L.; Chul, H.C.; Chang, J.K. Anti-asthmatic action of quercetin and rutin in conscious guinea-pigs challenged with aerosolized ovalbumin. Arch. Pharm. Res. 2007, 30, 1599–1607. [Google Scholar] [CrossRef]
- Agbor, G.A.; Longo, F.; Makong, E.A.; Tarkang, P.A. Evaluation of the antidiarrheal and antioxidant properties of Justicia hypocrateriformis. Pharm. Biol. 2014, 52, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Crowe, A.; Wong, P. Potential roles of P-gp and calcium channels in loperamide and diphenoxylate transport. Toxicol. Appl. Pharmacol. 2003, 193, 127–137. [Google Scholar] [CrossRef]
- Murek, M.; Kopic, S.; Geibel, J. Evidence for intestinal chloride secretion. Exp. Physiol. 2010, 95, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Seraglio, S.K.T.; Valese, A.C.; Daguer, H.; Bergamo, G.; Azevedo, M.S.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Development and validation of a LC-ESI-MS/MS method for the determination of phenolic compounds in honeydew honeys with the diluted-and-shoot approach. Food Res. Int. 2016, 87, 60–67. [Google Scholar] [CrossRef]
- Syed, M.; Khan, M.N.; Khadim, A.; Shadab, H.; Perveen, A.; El-Seedi, H.R.; Musharraf, S.G. Chemical fingerprinting of three Anemone species and an adulteration study to detect cross mixing of medicinal plants by HPLC-HR-ESI-MS/MS method. J. King Saud Univ.—Sci. 2021, 33, 101461. [Google Scholar] [CrossRef]
- Qamar, M.; Akhtar, S.; Ismail, T.; Yuan, Y.; Ahmad, N.; Tawab, A.; Ismail, A.; Barnard, R.T.; Cooper, M.A.; Blaskovich, M.A.T.; et al. Syzygium cumini (L.), Skeels fruit extracts: In vitro and in vivo anti-inflammatory properties. J. Ethnopharmacol. 2021, 271, 113805. [Google Scholar] [CrossRef]
- Rowan, A.N. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 1979; Volume 8, ISBN 0309186633. [Google Scholar]
- Ghayur, M.N.; Gilani, A. ALPHA—Adrenergic Receptor Mediated Hypertensive and Vasoconstrictor Effects of Dietary Radish Leaves Extract. J. Health Sci. 2007, 53, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Elasoru, S.E.; Rhana, P.; Barreto, T.D.O.; de Souza, D.L.N.; Menezes-Filho, J.E.R.; Souza, D.S.; Moreira, M.V.L.; Campos, M.T.G.; Adedosu, O.T.; Roman-Campos, D.; et al. Andrographolide protects against isoproterenol-induced myocardial infarction in rats through inhibition of L-type Ca2+ and increase of cardiac transient outward K+ currents. Eur. J. Pharmacol. 2021, 906, 174194. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, M.H.; Munir, S.; Khalid, U.A.; Asrar, M.; Gilani, A.H. Antidiarrhoeal, antisecretory and antispasmodic activities of Matricaria chamomilla are mediated predominantly through K(+)-channels activation. BMC Complement. Altern. Med. 2015, 15, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
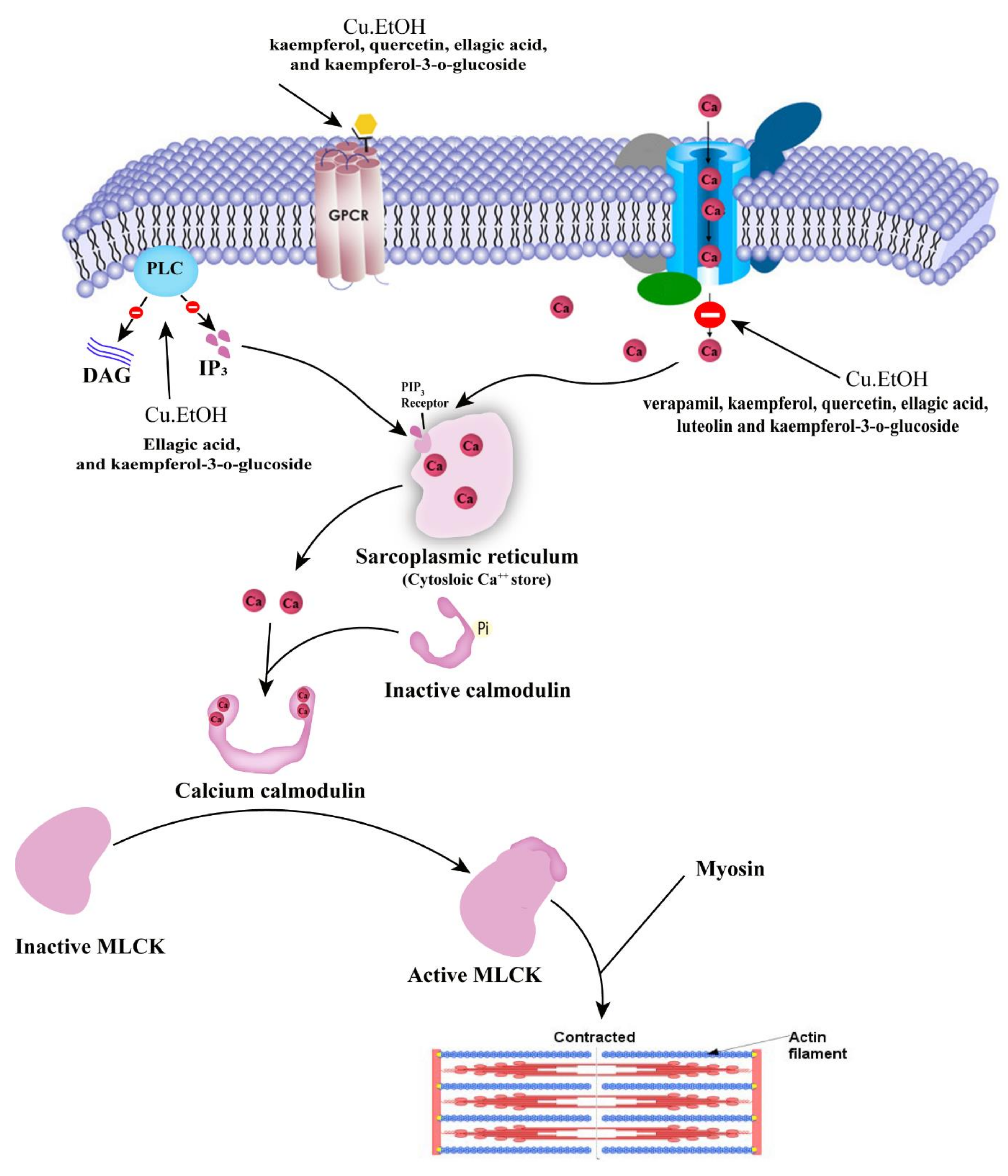{kind=link}
| Sr. No | Rt (min) | Molecular Weight | Observed MS (m/z) | Calculated MS (m/z) | Error (ppm) | Precursor Type | ESI-IT MS/MS (Ions) | Empirical Formula | Proposed Compound | Class |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. | 0.83 | 414.7 | 413.3785 | 413.3781 | −1 | [M−H]− | 412.08, 391.33, 365.33, 352.33, 311.25, 297, 285.08, 255.08, 171 | C29H50O | β-Sitosterol | Phytosterols |
| 2. | 0.89 | 312.5 | 311.3342 | 311.3335 | −2.2 | [M−H]− | 311, 221, 184, 119 | C20H32O2 | Arachidic acid | Fatty acids/ω-6 fatty acid |
| 3. | 1.62 | 162.14 | 161.0252 | 161.0249 | −1.9 | [M−H]− | 161, 133, 117, | C9H6O3 | Umbelliferone | Hydroxycoumarins |
| 4. | 2.8 | 462.36 | 461.0783 | 461.0775 | −1.7 | [M−H]− | 461, 285, 175, 151, 133,107 | C21H18O12 | Luteolin 7-O glucuronide | Flavone glucuronide |
| 5. | 3.4 | 458.37 | 457.0756 | 457.0749 | −1.5 | [M−H]− | 305, 169, 125 | C22H18O11 | Epigallocatechin gallate | Catechin gallates |
| 6. | 4.06 | 594.52 | 593.5036 | 593.5031 | −0.8 | [M−H]− | 593, 285, 151, 133, 107 | C27H30O15 | Luteolin 7-O-rutinoside | Flavonoid glycosides |
| 7. | 4.2 | 624.54 | 623.1618 | 623.1615 | −0.5 | [M−H]− | 623,315, 300, 271, 285, 151 | C28H32O16 | Narcissin/Isorhmnetin 3-O-rutinoside | Flavonoid glycosides |
| 8. | 4.2 | 462.36 | 461.073 | 461.0735 | 1.1 | [M−H]− | 461, 285, 257, 229, 175, 113 | C21H18O12 | Kaempferol-3-O-glucuronoside/Kaempferol 3-glucuronide | Flavonoid glycosides |
| 9. | 4.28 | 448.38 | 447.0943 | 447.0938 | −1.1 | [M−H]− | 447.09, 284, 285, 271, 284, 253, 179, 151 | C21H20O11 | Kaempferol-3-O-glucoside | Flavonoid glycosides |
| 10. | 4.3 | 178.18 | 177.05125 | 177.05134 | 0.5 | [M−H]− | 177, 135, 133, 119 | C10H10O3 | 4-Methoxycinnamic acid | Cinnamic acids |
| 11. | 4.3 | 346.37 | 345.031 | 345.0312 | 0.6 | [M−H]− | 345.52, 300, 226, 221, 206 | C19H22O6 | Gibberellin A3 | Diterpenoids |
| 12. | 4.34 | 478.4 | 477.1038 | 477.1034 | −0.8 | [M−H]− | 477,314, 299, 285, 271, 243,151 | C22H22O12 | Isorhamnetin-3-O-glucoside | Flavonoid glycosides |
| 13. | 4.63 | 192.17 | 191.0357 | 191.0355 | −1 | [M−H]− | 192, 191, 176, 148, 137, 104 | C10H8O4 | Scopoletin | Hydroxycoumarins |
| 14. | 4.7 | 432.38 | 431.0984 | 431.099 | 1.4 | [M−H]− | 431, 285, 284, 255, 227, 183 | C21H20O10 | Kaempferol-3-O-rhamnoside | Flavonoid glycosides |
| 15. | 5.2 | 448.38 | 447.0933 | 447.0934 | 0.2 | [M−H]− | 445.25, 417.08, 357.17, 327.17, 297, 269, 225, 171 | C21H20O11 | Orientin | Flavone glucoside |
| 16. | 6.2 | 256.42 | 255.2325 | 255.2322 | −1.2 | [M−H]− | 255, 235, 217, 181 | C16H32O2 | Palmitic acid | Fatty acids |
| 17. | 6.3 | 270.24 | 269.0425 | 269.0434 | 3.3 | [M−H]− | 241, 225, 197, 157, 133 | C15H10O5 | Genistein | Isoflavones |
| 18. | 6.45 | 290.27 | 289.0717 | 289.0719 | 0.7 | [M−H]− | 290, 289, 245, 179, 165.08, 151.25 | C15H14O6 | Epicatechin | Catechin/Flavonoids |
| 19. | 6.6 | 272.25 | 271.0632 | 271.0639 | 2.6 | [M−H]− | 271, 177, 151, 119, 107 | C15H12O5 | Naringenin | Flavanones |
| 20. | 7.1 | 286.24 | 285.042 | 285.0415 | −1.8 | [M−H]− | 285, 217, 175, 151, 133,107 | C15H10O6 | Luteolin | Flavones |
| 21. | 7.4 | 196.16 | 195.0508 | 195.0503 | −2.6 | [M−H]− | 75,99, 101,129, 159, 177 | C6H12O7 | Gluconic acid | Sugar acids and derivatives |
| 22. | 7.6 | 164.16 | 163.0428 | 163.0432 | 2.5 | [M−H]− | 163.0, 119, 93, 65 | C9H8O3 | p-Coumaric acid | Hydroxycinnamic acids |
| 23. | 8.7 | 610.56 | 609.1817 | 609.1826 | 1.5 | [M−H]− | 609, 486, 301, 285,242, 151 | C28H34O15 | Hesperidin | Flavonoid glycosides |
| 24. | 8.8 | 302.19 | 300.9993 | 300.9989 | −1.3 | [M−H]− | 300.08, 283, 257.18,201, 207, 172 | C14H6O8 | Ellagic acid | Tannins |
| 25. | 9.9 | 194.18 | 193.0509 | 193.0512 | 1.6 | [M−H]− | 193, 178, 149, 134 | C10H10O4 | Isoferulic acid | Cinnamic acids |
| 26. | 9.95 | 194.18 | 193.0505 | 193.0503 | −1.0 | [M−H]− | 181, 164, 133, 118, 114.1, 104 | C10H10O4 | Ferulic acid | Cinnamic acids |
| 27. | 15.51 | 302.23 | 301.0359 | 301.0354 | −1.7 | [M−H]− | 301.08, 286, 273.08, 179, 151, 121,107 | C15H10O7 | Quercetin | Flavonoids |
| 28. | 16.7 | 286.23 | 285.0405 | 285.0406 | 0.4 | [M-H]- | 286, 247, 227, 219, 209 151, 142.97, 117 | C15H10O6 | Kaempferol | Flavanols |
| 29. | 16.8 | 270.24 | 269.045 | 269.0455 | 1.9 | [M−H]− | 269, 240, 225, 151, 117 | C15H10O5 | Apigenin | Flavonoids |
| 30. | 17.5 | 516.4 | 515.013 | 515.0128 | −0.4 | [M−H]− | 471, 394, 353, 341 327.25, 317, | C25H24O12 | 1,4-Dicaffeoylquinic acid | Quinic acids |
| 31. | 21.4 | 228.37 | 227.2018 | 227.2021 | 1.3 | [M−H]− | 227, 209, 67 | C14H28O2 | Myristic acid | Fatty acids |
| 32. | 22.6 | 430.71 | 431.3878 | 431.3883 | 1.2 | [M+H]+ | 432, 430, 205, 166, 165, 136, | C29H50O2 | α-Tocopherol | Fat-soluble vitamins |
| Analytes | Λ (nm) | Rt (min) | Linear Regression Data | LOD (µg/mL) | LOQ (µg/mL) | Concentration (µg/g) | Precision (RSD %) | Recovery | Analytes + Extract (µg/g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Range (µg/mL) | Equation | r2 | Inter Day | Intra Day | Mean | RSD% | 50 µg | 100 µg | ||||||
| Stigmasterol | 250 | 18.2 | 7.81−500 | y = 3.58x + 5.86 | 0.9999 | 0.63 | 1.93 | 243.66 | 1.11 | 1.67 | 99.55 ± 1.12 | 1.13 | 292.78 | 341.16 |
| β-sitosterol | 19.5 | 7.81−500 | y = 41.589x + 8.04 | 0.9998 | 0.56 | 1.70 | 317.04 | 0.44 | 0.68 | 99.20 ± 0.54 | 0.54 | 365.09 | 416.97 | |
| Scopoletin | 280 | 5.4 | 7.81−500 | y = 50.954x + 6.52 | 0.9997 | 0.44 | 1.33 | 217.40 | 0.97 | 1.60 | 99.23 ± 1.58 | 1.59 | 263.75 | 315.94 |
| 1,4-Dicaffeoylquinic acid | 7.2 | 7.81−500 | y = 42.941x + 8.99 | 0.9994 | 0.63 | 1.92 | 452.18 | 0.99 | 1.20 | 98.5 8± 0.46 | 0.47 | 499.45 | 550.91 | |
| Luteolin | 13.4 | 7.81−500 | y = 56.949x + 6.10 | 0.9999 | 0.31 | 0.94 | 617.17 | 0.99 | 0.40 | 98.60 ± 0.88 | 0.89 | 665.72 | 716.35 | |
| Kaempferol | 19.4 | 7.81−500 | y = 70.954x + 4.82 | 0.9999 | 0.20 | 0.61 | 783.02 | 1.67 | 1.42 | 100.00 ± 0.82 | 0.82 | 831.95 | 882.09 | |
| Ferulic acid | 24.9 | 7.81−500 | y = 58.902x + 5.68 | 0.9999 | 0.28 | 0.85 | 355.35 | 0.71 | 1.44 | 99.58 ± 1.13 | 1.14 | 404.45 | 444.85 | |
| Epicatechin | 320 | 4.7 | 7.81−500 | y = 51.969x + 6.77 | 0.9996 | 0.39 | 1.17 | 370.45 | 1.33 | 1.39 | 99.39 ± 1.51 | 1.51 | 418.70 | 469.41 |
| Ellagic acid | 7.4 | 7.81−500 | y = 77.965x + 8.34 | 0.9998 | 0.28 | 0.83 | 542.71 | 1.29 | 0.92 | 99.70 ± 0.48 | 0.48 | 591.11 | 641.78 | |
| Kaempferol-3-O-glucoside | 11.6 | 7.81−500 | y = 67.572x + 7.34 | 0.9999 | 0.32 | 0.96 | 457.81 | 0.48 | 1.28 | 99.95 ± 0.06 | 0.06 | 505.63 | 556.55 | |
| Quercetin | 13.8 | 7.81−500 | y = 61.58x + 4.48 | 0.9999 | 0.27 | 0.81 | 693.83 | 1.75 | 1.13 | 99.20 ± 0.87 | 0.88 | 742.80 | 792.49 | |
| Apigenin | 17.7 | 7.81−500 | y = 53.583x + 3.33 | 0.9999 | 0.31 | 0.93 | 578.93 | 0.73 | 0.89 | 99.53 ± 0.55 | 0.55 | 627.26 | 676.78 | |
| Parameters | 1,4-Dicaffeoylquinic Acid | Epicatechin | Ferulic Acid | Beta Sitosterol | Kaempferol-3-O-Glucoside | Ellagic Acid | Luteolin | Apigenin | Stigmasterol | Scopoletin | Quercetin | Kaempferol |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight (MW) | 516.457 | 290.272 | 194.187 | 414.713 | 448.382 | 302.197 | 286.24 | 270.241 | 412.698 | 192.171 | 302.24 | 286.24 |
| QPlogP o/w | 0.613 | 0.469 | 1.371 | 7.622 | −0.748 | −1.295 | 0.926 | 1.605 | 7.737 | 0.854 | 0.367 | 1.041 |
| QPlog S | −4.157 | −2.616 | −1.864 | −8.638 | −2.452 | −1.918 | −3.067 | −3.332 | −9.204 | −1.742 | −2.909 | −3.157 |
| QPlog HERG | −4.992 | −4.732 | −2.239 | −4.684 | −4.932 | −3.842 | −5.023 | −5.114 | −4.938 | −3.781 | −5.109 | −5.201 |
| QPP Caco | 0.259 | 53.201 | 63.536 | 3381.654 | 9.636 | 7.907 | 40.856 | 114.487 | 3379.988 | 632.797 | 18.199 | 51.24 |
| QPlog BB | −5.175 | −1.882 | −1.175 | −0.354 | −2.937 | −2.395 | −1.955 | −1.446 | −0.313 | −0.571 | −2.419 | −1.893 |
| QPP MDCK | 0.084 | 20.76 | 31.988 | 1846.188 | 3.275 | 2.645 | 15.606 | 47.531 | 1845.206 | 301.676 | 6.511 | 19.934 |
| QPlog Kp | −6.788 | −4.728 | −3.674 | −1.651 | −5.636 | −6.701 | −4.888 | −3.989 | −1.747 | −3.065 | −5.544 | −4.641 |
| VDss (human) | 1.96 | 1.027 | −1.367 | 0.193 | 1.444 | 0.375 | 1.153 | 0.822 | 0.178 | 0.034 | 1.559 | 1.274 |
| CNS permeability | −3.804 | −3.298 | −2.612 | −1.705 | −3.908 | −3.533 | −2.251 | −2.061 | −1.652 | −2.32 | −3.065 | −2.228 |
| Bioavailability Score | - | 0.55 | 0.85 | - | 0.17 | 0.55 | 0.55 | 0.55 | - | 0.55 | 0.55 | 0.55 |
| QPlog Khsa | −0.738 | −0.412 | −0.612 | 2.077 | −0.751 | −0.658 | −0.198 | −0.039 | 2.169 | −0.481 | −0.343 | −0.191 |
| Percent Human Oral Absorption | 0 | 60.584 | 67.241 | 100 | 14.261 | 35.438 | 61.205 | 73.192 | 100 | 82.085 | 51.649 | 63.637 |
| GI absorption | Low | High | High | High | Low | High | High | High | High | High | High | High |
| P-glycoprotein substrate | Yes | Yes | No | No | No | No | No | No | No | No | No | No |
| P-glycoprotein I inhibitor | No | No | No | Yes | No | No | No | No | Yes | No | No | No |
| P-glycoprotein II inhibitor | No | No | No | Yes | No | No | No | No | Yes | No | No | No |
| Primary metabolites | 6 | 7 | 2 | 3 | 7 | 4 | 4 | 3 | 5 | 2 | 5 | 4 |
| CYP1A2 inhibitor | No | No | No | No | No | Yes | Yes | Yes | No | Yes | Yes | Yes |
| CYP2C19 inhibitor | No | No | No | No | No | No | No | No | No | No | No | No |
| CYP2C9 inhibitor | No | No | No | No | No | No | No | No | No | No | No | No |
| CYP2D6 inhibitor | No | No | No | No | No | No | Yes | Yes | No | No | Yes | Yes |
| CYP3A4 inhibitor | Yes | No | No | Yes | No | No | Yes | Yes | Yes | No | Yes | Yes |
| Total Clearance (mL/min/kg) | −0.062 | 0.183 | 0.623 | 0.628 | 0.462 | 0.537 | 0.495 | 0.566 | 0.618 | 0.73 | 0.407 | 0.477 |
| Renal OCT2 substrate | No | No | No | No | No | No | No | No | No | No | No | No |
| AMES toxicity | No | No | No | No | No | No | No | No | No | No | No | No |
| Oral Rat Acute Toxicity (LD50) | 2.543 | 2.428 | 2.282 | 2.552 | 2.546 | 2.399 | 2.455 | 2.45 | 2.54 | 1.95 | 2.471 | 2.449 |
| Oral Rat Chronic Toxicity (LOAEL) (mg/kg/day) | 3.889 | 2.5 | 2.065 | 0.855 | 4.53 | 2.698 | 2.409 | 2.298 | 0.872 | 1.378 | 2.612 | 2.505 |
| Hepatotoxicity | No | No | No | No | No | No | No | No | No | No | No | No |
| Skin Sensitization | No | No | No | No | No | No | No | No | No | No | No | No |
| Lipinski violations | - | 0 | 0 | - | 2 | 0 | 0 | 0 | - | 0 | 0 | 0 |
| Ghose violations | - | 0 | 0 | - | 0 | 0 | 0 | 0 | - | 0 | 0 | 0 |
| Veber violations | - | 0 | 0 | - | 1 | 1 | 0 | 0 | - | 0 | 0 | 0 |
| Egan violations | - | 0 | 0 | - | 1 | 1 | 0 | 0 | - | 0 | 0 | 0 |
| Muegge violations | - | 0 | 1 | - | 3 | 0 | 0 | 0 | - | 1 | 0 | 0 |
| Name | Docking Score | ∆ GBinding | Log Ki (µMolar) | ∆ GCoulomb | ∆ GCovalent | ∆ GHbond | ∆ GLipophilic | ∆GSolv GB | ∆ GvdW | Residue-Ligand Interactions with Distance (Å) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hydrogen Bonds | Electrostatic/Hydrophobic Bonds | ||||||||||
| Voltage-gated calcium channel beta2a (VGCC, PDB:1T0J) | |||||||||||
| Kaempferol-3-O-glucoside | −5.21 | −36.36 | −12.56 | −19.31 | 0.93 | −3.68 | −8.18 | 28.36 | −33.52 | Conventional Hydrogen Bond: Lys254 (1.80), Arg424 (3.04), Asp251 (1.90), Asp319 (1.74), Ile261 (2.88) Carbon Hydrogen Bond: Lys254 (3.03), Asp251 (2.91), Asp319 (3.06) | πAlkyl: Ile263 (4.40), Ile263 (4.75), Arg265 (5.41) |
| Luteolin | −5.181 | −29.94 | −9.77 | −23.33 | 1.24 | −1.53 | −3.41 | 22.59 | −24.63 | Conventional Hydrogen Bond: Arg227 (2.41), Val109 (1.78), Glu381 (1.73) | Electrostatic π Anion Asp384 (4.00), Asp384 (3.53) π−π T-shaped: Tyr402 (5.79) |
| Ellagic acid | −4.806 | −35.03 | −11.98 | −22.11 | 1.12 | −1.51 | −11.31 | 26.81 | −27.61 | Conventional Hydrogen Bond: Phe92 (1.95), Arg227 (1.98), Tyr40 (2.89) Carbon Hydrogen Bond: Asp91 (2.87), Arg227 (2.99) | π−Lone pair: Val109 (2.80) π−π Stacked: Tyr402 (5.40) πAlkyl: Lys110 (4.87), Val109 (5.29), Lys110 (5.49), Ala405 (5.33) |
| Kaempferol | −3.822 | −34.67 | −11.83 | −20.81 | 0.31 | −1.36 | −6.91 | 20.73 | −23.84 | Conventional Hydrogen Bond: Asp91 (2.73), Glu381 (1.81), Val109 (1.75) | π−π T-shaped: Phe92 (5.30) |
| Quercetin | −3.421 | −26.09 | −8.10 | −15.86 | 0.46 | −3.5 | −4.44 | 16.35 | −18.58 | Conventional Hydrogen Bond: Lys247 (2.09), Lys254 (1.93), Arg265 (1.96), Arg424 (2.14), Asp319 (1.71) | π−Alkyl: Lys247 (5.30), Ile263 (5.49) |
| Verapamil | −2.351 | −42.52 | −15.24 | −16.52 | 1.27 | −1.14 | −14.39 | 33.32 | −44.34 | Conventional Hydrogen Bond: Arg227 (2.65) Carbon Hydrogen Bond: Tyr402 (2.72), Asp384 (2.54), Tyr402 (2.69), Glu111 (2.61), Ser330 (2.75), Pro336 (2.49), Glu381 (2.67), Ser382 (2.43) | Electrostatic Attractive Charge:Asp384 (4.51) π−Cation: Arg227 (4.00) π−Anion: Asp384 (3.66) Alkyl: Ala335 (4.32), Ala405 (3.99), Lys110 (4.24), Pro326 (5.46), Ile338 (4.46) π−Alkyl: Phe92 (5.37), Lys110 (5.11), Ala409 (4.44)) |
| Calcium/calmodulin-dependent protein kinase IIB (CAMK2B, PDB: 3BHH) | |||||||||||
| Kaempferol-3-O-Glucoside | −6.39 | −41.23 | −14.68 | −39.64 | 3.62 | −6.08 | −6.07 | 43.77 | −32.21 | Conventional Hydrogen Bond: Arg187 (1.91), Asn256 (2.07), Lys138 (2.17), Lys227 (1.95), Lys227 (1.99), Glu189 (1.78), Glu140 (1.86) Carbon Hydrogen Bond: Glu189 (2.82), Glu189 (2.62) | Electrostatic π−Cation: Arg298 (4.27), Arg298 (3.18) |
| Kaempferol | −5.72 | −36.84 | −12.77 | −22.11 | 4.46 | −1.9 | −10.67 | 25.72 | −26.29 | Conventional Hydrogen Bond: Trp215 (1.86), Glu217 (1.74), Phe294 (2.13) πDonor Hydrogen Bond: Trp215 (2.44) | Electrostatic π−Cation: Arg66 (3.87), Arg298 (4.37) π−π T-Shaped: Trp215 (4.32) π−Alkyl: Arg297 (4.94), Arg298 (4.83) |
| Ellagic Acid | −5.231 | −18.31 | −4.72 | −26.11 | 2.2 | −4.38 | −2.8 | 36.58 | −23.57 | Conventional Hydrogen Bond: Asn256 (2.69), Arg66 (2.59), Arg297 (2.09), Glu59 (2.02), Leu300 (2.02), Glu59 (1.89), Leu300 (2.79) Carbon Hydrogen Bond: Arg297 (2.66) | Electrostatic π−Cation: Lys259 (4.40), Lys259 (3.50 Electrostatic π−Anion: Glu83 (3.77), Glu83 (3.94), Glu83 (3.29 |
| Quercetin | −4.999 | −30.78 | −10.14 | −29.96 | 3.95 | −3.98 | −4.18 | 36.29 | −27.17 | Conventional Hydrogen Bond: Arg66 (2.06), Glu217 (1.70), Glu217 (1.72), Glu59 (2.16) Carbon Hydrogen Bond: Arg297 (2.71) | Electrostatic π−Cation: Lys259 (4.03), Arg66 (3.66) π−π T-Shaped: Trp215 (5.10), Trp215 (5.38), Trp215 (4.95) |
| Luteolin | −4.885 | −30.39 | −9.97 | −24.26 | 1.96 | −3.59 | −4.59 | 28.74 | −28.33 | Conventional Hydrogen Bond: Lys293 (1.95), Lys293 (2.19), Carbon Hydrogen Bond: Arg298 (2.87) π −Donor Hydrogen Bond: Arg66 (3.77), Arg66 (2.96) | Electrostatic Attractive Charge: Arg66 (3.83), Electrostatic π−Cation: Arg66 (3.77), Arg66 (3.00) π−π Stacked: Phe294 (5.13) π−π T-Shaped: Trp215 (4.92), Trp215 (4.96) Amide−π Stacked: Phe214 (4.95), Trp215 (4.95) π−Alkyl: Pro212 (4.15), Pro212 (4.38), Arg297 (4.71), Lys69 (4.40), Pro212 (5.36) |
| Verapamil | −1.537 | −36.04 | −12.42 | 26.1 | −0.27 | −1.06 | −15.28 | 1.48 | −44.54 | Carbon Hydrogen Bond: Arg298 (2.82), Glu217 (2.48), Asp216 (2.53), Glu82 (2.98), Glu82 (2.99) π −Donor Hydrogen Bond: Asn256 (2.93) | Electrostatic π−Cation: Arg298 (3.44) π−π T-Shaped: Trp215 (4.88) Alkyl: Ala258 (4.48), Pro212 (3.94) π−Alkyl: Ala258 (5.05) |
| Myosin light chain kinase-1 (MLCK-1, PDB: 6C6M) | |||||||||||
| Quercetin | −6.336 | −36.92 | −12.81 | −35.65 | 4.07 | −2.65 | −9.55 | 31.81 | −23.81 | Conventional Hydrogen Bond: Arg456 (2.20), His470 (1.86), Glu436 (1.61), Trp447 (1.56) | π−Alkyl: Val463 (4.66), Val463 (5.10), Ala446 (4.26), Val463 (5.36) |
| Luteolin | −5.319 | −28.79 | −9.27 | −23.92 | 4.03 | −2.44 | −3.17 | 26.21 | −28.28 | Conventional Hydrogen Bond: Trp447 (1.94), Arg456 (2.09), Val454 (2.07), Glu465 (2.64) Carbon Hydrogen Bond: Trp447 (2.88), Arg456 (3.09) | π−Alkyl: Val463 (4.73), Val463 (4.02) |
| Kaempferol | −5.196 | −42.83 | −15.37 | −34.11 | 2.02 | −2.16 | −9.6 | 30.55 | −28.39 | Conventional Hydrogen Bond: Arg456 (2.18), His470 (1.85), Trp447 (1.66) π −Donor Hydrogen Bond: Trp447 (2.92) | π−Alkyl: Val463 (4.56), Val463 (5.03), Ala446 (4.16), Val463 (5.44) |
| Kaempferol-3-O-Glucoside | −5.184 | −37.68 | −13.14 | −52.04 | 8.12 | −4.28 | −8.2 | 51.95 | −32.55 | Conventional Hydrogen Bond: Arg456 (2.95), Arg456 (1.82), Arg456 (2.60), Arg456 (2.32), Arg456 (2.16), Val445 (1.77), His470 (1.89), Glu465 (1.79) Carbon Hydrogen Bond: Glu444 (2.50), Val445 (2.53), Val445 (2.62), His470 (2.90), Val445 (2.63) π −Donor Hydrogen Bond: Glu465 (2.76) | π−σ: His470 (2.83) π−π T-Shaped: His470 (5.02) |
| Ellagic Acid | −4.453 | −32.82 | −11.02 | −9.37 | 1.84 | −1.53 | −9.78 | 18.6 | −30.41 | Conventional Hydrogen Bond: Trp447 (1.91) Carbon Hydrogen Bond: Ala446 (2.53), Trp447 (2.34), Trp447 (2.44) | Electrostatic π−Cation: Arg456 (3.78), Arg456 (4.38) Electrostatic π−Anion: Glu444 (4.67) π−Alkyl: Val463 (3.97), Val463 (5.26), Val463 (4.57), Val463 (5.38) |
| Verapamil | −2.844 | −33.73 | −11.42 | 7.71 | 7.06 | −1.13 | −14.36 | 4.17 | −36.53 | Conventional Hydrogen Bond: Arg456 (2.49), Arg456 (2.03) Carbon Hydrogen Bond: Leu449 (2.93), Glu450 (2.89), Glu465 (2.55), Glu465 (2.59), Asp481 (2.54), Pro453 (2.63) π −Donor Hydrogen Bond: Arg480 (4.17) | Electrostatic π−Cation: Arg480 (4.17) Alkyl: Leu449 (4.89), Val454 (4.56), Ile461 (5.25) |
| Phosphoinositide Phospholipase C-Gamma-1 (PLCγ1, PDB: 4EY0) | |||||||||||
| Quercetin | −9.119 | −25.63 | −7.90 | −15.74 | 2.03 | −2.81 | −10.72 | 35.42 | −32.32 | Conventional Hydrogen Bond: Ser612 (2.57), Glu548 (2.20), Glu548 (1.81), Tyr595 (2.79) Carbon Hydrogen Bond: Ser631 (3.00) | π−π T-Shaped: Phe621 (5.46), Phe621 (5.47) π−Alkyl: Leu632 (5.19), Pro619 (4.48), Pro619 (4.14) |
| Luteolin | −8.941 | −29.04 | −9.38 | −21.27 | 2.61 | −3.06 | −9.3 | 36.96 | −32.96 | Conventional Hydrogen Bond: Ser612 (2.11), Glu548 (1.93), Glu548 (1.99) | π−π T-Shaped: Phe621 (5.57), Phe621 (5.55) π−Alkyl: Leu632 (5.37), Leu632 (5.39), Pro619 (4.14), Pro619 (4.63) |
| Kaempferol | −8.159 | −25.09 | −7.67 | −16.35 | 1.09 | −2.55 | −10.81 | 36.25 | −30.94 | Conventional Hydrogen Bond: Tyr595 (2.30), Glu548 (2.42), Glu548 (1.84) | π−π T-Shaped: Phe621 (5.48), Phe621 (5.33) π−Alkyl: Leu632 (5.18), Pro619 (4.76), Pro619 (3.99) |
| Ellagic Acid | −5.668 | −44.22 | −15.98 | −26.45 | 2.36 | −4.11 | −11.4 | 25.55 | −26.49 | Conventional Hydrogen Bond: Asp630 (2.14), Val628 (2.24), Glu667 (2.01), Asp634 (1.92), Glu667 (1.75) | π−π T-Shaped: Phe629 (5.23), Phe629 (4.78) π−Alkyl: Leu627 (5.49), Leu627 (4.77) |
| Kaempferol-3-O-Glucoside | −5.566 | −36.39 | −12.57 | −31.94 | 2.91 | −4.4 | −8.27 | 45.71 | −37.77 | Conventional Hydrogen Bond: Lys666 (1.96), Asp630 (1.83), Asp634 (1.74), Val628 (2.98), Glu667 (1.99) Carbon Hydrogen Bond: Asp634 (2.75) | π−π T-Shaped: Phe629 (5.10), His638 (5.10) Amide−π Stacked: Thr637 (4.75), His638 (4.75) π−Alkyl: Leu627 (4.80) |
| Verapamil | −4.069 | −45.13 | −16.37 | −46.51 | 10.09 | −0.67 | −24.7 | 58.56 | −39.94 | Conventional Hydrogen Bond: Leu632 (2.24) Carbon Hydrogen Bond: Pro619 (2.84), Ser631 (2.66), Ser631 (2.53), Tyr595 (2.34), Gln614 (2.76), Gly617 (2.93), Asp630 (2.71), Glu548 (2.59), Glu548 (2.66), Glu548 (2.84) | π−π T-Shaped: Phe621 (5.22) Alkyl: Pro619 (4.38), Pro619 (4.85) π−Alkyl: Phe551 (4.95), Phe621 (5.37), Pro619 (4.64), Leu632 (4.70) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahid, M.; Saqib, F.; Ahmedah, H.T.; Gavris, C.M.; De Feo, V.; Hogea, M.; Moga, M.; Chicea, R. Cucumis sativus L. Seeds Ameliorate Muscular Spasm-Induced Gastrointestinal and Respiratory Disorders by Simultaneously Inhibiting Calcium Mediated Signaling Pathway. Pharmaceuticals 2021, 14, 1197. https://doi.org/10.3390/ph14111197
Wahid M, Saqib F, Ahmedah HT, Gavris CM, De Feo V, Hogea M, Moga M, Chicea R. Cucumis sativus L. Seeds Ameliorate Muscular Spasm-Induced Gastrointestinal and Respiratory Disorders by Simultaneously Inhibiting Calcium Mediated Signaling Pathway. Pharmaceuticals. 2021; 14(11):1197. https://doi.org/10.3390/ph14111197
Chicago/Turabian StyleWahid, Muqeet, Fatima Saqib, Hanadi Talal Ahmedah, Claudia Mihaela Gavris, Vincenzo De Feo, Mircea Hogea, Marius Moga, and Radu Chicea. 2021. "Cucumis sativus L. Seeds Ameliorate Muscular Spasm-Induced Gastrointestinal and Respiratory Disorders by Simultaneously Inhibiting Calcium Mediated Signaling Pathway" Pharmaceuticals 14, no. 11: 1197. https://doi.org/10.3390/ph14111197
APA StyleWahid, M., Saqib, F., Ahmedah, H. T., Gavris, C. M., De Feo, V., Hogea, M., Moga, M., & Chicea, R. (2021). Cucumis sativus L. Seeds Ameliorate Muscular Spasm-Induced Gastrointestinal and Respiratory Disorders by Simultaneously Inhibiting Calcium Mediated Signaling Pathway. Pharmaceuticals, 14(11), 1197. https://doi.org/10.3390/ph14111197