
STW 5 Herbal Preparation Modulates Wnt3a and Claudin 1 Gene Expression in Zebrafish IBS-like Model



 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
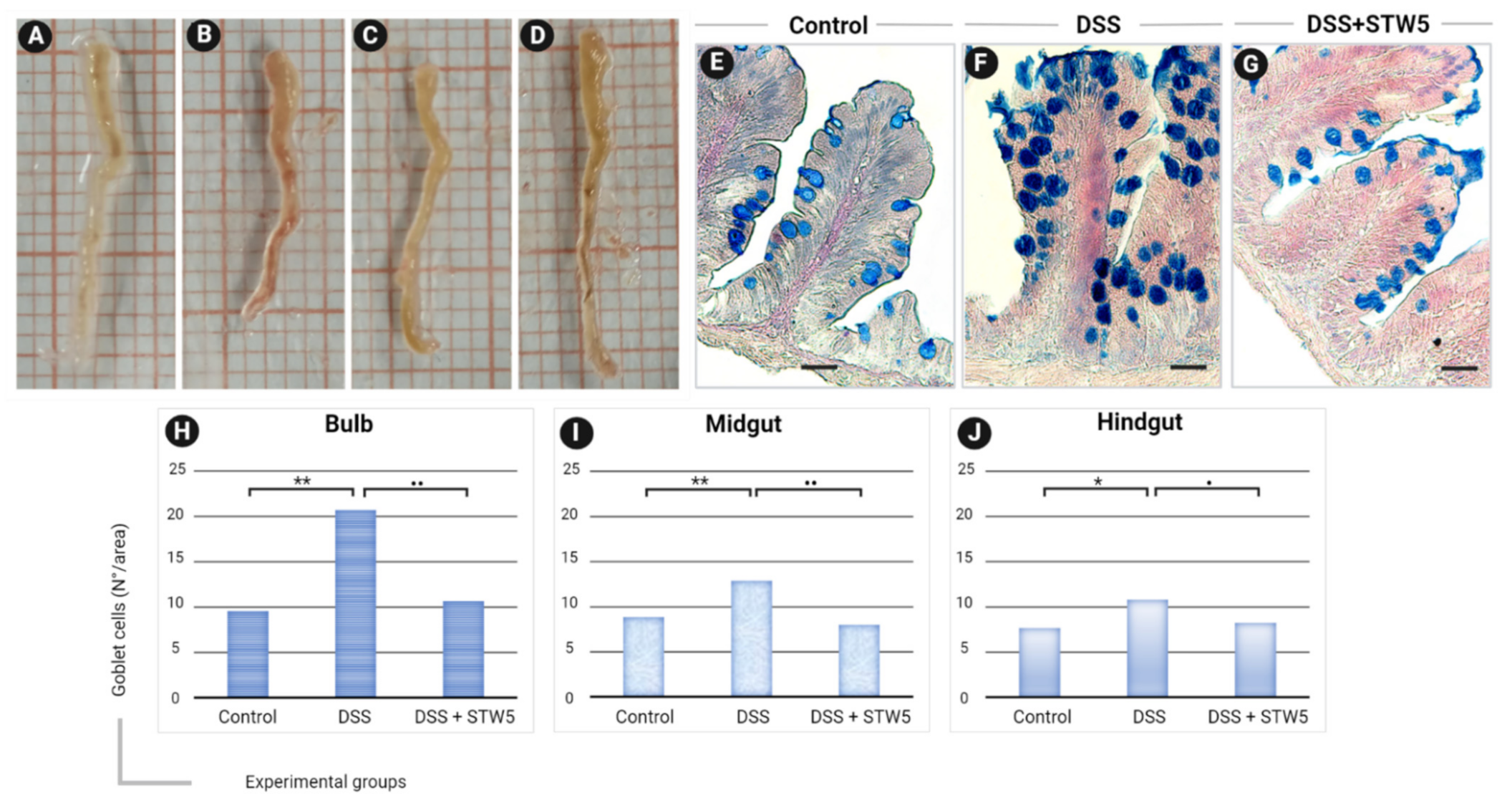
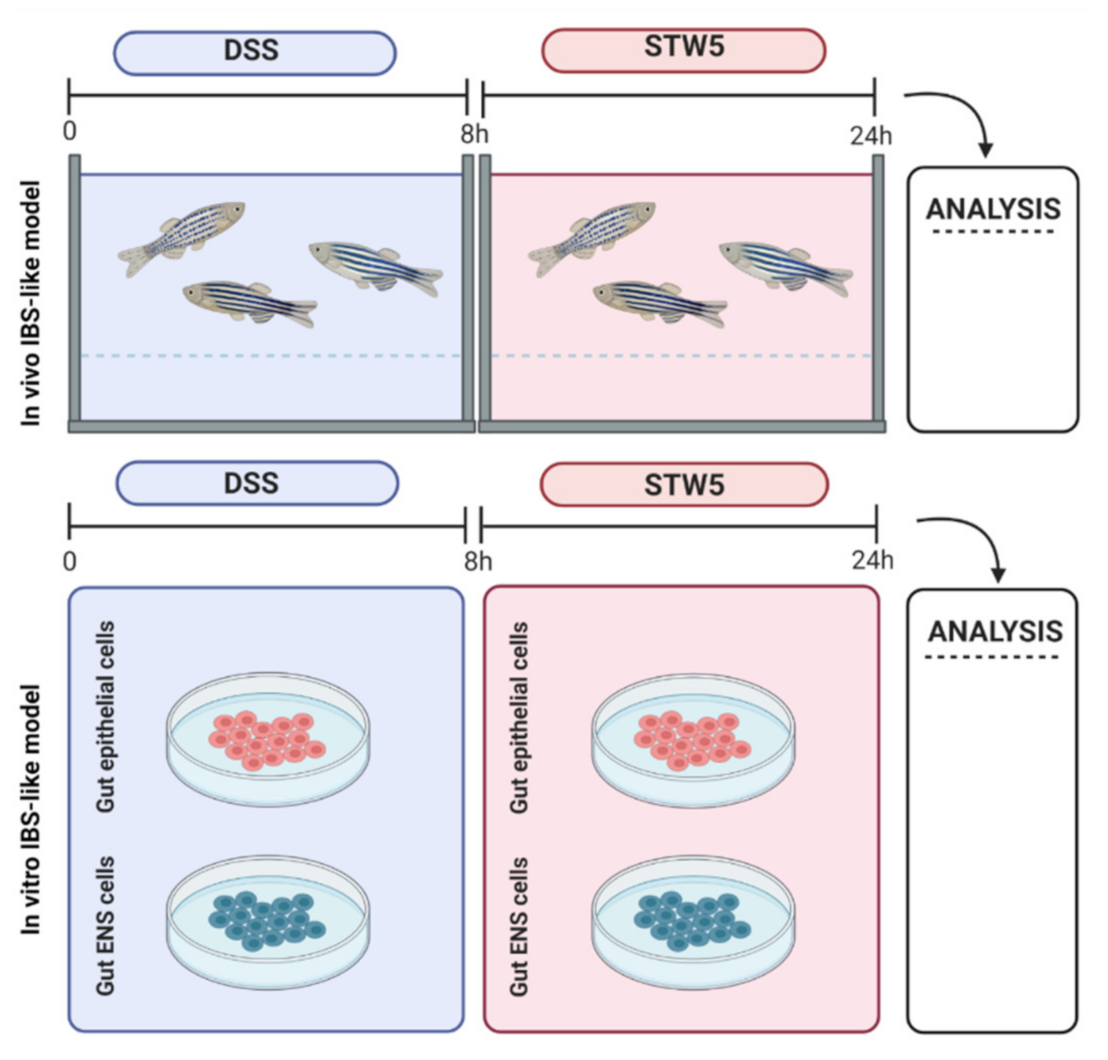
2.1. In Vivo Model of IBS and Impact of STW 5 on Morphological Changes
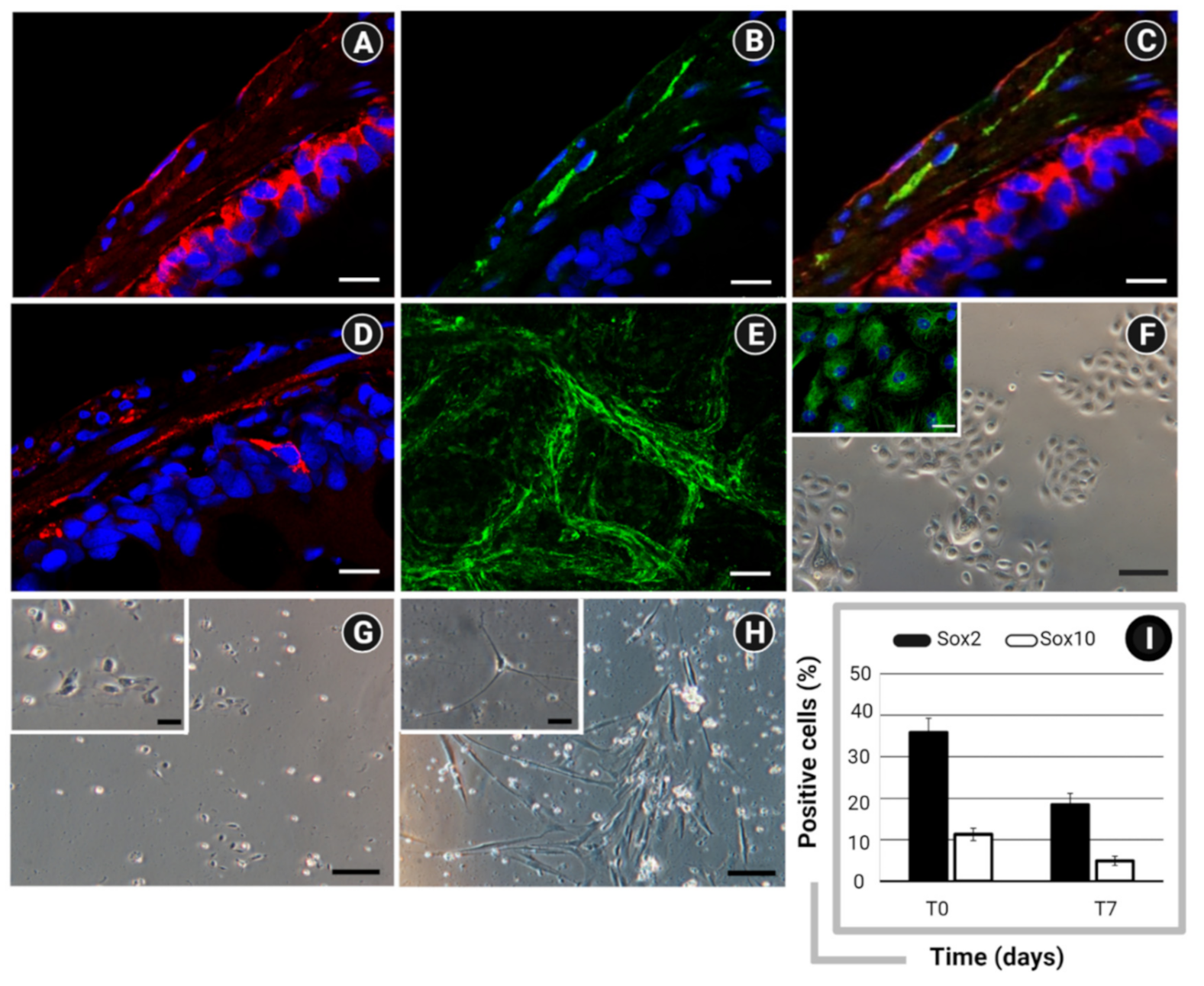
2.2. In Vitro Models of IBS
2.3. In Vivo Response to STW 5
2.4. In Vitro Molecular Efficacy of STW 5
3. Discussion
4. Materials and Methods
4.1. Herbal Preparation
4.2. Animals
4.3. Investigation of Canonical Wnt Components in Zebrafish Gut
4.4. Assessment of Zebrafish Models of IBS
4.4.1. In Vivo Model
Histochemistry
qRT-PCR
4.4.2. In Vitro Models
EP Cell Cultures
Whole-Mount Immunofluorescent Staining
ENS Cell Cultures
Morphological and Immunophenotypic Characterization of EPc and ENSc
Impact of STW 5 on EPc and ENSc
4.4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Drossman, D.A. Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features and Rome IV. Gastroenterology 2016, 150, 1262–1279.e2. [Google Scholar] [CrossRef] [Green Version]
- Enck, P.; Aziz, Q.; Barbara, G.; Farmer, A.D.; Fukudo, S.; Mayer, E.A.; Niesler, B.; Quigley, E.M.; Rajilić-Stojanović, M.; Schemann, M.; et al. Irritable bowel syndrome. Nat. Rev. Dis. Primers 2016, 2, 16014. [Google Scholar] [CrossRef] [Green Version]
- Schmulson, M.J.; Drossman, D.A. What Is New in Rome IV. J. Neurogastroenterol. Motil. 2017, 23, 151–163. [Google Scholar] [CrossRef]
- Lacy, B.E.; Chey, W.D.; Lembo, A.J. New and Emerging Treatment Options for Irritable Bowel Syndrome. Gastroenterol. Hepatol. 2015, 11, 1–19. [Google Scholar]
- Lembo, A.J.; Lacy, B.E.; Zuckerman, M.J.; Schey, R.; Dove, L.S.; Andrae, D.A.; Davenport, J.M.; McIntyre, G.; Lopez, R.; Turner, L.; et al. Eluxadoline for Irritable Bowel Syndrome with Diarrhea. N. Engl. J. Med. 2016, 374, 242–253. [Google Scholar] [CrossRef]
- Chadwick, V.S.; Chen, W.; Shu, D.; Paulus, B.; Bethwaite, P.; Tie, A.; Wilson, I. Activation of the mucosal immune system in irritable bowel syndrome. Gastroenterology 2002, 122, 1778–1783. [Google Scholar] [CrossRef]
- Stasi, C.; Rosselli, M.; Bellini, M.; Laffi, G.; Milani, S. Altered neuro-endocrine-immune pathways in the irritable bowel syndrome: The top-down and the bottom-up model. J. Gastroenterol. 2012, 47, 1177–1185. [Google Scholar] [CrossRef]
- Hughes, P.A.; Harrington, A.M.; Castro, J.; Liebregts, T.; Adam, B.; Grasby, D.J.; Isaacs, N.J.; Maldeniya, L.; Martin, C.M.; Persson, J.; et al. Sensory neuro-immune interactions differ between irritable bowel syndrome subtypes. Gut 2013, 62, 1456–1465. [Google Scholar] [CrossRef]
- Hughes, K.R.; Sablitzky, F.; Mahida, Y.R. Expression profiling of Wnt family of genes in normal and inflammatory bowel disease primary human intestinal myofibroblasts and normal human colonic crypt epithelial cells. Inflamm. Bowel Dis. 2011, 17, 213–220. [Google Scholar] [CrossRef]
- Ford, A.C.; Moayyedi, P.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.; Quigley, E.M.; The Task Force on the Management of Functional Bowel Disorders. American College of Gastroenterology monograph on the management of irritable bowel syndrome and chronic idiopathic constipation. Am. J. Gastroenterol. 2014, 109 (Suppl. 1), S2–S26. [Google Scholar] [CrossRef]
- Chey, W.D.; Kurlander, J.; Eswaran, S. Irritable bowel syndrome: A clinical review. JAMA 2015, 313, 949–958. [Google Scholar] [CrossRef]
- Liu, J.P.; Yang, M.; Liu, Y.X.; Wei, M.; Grimsgaard, S. Herbal medicines for treatment of irritable bowel syndrome. Cochrane Database Syst. Rev. 2006, CD004116. [Google Scholar] [CrossRef]
- Holtmann, G.; Schrenk, D.; Madisch, A.; Allescher, H.D.; Ulrich-Merzenich, G.; Mearin, F.; Larrey, D.; Malfertheiner, P. Use of Evidence-Based Herbal Medicines for Patients with Functional Gastrointestinal Disorders: A Conceptional Framework for Risk-Benefit Assessment and Regulatory Approaches. Dig. Dis. 2020, 38, 269–279. [Google Scholar] [CrossRef]
- Rösch, W.; Vinson, B.; Sassin, I. A randomised clinical trial comparing the efficacy of a herbal preparation STW 5 with the prokinetic drug cisapride in patients with dysmotility type of functional dyspepsia. Z. Gastroenterol. 2002, 40, 401–408. [Google Scholar] [CrossRef]
- Madisch, A.; Holtmann, G.; Mayr, G.; Vinson, B.; Hotz, J. Treatment of functional dyspepsia with a herbal preparation. A double-blind, randomized, placebo-controlled, multicenter trial. Digestion 2004, 69, 45–52. [Google Scholar] [CrossRef]
- Madisch, A.; Holtmann, G.; Plein, K.; Hotz, J. Treatment of irritable bowel syndrome with herbal preparations: Results of a double-blind, randomized, placebo-controlled, multi-centre trial. Aliment. Pharmacol. Ther. 2004, 19, 271–279. [Google Scholar] [CrossRef]
- Raedsch, R.; Hanisch, J.; Bock, P.; Sibaev, A.; Vinson, B.; Gundermann, K.J. Assessment of the efficacy and safety of the phytopharmacon STW 5 versus metoclopramide in functional dyspepsia—A retrolective cohort study. Z. Gastroenterol. 2007, 45, 1041–1048. [Google Scholar] [CrossRef]
- von Arnim, U.; Peitz, U.; Vinson, B.; Gundermann, K.J.; Malfertheiner, P. STW 5, a phytopharmacon for patients with functional dyspepsia: Results of a multicenter, placebo-controlled double-blind study. Am. J. Gastroenterol. 2007, 102, 1268–1275. [Google Scholar] [CrossRef]
- Braden, B.; Caspary, W.; Börner, N.; Vinson, B.; Schneider, A.R. Clinical effects of STW 5 (Iberogast) are not based on acceleration of gastric emptying in patients with functional dyspepsia and gastroparesis. Neurogastroenterol. Motil. 2009, 21, 632–638. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.; Kelber, O.; Lorkowski, G.; Storr, M. Evaluating the Multitarget Effects of Combinations through Multistep Clustering of Pharmacological Data: The Example of the Commercial Preparation Iberogast. Planta Med. 2017, 83, 1130–1140. [Google Scholar] [CrossRef]
- Allescher, H.D.; Abdel-Aziz, H. Mechanism of Action of STW 5 in Functional Dyspepsia and IBS: The Origin of Multi-Target. Dig. Dis. 2017, 35 (Suppl. 1), 18–24. [Google Scholar] [CrossRef] [Green Version]
- Allescher, H.D.; Wagner, H. STW 5/Iberogast: Multi-target-action for treatment of functional dyspepsia and irritable bowel syndrome. Wien. Med. Wochenschr. 2007, 157, 301–307. [Google Scholar] [CrossRef]
- Ottillinger, B.; Storr, M.; Malfertheiner, P.; Allescher, H.D. STW 5 (Iberogast®)--a safe and effective standard in the treatment of functional gastrointestinal disorders. Wien. Med. Wochenschr. 2013, 163, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Malfertheiner, P. STW 5 (Iberogast) Therapy in Gastrointestinal Functional Disorders. Dig. Dis. 2017, 35 (Suppl. 1), 25–29. [Google Scholar] [CrossRef]
- Di Liddo, R.; Bertalot, T.; Schuster, A.; Schrenk, S.; Tasso, A.; Zanusso, I.; Conconi, M.T.; Schäfer, K.H. Anti-inflammatory activity of Wnt signaling in enteric nervous system: In vitro preliminary evidences in rat primary cultures. J. Neuroinflamm. 2015, 12, 23. [Google Scholar] [CrossRef] [Green Version]
- Le Guen, L.; Marchal, S.; Faure, S.; de Santa Barbara, P. Mesenchymal-epithelial interactions during digestive tract development and epithelial stem cell regeneration. Cell Mol. Life Sci. 2015, 72, 3883–3896. [Google Scholar] [CrossRef] [Green Version]
- Simonetti, M.; Agarwal, N.; Stösser, S.; Bali, K.K.; Karaulanov, E.; Kamble, R.; Pospisilova, B.; Kurejova, M.; Birchmeier, W.; Niehrs, C.; et al. Wnt-Fzd signaling sensitizes peripheral sensory neurons via distinct noncanonical pathways. Neuron 2014, 83, 104–121. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M. WNT signaling in stem cell biology and regenerative medicine. Curr. Drug Targets 2008, 9, 565–570. [Google Scholar] [CrossRef]
- Ratanasirintrawoot, S.; Israsena, N. Stem Cells in the Intestine: Possible Roles in Pathogenesis of Irritable Bowel Syndrome. J. Neurogastroenterol. Motil. 2016, 22, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Neumann, J.; Schaale, K.; Farhat, K.; Endermann, T.; Ulmer, A.J.; Ehlers, S.; Reiling, N. Frizzled1 is a marker of inflammatory macrophages, and its ligand Wnt3a is involved in reprogramming Mycobacterium tuberculosis-infected macrophages. FASEB J. 2010, 24, 4599–4612. [Google Scholar] [CrossRef]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef]
- Manicassamy, S.; Reizis, B.; Ravindran, R.; Nakaya, H.; Salazar-Gonzalez, R.M.; Wang, Y.C.; Pulendran, B. Activation of beta-catenin in dendritic cells regulates immunity versus tolerance in the intestine. Science 2010, 329, 849–853. [Google Scholar] [CrossRef] [Green Version]
- Di Liddo, R.; Bertalot, T.; Schuster, A.; Schrenk, S.; Müller, O.; Apfel, J.; Reischmann, P.; Rajendran, S.; Sfriso, R.; Gasparella, M.; et al. Fluorescence-based gene reporter plasmid to track canonical Wnt signaling in ENS inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G337–G346. [Google Scholar] [CrossRef] [Green Version]
- Sen, M.; Ghosh, G. Transcriptional outcome of Wnt-Frizzled signal transduction in inflammation: Evolving concepts. J. Immunol. 2008, 181, 4441–4445. [Google Scholar] [CrossRef] [Green Version]
- Jridi, I.; Canté-Barrett, K.; Pike-Overzet, K.; Staal, F.J.T. Inflammation and Wnt Signaling: Target for Immunomodulatory Therapy? Front. Cell Dev. Biol. 2020, 8, 615131. [Google Scholar] [CrossRef]
- Yang, Y.; Tomkovich, S.; Jobin, C. Could a swimming creature inform us on intestinal diseases? Lessons from zebrafish. Inflamm. Bowel Dis. 2014, 20, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Scanzi, J.; Accarie, A.; Muller, E.; Pereira, B.; Aissouni, Y.; Goutte, M.; Joubert-Zakeyh, J.; Picard, E.; Boudieu, L.; Mallet, C.; et al. Colonic overexpression of the T-type calcium channel Ca. Neurogastroenterol. Motil. 2016, 28, 1632–1640. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M.; Carlson, P.; Valentin, N.; Acosta, A.; O’Neill, J.; Eckert, D.; Dyer, R.; Na, J.; Klee, E.W.; Murray, J.A. Pilot study of small bowel mucosal gene expression in patients with irritable bowel syndrome with diarrhea. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G365–G376. [Google Scholar] [CrossRef] [Green Version]
- D’Antongiovanni, V.; Pellegrini, C.; Fornai, M.; Colucci, R.; Blandizzi, C.; Antonioli, L.; Bernardini, N. Intestinal epithelial barrier and neuromuscular compartment in health and disease. World J. Gastroenterol. 2020, 26, 1564–1579. [Google Scholar] [CrossRef]
- You, J.; Nguyen, A.V.; Albers, C.G.; Lin, F.; Holcombe, R.F. Wnt pathway-related gene expression in inflammatory bowel disease. Dig. Dis. Sci. 2008, 53, 1013–1019. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Dharmani, P.; Leung, P.; Chadee, K. Tumor necrosis factor-α and Muc2 mucin play major roles in disease onset and progression in dextran sodium sulphate-induced colitis. PLoS ONE 2011, 6, e25058. [Google Scholar] [CrossRef]
- Cheng, P.; Yao, J.; Wang, C.; Zhang, L.; Kong, W. Molecular and cellular mechanisms of tight junction dysfunction in the irritable bowel syndrome. Mol. Med. Rep. 2015, 12, 3257–3264. [Google Scholar] [CrossRef] [Green Version]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Graham, V.; Khudyakov, J.; Ellis, P.; Pevny, L. SOX2 functions to maintain neural progenitor identity. Neuron 2003, 39, 749–765. [Google Scholar] [CrossRef] [Green Version]
- Heanue, T.A.; Pachnis, V. Prospective identification and isolation of enteric nervous system progenitors using Sox2. Stem Cells 2011, 29, 128–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laranjeira, C.; Sandgren, K.; Kessaris, N.; Richardson, W.; Potocnik, A.; Vanden Berghe, P.; Pachnis, V. Glial cells in the mouse enteric nervous system can undergo neurogenesis in response to injury. J. Clin. Investig. 2011, 121, 3412–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colomier, E.; Algera, J.; Melchior, C. Pharmacological Therapies and Their Clinical Targets in Irritable Bowel Syndrome With Diarrhea. Front. Pharmacol. 2020, 11, 629026. [Google Scholar] [CrossRef]
- Ohman, L.; Simrén, M. Pathogenesis of IBS: Role of inflammation, immunity and neuroimmune interactions. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 163–173. [Google Scholar] [CrossRef]
- Abdul Rani, R.; Raja Ali, R.A.; Lee, Y.Y. Irritable bowel syndrome and inflammatory bowel disease overlap syndrome: Pieces of the puzzle are falling into place. Intest. Res. 2016, 14, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, J.R.; Jobin, C. Think small: Zebrafish as a model system of human pathology. J. Biomed. Biotechnol. 2012, 2012, 817341. [Google Scholar] [CrossRef]
- Chassaing, B.; Srinivasan, G.; Delgado, M.A.; Young, A.N.; Gewirtz, A.T.; Vijay-Kumar, M. Fecal lipocalin 2, a sensitive and broadly dynamic non-invasive biomarker for intestinal inflammation. PLoS ONE 2012, 7, e44328. [Google Scholar] [CrossRef] [Green Version]
- Spiller, R.; Garsed, K. Infection, inflammation, and the irritable bowel syndrome. Dig. Liver Dis. 2009, 41, 844–849. [Google Scholar] [CrossRef]
- Buhner, S.; Li, Q.; Vignali, S.; Barbara, G.; De Giorgio, R.; Stanghellini, V.; Cremon, C.; Zeller, F.; Langer, R.; Daniel, H.; et al. Activation of human enteric neurons by supernatants of colonic biopsy specimens from patients with irritable bowel syndrome. Gastroenterology 2009, 137, 1425–1434. [Google Scholar] [CrossRef]
- Clarke, G.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Irritable bowel syndrome: Towards biomarker identification. Trends Mol. Med. 2009, 15, 478–489. [Google Scholar] [CrossRef]
- Dinan, T.G.; Quigley, E.M.; Ahmed, S.M.; Scully, P.; O’Brien, S.; O’Mahony, L.; O’Mahony, S.; Shanahan, F.; Keeling, P.W. Hypothalamic-pituitary-gut axis dysregulation in irritable bowel syndrome: Plasma cytokines as a potential biomarker? Gastroenterology 2006, 130, 304–311. [Google Scholar] [CrossRef]
- Liebregts, T.; Adam, B.; Bredack, C.; Röth, A.; Heinzel, S.; Lester, S.; Downie-Doyle, S.; Smith, E.; Drew, P.; Talley, N.J.; et al. Immune activation in patients with irritable bowel syndrome. Gastroenterology 2007, 132, 913–920. [Google Scholar] [CrossRef]
- Patel, S.R.; Singh, A.; Misra, V.; Misra, S.P.; Dwivedi, M.; Trivedi, P. Levels of interleukins 2, 6, 8, and 10 in patients with irritable bowel syndrome. Indian J. Pathol. Microbiol. 2017, 60, 385–389. [Google Scholar] [CrossRef]
- Dinan, T.G.; Clarke, G.; Quigley, E.M.; Scott, L.V.; Shanahan, F.; Cryan, J.; Cooney, J.; Keeling, P.W. Enhanced cholinergic-mediated increase in the pro-inflammatory cytokine IL-6 in irritable bowel syndrome: Role of muscarinic receptors. Am. J. Gastroenterol. 2008, 103, 2570–2576. [Google Scholar] [CrossRef]
- Xia, Y.; Hu, H.Z.; Liu, S.; Ren, J.; Zafirov, D.H.; Wood, J.D. IL-1beta and IL-6 excite neurons and suppress nicotinic and noradrenergic neurotransmission in guinea pig enteric nervous system. J. Clin. Investig. 1999, 103, 1309–1316. [Google Scholar] [CrossRef]
- O’Malley, D.; Quigley, E.M.; Dinan, T.G.; Cryan, J.F. Do interactions between stress and immune responses lead to symptom exacerbations in irritable bowel syndrome? Brain Behav. Immun. 2011, 25, 1333–1341. [Google Scholar] [CrossRef]
- Natale, L.; Piepoli, A.L.; De Salvia, M.A.; De Salvatore, G.; Mitolo, C.I.; Marzullo, A.; Portincasa, P.; Moschetta, A.; Palasciano, G.; Mitolo-Chieppa, D. Interleukins 1 beta and 6 induce functional alteration of rat colonic motility: An in vitro study. Eur. J. Clin. Investig. 2003, 33, 704–712. [Google Scholar] [CrossRef]
- Kindt, S.; Vanden Berghe, P.; Boesmans, W.; Roosen, L.; Tack, J. Prolonged IL-1beta exposure alters neurotransmitter and electrically induced Ca(2+) responses in the myenteric plexus. Neurogastroenterol. Motil. 2010, 22, 321–e385. [Google Scholar] [CrossRef]
- McKernan, D.P.; Gaszner, G.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Altered peripheral toll-like receptor responses in the irritable bowel syndrome. Aliment. Pharmacol. Ther. 2011, 33, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Buckley, M.M.; O’Halloran, K.D.; Rae, M.G.; Dinan, T.G.; O’Malley, D. Modulation of enteric neurons by interleukin-6 and corticotropin-releasing factor contributes to visceral hypersensitivity and altered colonic motility in a rat model of irritable bowel syndrome. J. Physiol. 2014, 592, 5235–5250. [Google Scholar] [CrossRef] [Green Version]
- Kyoko, O.O.; Kono, H.; Ishimaru, K.; Miyake, K.; Kubota, T.; Ogawa, H.; Okumura, K.; Shibata, S.; Nakao, A. Expressions of tight junction proteins Occludin and Claudin-1 are under the circadian control in the mouse large intestine: Implications in intestinal permeability and susceptibility to colitis. PLoS ONE 2014, 9, e98016. [Google Scholar] [CrossRef]
- Allescher, H.D.; Burgell, R.; Malfertheiner, P.; Mearin, F. Multi-target Treatment for Irritable Bowel Syndrome with STW 5: Pharmacological Modes of Action. J. Gastrointestin. Liver Dis. 2020, 29, 227–233. [Google Scholar] [CrossRef]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 regulates cellular transformation and metastatic behavior in colon cancer. J. Clin. Investig. 2005, 115, 1765–1776. [Google Scholar] [CrossRef] [Green Version]
- Miwa, N.; Furuse, M.; Tsukita, S.; Niikawa, N.; Nakamura, Y.; Furukawa, Y. Involvement of claudin-1 in the beta-catenin/Tcf signaling pathway and its frequent upregulation in human colorectal cancers. Oncol. Res. 2001, 12, 469–476. [Google Scholar] [CrossRef]
- O’Garra, A.; Vieira, P. T(H)1 cells control themselves by producing interleukin-10. Nat. Rev. Immunol. 2007, 7, 425–428. [Google Scholar] [CrossRef]
- Krueger, D.; Gruber, L.; Buhner, S.; Zeller, F.; Langer, R.; Seidl, S.; Michel, K.; Schemann, M. The multi-herbal drug STW 5 (Iberogast) has prosecretory action in the human intestine. Neurogastroenterol. Motil. 2009, 21, 1203-e1110. [Google Scholar] [CrossRef]
- Allam, S.; Krueger, D.; Demir, I.E.; Ceyhan, G.; Zeller, F.; Schemann, M. Extracts from peppermint leaves, lemon balm leaves and in particular angelica roots mimic the pro-secretory action of the herbal preparation STW 5 in the human intestine. Phytomedicine 2015, 22, 1063–1070. [Google Scholar] [CrossRef]
- Aleström, P.; D’Angelo, L.; Midtlyng, P.J.; Schorderet, D.F.; Schulte-Merker, S.; Sohm, F.; Warner, S. Zebrafish: Housing and husbandry recommendations. Lab. Anim. 2020, 54, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Oehlers, S.H.; Flores, M.V.; Hall, C.J.; Okuda, K.S.; Sison, J.O.; Crosier, K.E.; Crosier, P.S. Chemically induced intestinal damage models in zebrafish larvae. Zebrafish 2013, 10, 184–193. [Google Scholar] [CrossRef]
- Chira, A.; Dumitrascu, D.L. Serum biomarkers for irritable bowel syndrome. Clujul Med. 2015, 88, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Bertiaux-Vandaële, N.; Youmba, S.B.; Belmonte, L.; Lecleire, S.; Antonietti, M.; Gourcerol, G.; Leroi, A.M.; Déchelotte, P.; Ménard, J.F.; Ducrotté, P.; et al. The expression and the cellular distribution of the tight junction proteins are altered in irritable bowel syndrome patients with differences according to the disease subtype. Am. J. Gastroenterol. 2011, 106, 2165–2173. [Google Scholar] [CrossRef]
- Kumar, S.; Shukla, R.; Ranjan, P.; Kumar, A. Interleukin-10: A Compelling Therapeutic Target in Patients With Irritable Bowel Syndrome. Clin. Ther. 2017, 39, 632–643. [Google Scholar] [CrossRef]
- Schäfer, K.H.; Saffrey, M.J.; Burnstock, G.; Mestres-Ventura, P. A new method for the isolation of myenteric plexus from the newborn rat gastrointestinal tract. Brain Res. Brain Res. Protoc. 1997, 1, 109–113. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Manufacturing Company |
|---|---|
| Rabbit anti-FZD9 | Immunological Sciences |
| Rabbit anti-WNT3A | Immunological Sciences |
| Mouse anti-PAN Neuronal | Merck Millipore |
| Mouse anti-PAN Cytokeratin | Abcam |
| Mouse anti-Sox2 | Santa Cruz Biotechnology |
| Rabbit anti-Sox10 | Santa Cruz Biotechnology |
| Secondary antibody | |
| Goat anti-rabbit AF488 | Invitrogen |
| Goat anti-mouse AF594 | Invitrogen |
| Goat anti-rabbit PE | Santa Cruz Biotechnology |
| Goat anti-mouse PE | Santa Cruz Biotechnology |
| Gene Name | Primer Sequence (5′-3′) | Accession Number | |
|---|---|---|---|
| eukaryotic translation elongation factor 1 alpha | ef1α | F: TTCGAGAAGGAAGCCGCTG | AY422992 |
| R: CAGCAACAATCAGCACAGCAC | |||
| wnt family member 3A | wnt3A | F: GGCGACTACATGAAGGACAA | AY613787.1 |
| R: TACTTTGGCCGTAGGGTTTC | |||
| claudin 1 | cldn1 | F: CGCCACAGGTGAAGAGTAAA | NM_131770.1 |
| R: CCTCGCGTTAGTTGGAGTAAA | |||
| tumor necrosis factor α | tnfα | F: CCGTCTGCTTCACGCTCC | AY427649.1 |
| R: GTCTTTGATTCAGAGTTGTATCC | |||
| interleukin 8 | il8 | F: TGTTTTCCTGGCATTTCTGACC | XM_009306855 |
| R: TTTACAGTGTGGGCTTGGAGGG | |||
| interleukin 6 | il6 | F: GCTACACTGGCTACACTCTTC | NM_001261449.1 |
| R: GAGACTCTTTACGTCCACATCC | |||
| interleukin 10 | il10 | F: CTCTGCTCACGCTTCTTCTT | NM_001020785.2 |
| R: GCTCCCTCAGTCTTAAAGGAAA | |||
| interleukin 1β | il1β | F: GACATGCTCATGGCGAACG | AY340959.1 |
| R: GCAAATCGTGCATTGCAAGACG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piccione, M.; Facchinello, N.; Schrenk, S.; Gasparella, M.; Pathak, S.; Ammar, R.M.; Rabini, S.; Dalla Valle, L.; Di Liddo, R. STW 5 Herbal Preparation Modulates Wnt3a and Claudin 1 Gene Expression in Zebrafish IBS-like Model. Pharmaceuticals 2021, 14, 1234. https://doi.org/10.3390/ph14121234
Piccione M, Facchinello N, Schrenk S, Gasparella M, Pathak S, Ammar RM, Rabini S, Dalla Valle L, Di Liddo R. STW 5 Herbal Preparation Modulates Wnt3a and Claudin 1 Gene Expression in Zebrafish IBS-like Model. Pharmaceuticals. 2021; 14(12):1234. https://doi.org/10.3390/ph14121234
Chicago/Turabian StylePiccione, Monica, Nicola Facchinello, Sandra Schrenk, Marco Gasparella, Surajit Pathak, Ramy M. Ammar, Sabine Rabini, Luisa Dalla Valle, and Rosa Di Liddo. 2021. "STW 5 Herbal Preparation Modulates Wnt3a and Claudin 1 Gene Expression in Zebrafish IBS-like Model" Pharmaceuticals 14, no. 12: 1234. https://doi.org/10.3390/ph14121234
APA StylePiccione, M., Facchinello, N., Schrenk, S., Gasparella, M., Pathak, S., Ammar, R. M., Rabini, S., Dalla Valle, L., & Di Liddo, R. (2021). STW 5 Herbal Preparation Modulates Wnt3a and Claudin 1 Gene Expression in Zebrafish IBS-like Model. Pharmaceuticals, 14(12), 1234. https://doi.org/10.3390/ph14121234