
Olanzapine Increases Neural Chemorepulsant—Draxin Expression in the Adult Rat Hippocampus

 , , ,
, , ,  , , , and
, , , and 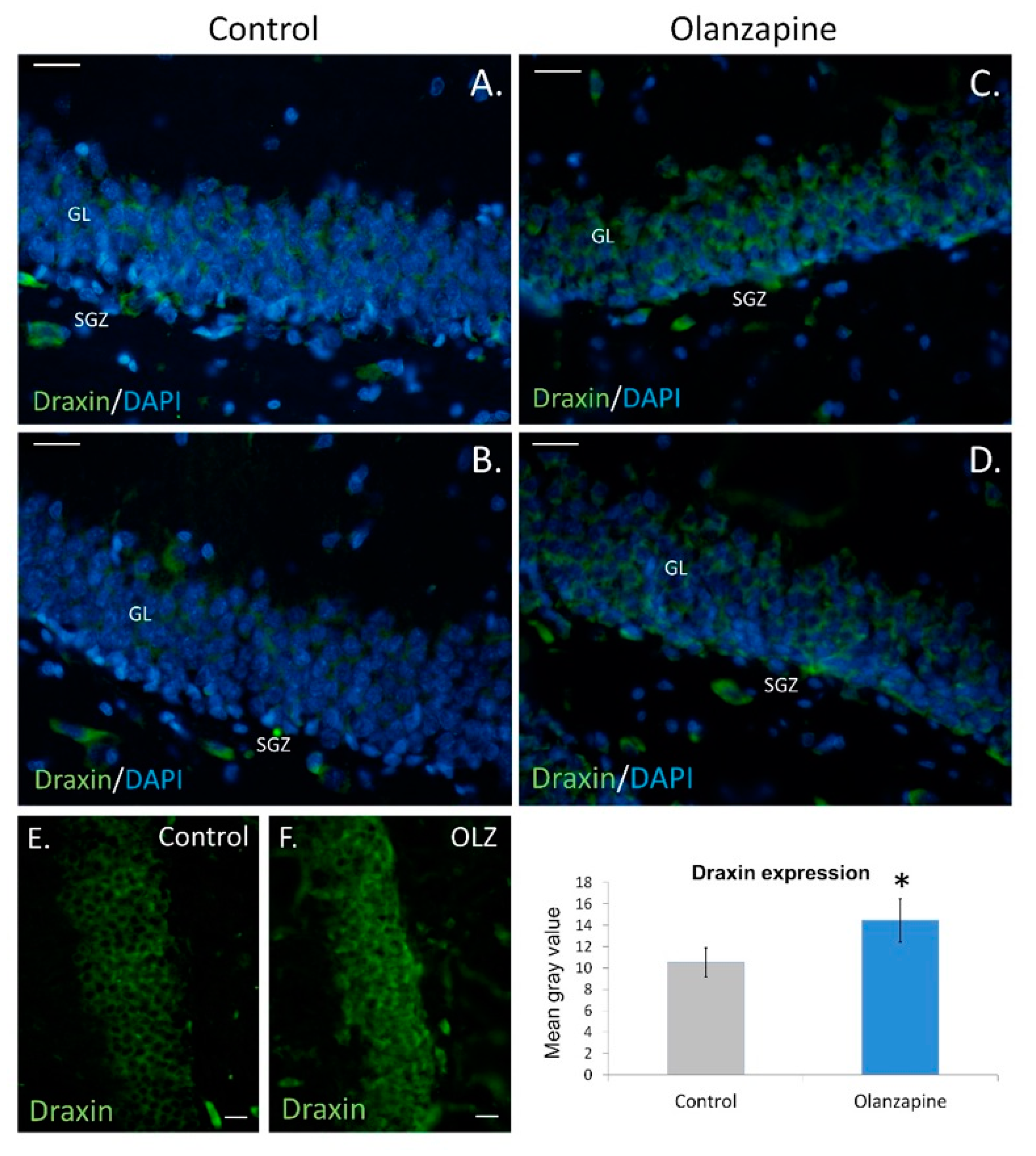{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
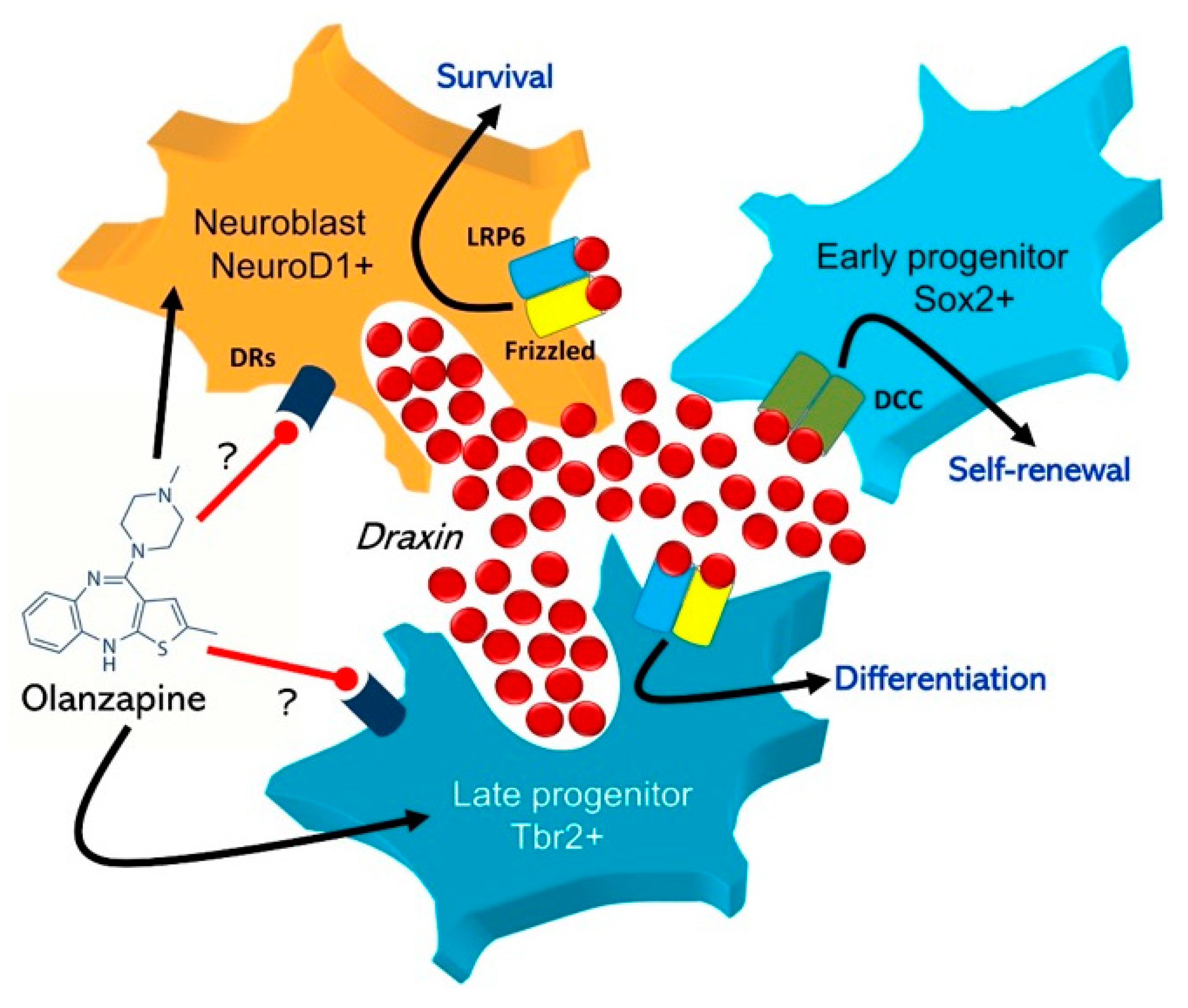
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shinmyo, Y.; Asrafuzzaman Riyadh, M.; Ahmed, G.; Bin Naser, I.; Tanaka, H. Draxin from neocortical neurons controls the guidance of thalamocortical projections into the neocortex. Nat. Commun. 2015, 14, 10232. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Ahmed, G.; Naser, I.B.; Shinmyo, Y.; Ito, A.; Tanaka, H. The combinatorial guidance activities of draxin and Tsukushi are essential for forebrain commissure formation. Dev. Biol. 2013, 374, 58–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naser, I.B.; Su, Y.; Islam, S.M.; Shinmyo, Y.; Zhang, S.; Ahmed, G.; Chen, S.; Tanaka, H. Analysis of a repulsive axon guidance molecule, draxin, on ventrally directed axon projection in chick early embryonic midbrain. Dev. Biol. 2009, 332, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Su, Y.; Shinmyo, Y.; Islam, S.M.; Naser, I.B.; Ahmed, G.; Tamamaki, N.; Tanaka, H. Draxin, a repulsive axon guidance protein, is involved in hippocampal development. Neurosci. Res. 2010, 66, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, S.; Islam, S.M.; Shinmyo, Y.; Naser, I.B.; Tanaka, H. Draxin is involved in the proper development of the dI3 interneuron in chick spinal cord. Dev. Dyn. 2010, 239, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.M.; Shinmyo, Y.; Okafuji, T.; Su, Y.; Naser, I.B.; Tanaka, H. Draxin, a repulsive guidance protein for spinal cord and forebrain commissures. Science 2009, 323, 388–393. [Google Scholar] [CrossRef]
- Liu, Y.; Bhowmick, T.; Liu, Y.; Gao, X.; Mertens, H.D.T.; Meijers, R. Structural Basis for Draxin-Modulated Axon Guidance and Fasciculation by Netrin-1 through DCC. Neuron 2018, 97, 261–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawarayama, H.; Yamada, H.; Amin, R.; Morita-Fujimura, Y.; Cooper, H.M.; Ikawa, S. Draxin-mediated Regulation of Granule Cell Progenitor Differentiation in the Postnatal Hippocampal Dentate Gyrus. Neuroscience 2020, 431, 184–192. [Google Scholar] [CrossRef]
- Tawarayama, H.; Yamada, H.; Amin, R.; Morita-Fujimura, Y.; Cooper, H.M.; Tanaka, H. Draxin regulates hippocampal neurogenesis in the postnatal dentate gyrus by inhibiting DCC-induced apoptosis. Sci. Rep. 2018, 16, 840. [Google Scholar] [CrossRef]
- Miyake, A.; Nihno, S.; Murakoshi, Y.; Satsuka, A.; Nakayama, Y.; Itoh, N. Neucrin, a novel secreted antagonist of canonical Wnt signaling, plays roles in developing neural tissues in zebrafish. Mech. Dev. 2012, 128, 577–590. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.T.; Nasrallah, H.A. Neuroprotective effects of the second generation antipsychotics. Schizophr Res. 2019, 208, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Krzystanek, M.; Krzystanek, E.; Skałacka, K.; Pałasz, A. Enhancement in Phospholipase D Activity as a New Proposed Molecular Mechanism of Haloperidol-Induced Neurotoxicity. Int. J. Mol. Sci. 2020, 4, 23–9265. [Google Scholar]
- Huhn, M.; Nikolakopoulou, A.; Schneider-Thoma, J.; Krause, M.; Samara, M.; Peter, N.; Arndt, T.; Bäckers, L.; Rothe, P.; Cipriani, A.; et al. Comparative efficacy and tolerability of 32 oral antipsychotics for the acute treatment of adults with multi-episode schizophrenia: A systematic review and network meta-analysis. Lancet 2019, 14, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Novick, D.; Montgomery, W.; Treuer, T.; Moneta, M.V.; Haro, J.M. Real-world Effectiveness of Antipsychotics for the Treatment of Negative Symptoms in Patients with Schizophrenia with Predominantly Negative Symptoms. Pharmacopsychiatry 2017, 50, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Krzystanek, M.; Pałasz, A. NMDA Receptor Model of Antipsychotic Drug-Induced Hypofrontality. Int. J. Mol. Sci. 2019, 21, 1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishara, D.; Olofinjana, O.; Sparshatt, A.; Kapur, S.; Taylor, D.; Patel, M.X. Olanzapine: A systematic review and meta-regression of the relationships between dose, plasma concentration, receptor occupancy, and response. J. Clin. Psychopharmacol. 2013, 33, 329–335. [Google Scholar] [CrossRef]
- Olijslagers, J.E.; Werkman, T.R.; McCreary, A.C.; Kruse, C.G.; Wadman, W.J. Modulation of midbrain dopamine neurotransmission by serotonin, a versatile interaction between neurotransmitters and significance for antipsychotic drug action. Curr. Neuropharmacol. 2006, 4, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Duggan, L.; Fenton, M.; Rathbone, J.; Dardennes, R.; El-Dosoky, A.; Indran, S. Olanzapine for schizophrenia. Cochrane Database Syst. Rev. 2005, 18, CD001359. [Google Scholar] [CrossRef]
- Wakade, C.G.; Mahadik, S.P.; Waller, J.L.; Chiu, F.C. Atypical neuroleptics stimulate neurogenesis in adult rat brain. J. Neurosci. Res. 2002, 1, 72–79. [Google Scholar] [CrossRef]
- Bai, O.; Chlan-Fourney, J.; Bowen, R.; Keegan, D.; Li, X.M. Expression of brain-derived neurotrophic factor mRNA in rat hippocampus after treatment with antipsychotic drugs. J. Neurosci. Res. 2003, 71, 1127–1131. [Google Scholar] [CrossRef]
- Lieberman, J.A.; Tollefson, G.D.; Charles, C.; Zipursky, R.; Sharma, T.; Kahn, R.S.; Keefe, R.S.; Green, A.I.; Gur, R.E.; McEvoy, J.; et al. HGDH Study Group. Antipsychotic drug effects on brain morphology in first-episode psychosis. Arch. Gen. Psychiatry 2005, 62, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Vita, A.; De Peri, L.; Deste, G.; Barlati, S.; Sacchetti, E. The Effect of Antipsychotic Treatment on Cortical Gray Matter Changes in Schizophrenia: Does the Class Matter? A Meta-analysis and Meta-regression of Longitudinal Magnetic Resonance Imaging Studies. Biol. Psychiatry. 2015, 78, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.T.; Zhang, Y.; Liu, R.; Wang, X.D.; Su, Y.A. Prenatal Exposure to Antipsychotics Disrupts the Plasticity of Dentate Neurons and Memory in Adult Male Mice. Int. J. Neuropsychopharmacol. 2019, 22, 71–82. [Google Scholar] [CrossRef]
- Zheng, P.; Hu, M.; Xie, Y.; Yu, Y.; Jaaro-Peled, H.; Huang, X.F. Aripiprazole and haloperidol protect neurite lesions via reducing excessive D2R-DISC1 complex formation. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cui, H.; Wang, L.; Kang, L.; Huang, G.; Su, Y. Potential Involvement of Draxin in the Axonal Projection of Cranial Nerves, Especially Cranial Nerve X, in the Chick Hindbrain. J. Histochem. Cytochem. 2016, 64, 412–424. [Google Scholar] [CrossRef] [Green Version]
- Łasut, B.; Pałasz, A.; Filipczyk, L.; Arias-Carrion, O.; Rojczyk, E.; Wiaderkiewicz, R. Long-term Treatment with Olanzapine Increases the Number of Sox2 and Doublecortin Expressing Cells in the Adult Subventricular Zone. CNS Neurol. Disord. Drug Targets 2018, 17, 458–463. [Google Scholar] [CrossRef]
- Song, J.C.; Seo, M.K.; Park, S.W.; Lee, J.G.; Kim, Y.H. Differential Effects of Olanzapine and Haloperidol on MK-801-induced Memory Impairment in Mice. Clin. Psychopharmacol. Neurosci. 2016, 31, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Rojczyk, E.; Pałasz, A.; Wiaderkiewicz, R. Effects of neuroleptics administration on adult neurogenesis in the rat hypothalamus. Pharmacol. Rep. 2015, 67, 1208–1214. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, Y.; Huang, X.F. Olanzapine Prevents the PCP-induced Reduction in the Neurite Outgrowth of Prefrontal Cortical Neurons via NRG1. Sci. Rep. 2016, 6, 19581. [Google Scholar] [CrossRef]
- Matsushima, Y.; Terada, K.; Takata, J.; Karube, Y.; Kamei, C.; Sugimoto, Y. Effects of fluvoxamine on nerve growth factor-induced neurite outgrowth inhibition by dexamethasone in PC12 cells. Biosci. Biotechnol. Biochem. 2019, 83, 659–665. [Google Scholar] [CrossRef]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamura, N.; Nakagawa, S.; Masuda, T.; Boku, S.; Kato, A.; Kusumi, I. The effect of dopamine on adult hippocampal neurogenesis. Prog Neuropsychopharmacol. Biol. Psychiatry. 2014, 50, 116–124. [Google Scholar]
- Höglinger, G.U.; Rizk, P.; Muriel, M.P.; Duyckaerts, C.; Oertel, W.H.; Hirsch, E.C. Dopamine depletion impairs precursor cell proliferation in Parkinson disease. Nat. Neurosci. 2004, 7, 726–735. [Google Scholar] [CrossRef]
- Singh, K.P.; Singh, M.K.; Singh, M. Effects of prenatal exposure to antipsychotic risperidone on developmental neurotoxicity, apoptotic neurodegeneration and neurobehavioral sequelae in rat offspring. Int. J. Dev. Neurosci. 2016, 52, 13–23. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pałasz, A.; Suszka-Świtek, A.; Francikowski, J.; Krzystanek, M.; Bogus, K.; Skałbania, J.; Worthington, J.J.; Mrzyk, I. Olanzapine Increases Neural Chemorepulsant—Draxin Expression in the Adult Rat Hippocampus. Pharmaceuticals 2021, 14, 298. https://doi.org/10.3390/ph14040298
Pałasz A, Suszka-Świtek A, Francikowski J, Krzystanek M, Bogus K, Skałbania J, Worthington JJ, Mrzyk I. Olanzapine Increases Neural Chemorepulsant—Draxin Expression in the Adult Rat Hippocampus. Pharmaceuticals. 2021; 14(4):298. https://doi.org/10.3390/ph14040298
Chicago/Turabian StylePałasz, Artur, Aleksandra Suszka-Świtek, Jacek Francikowski, Marek Krzystanek, Katarzyna Bogus, Jakub Skałbania, John J. Worthington, and Inga Mrzyk. 2021. "Olanzapine Increases Neural Chemorepulsant—Draxin Expression in the Adult Rat Hippocampus" Pharmaceuticals 14, no. 4: 298. https://doi.org/10.3390/ph14040298
APA StylePałasz, A., Suszka-Świtek, A., Francikowski, J., Krzystanek, M., Bogus, K., Skałbania, J., Worthington, J. J., & Mrzyk, I. (2021). Olanzapine Increases Neural Chemorepulsant—Draxin Expression in the Adult Rat Hippocampus. Pharmaceuticals, 14(4), 298. https://doi.org/10.3390/ph14040298