
Wound Healing Promotion by Hyaluronic Acid: Effect of Molecular Weight on Gene Expression and In Vivo Wound Closure

 ,
,  ,
, 
 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Physicochemical Analysis of HA Ointments
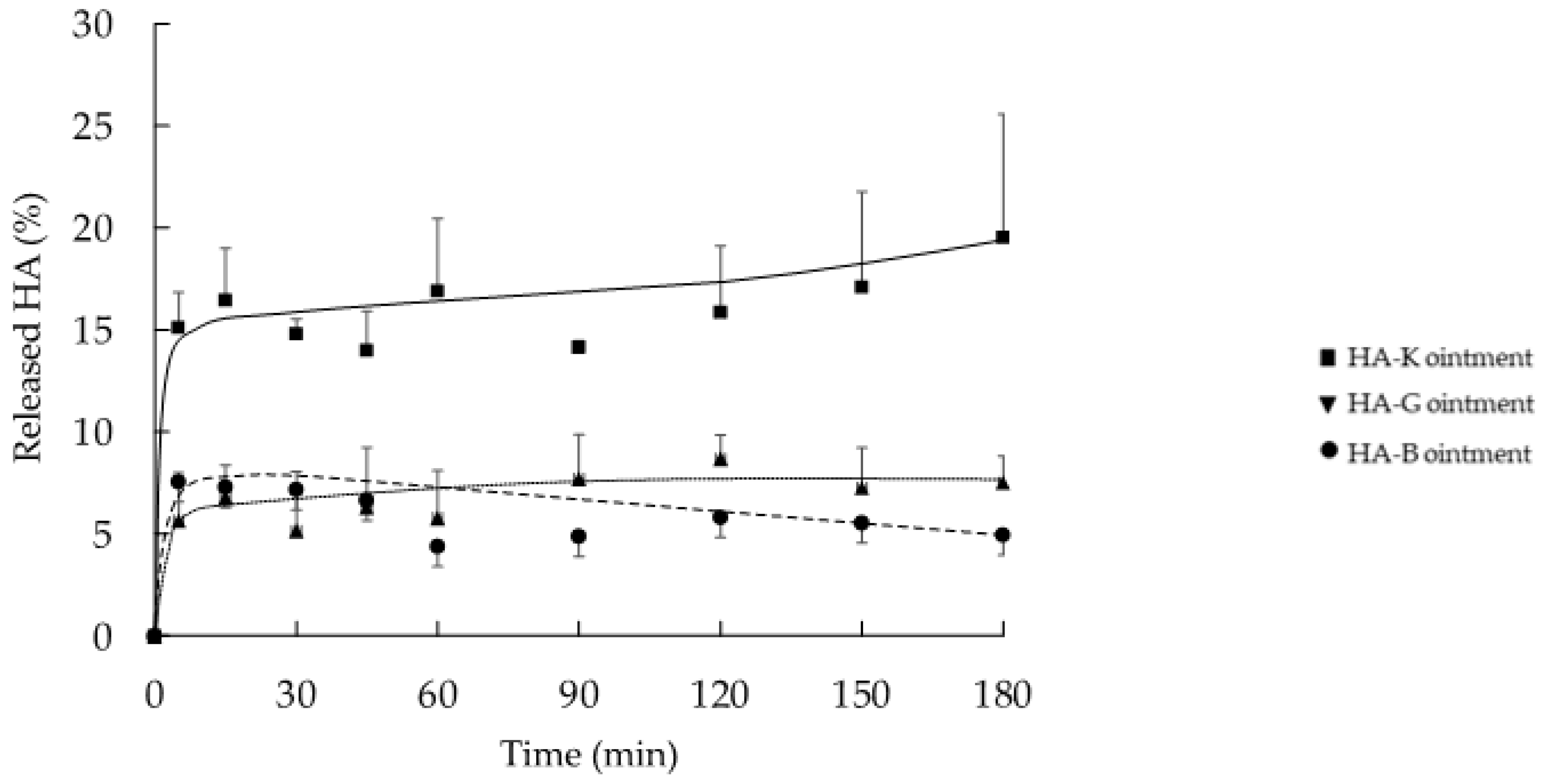
2.2. In Vitro Release Profile of HA from the Ointment Formulation
2.3. In Vitro Proliferation of HaCaT and NHDF Cell
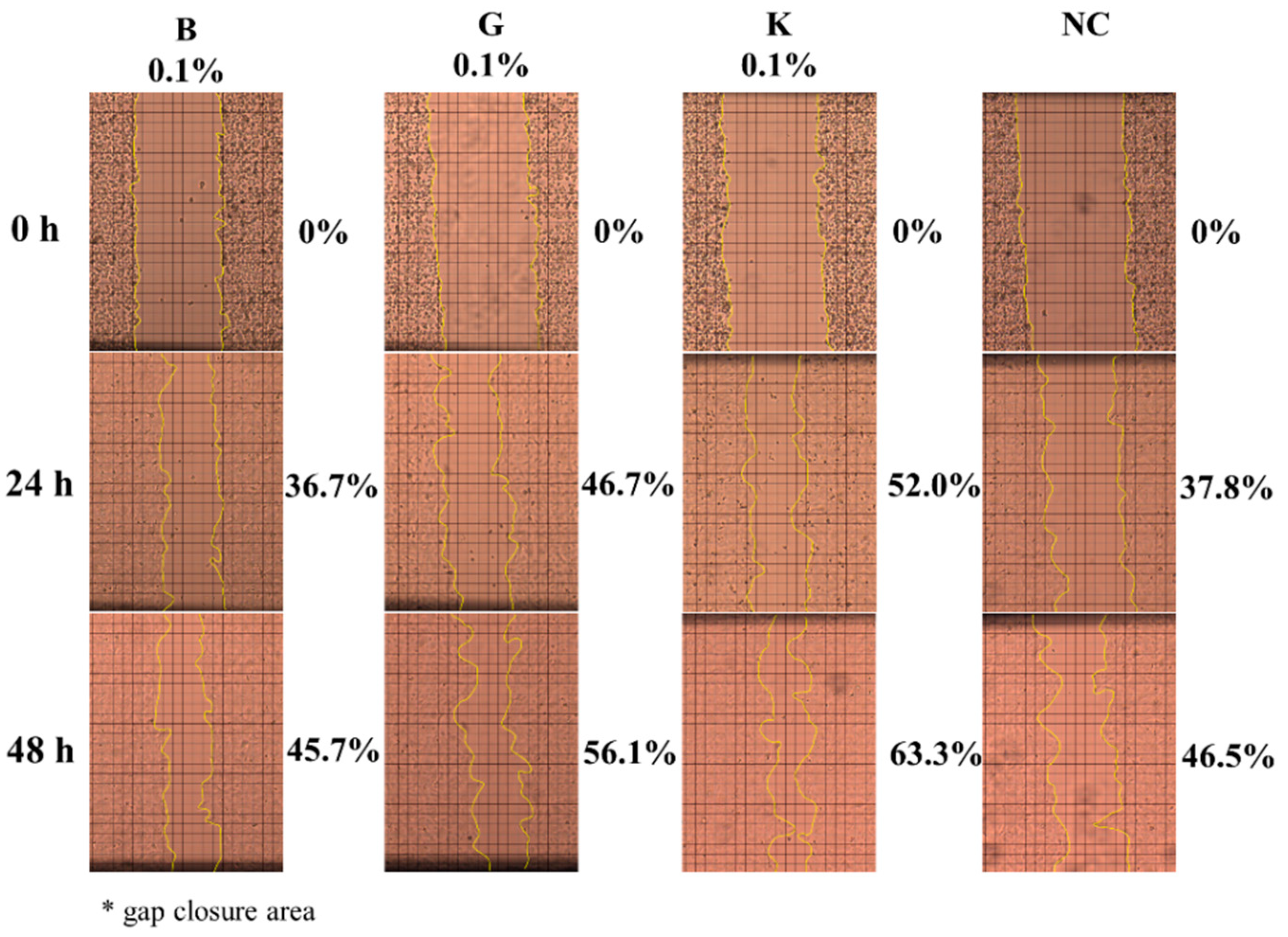
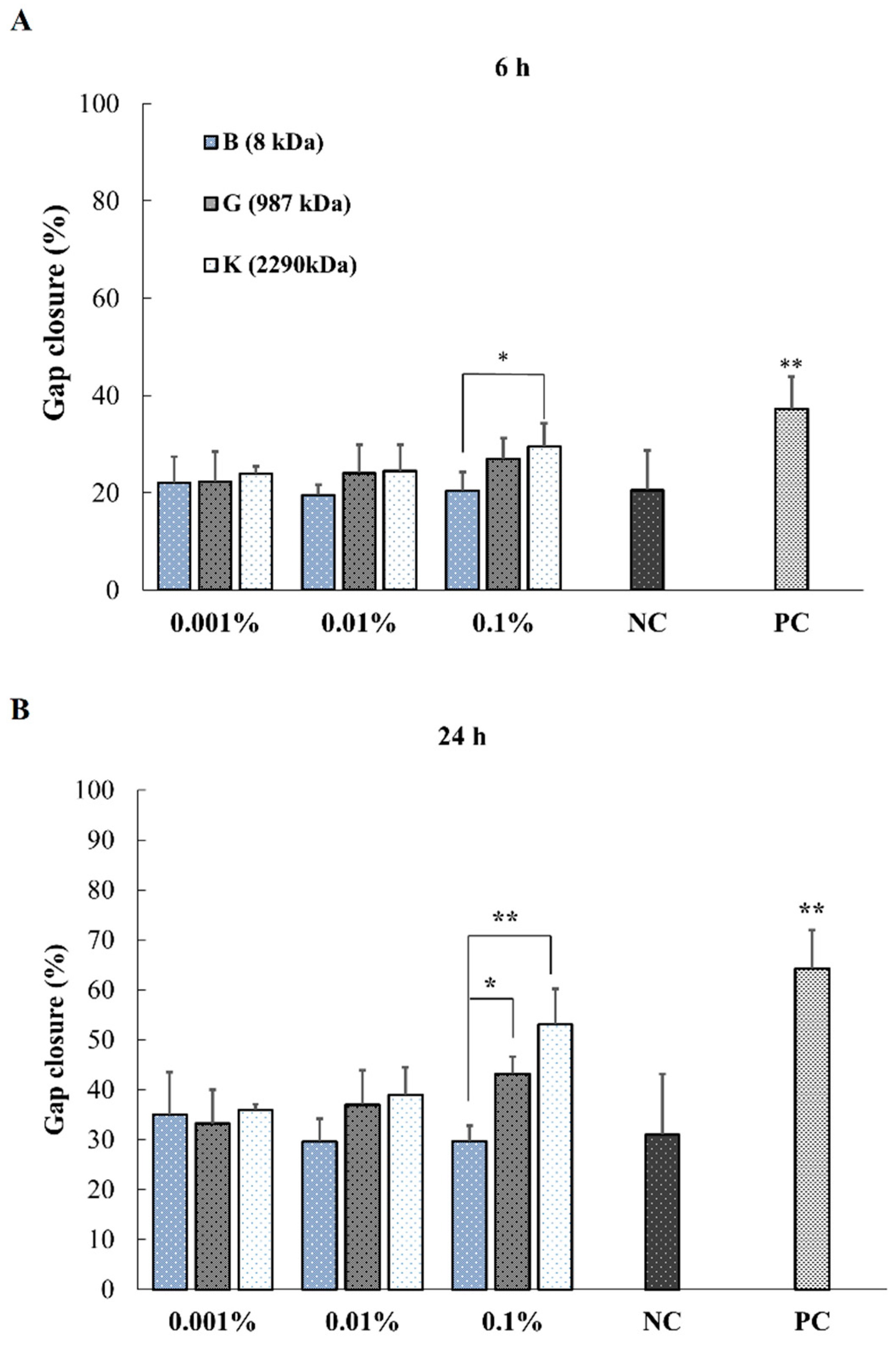
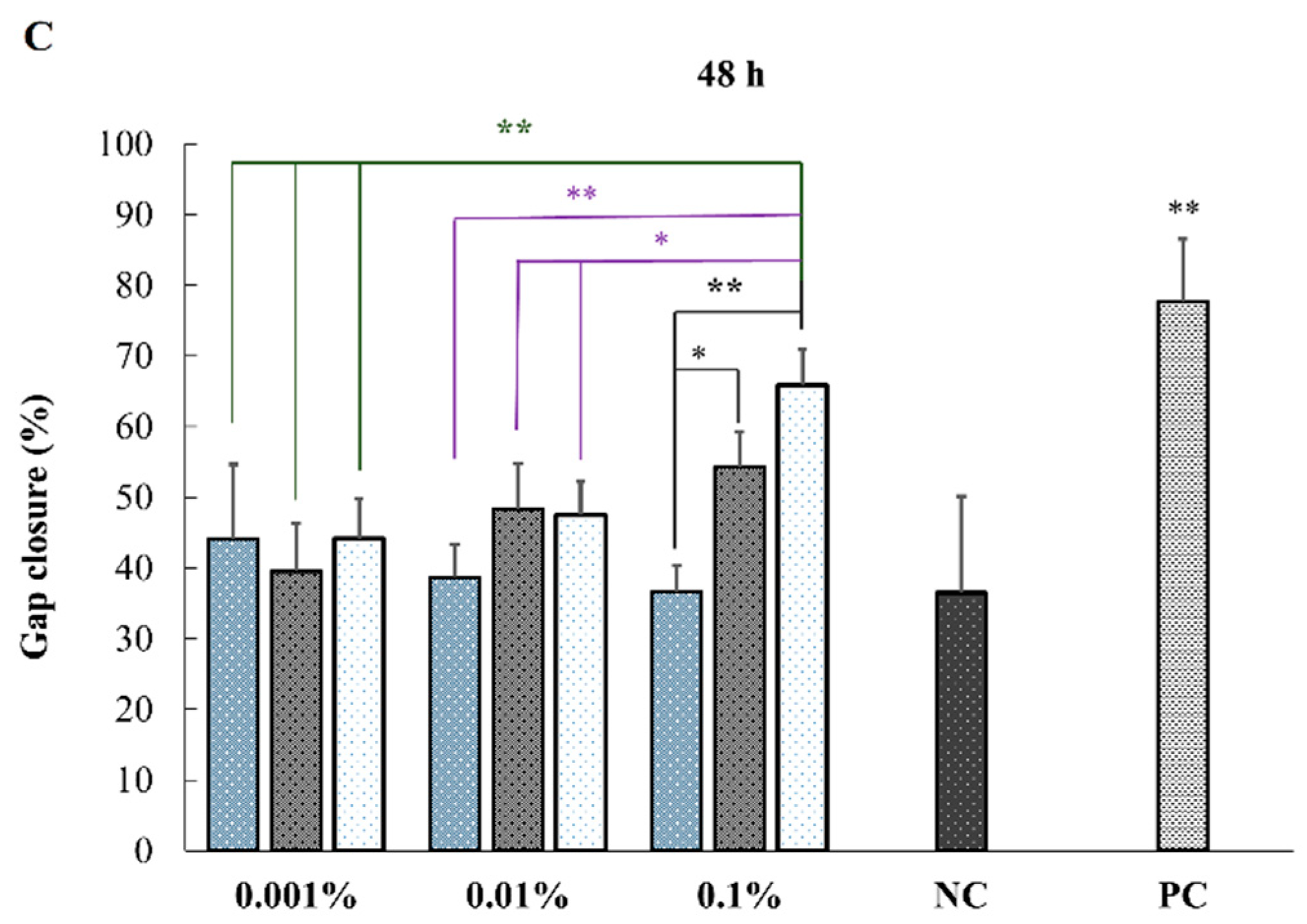
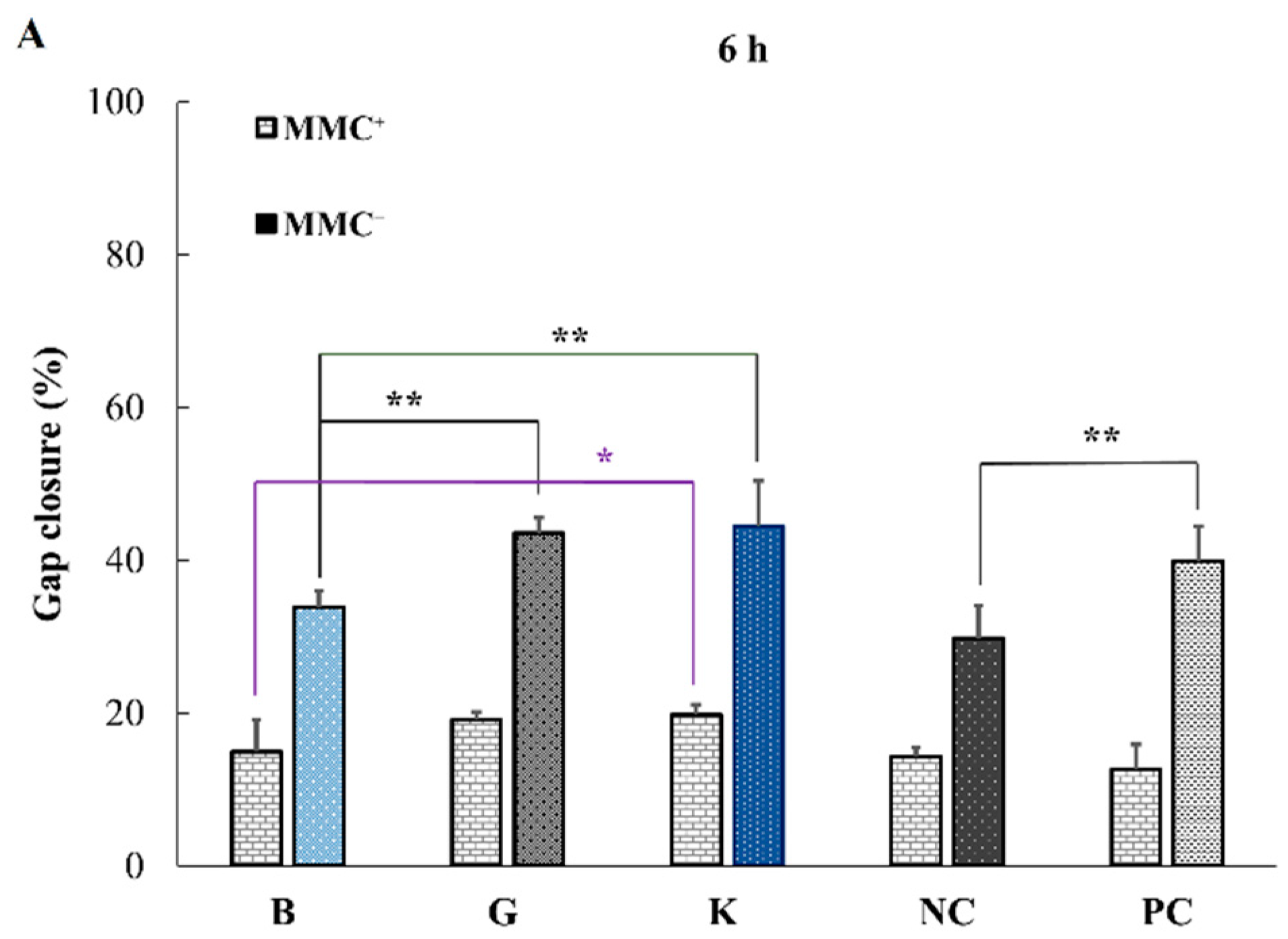
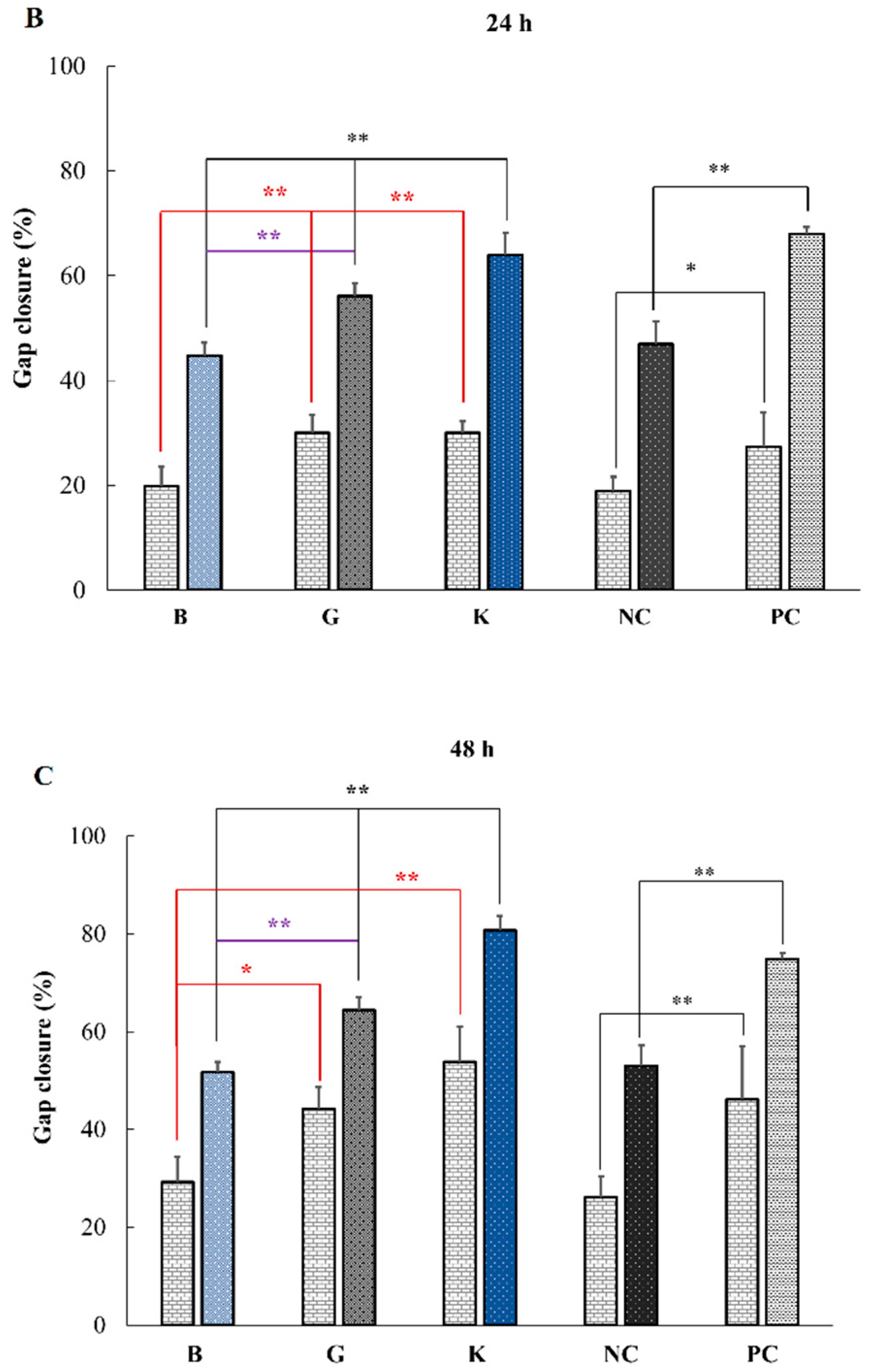
2.4. In Vitro Effect of HA on Keratinocytes or Fibroblasts Wound Closure
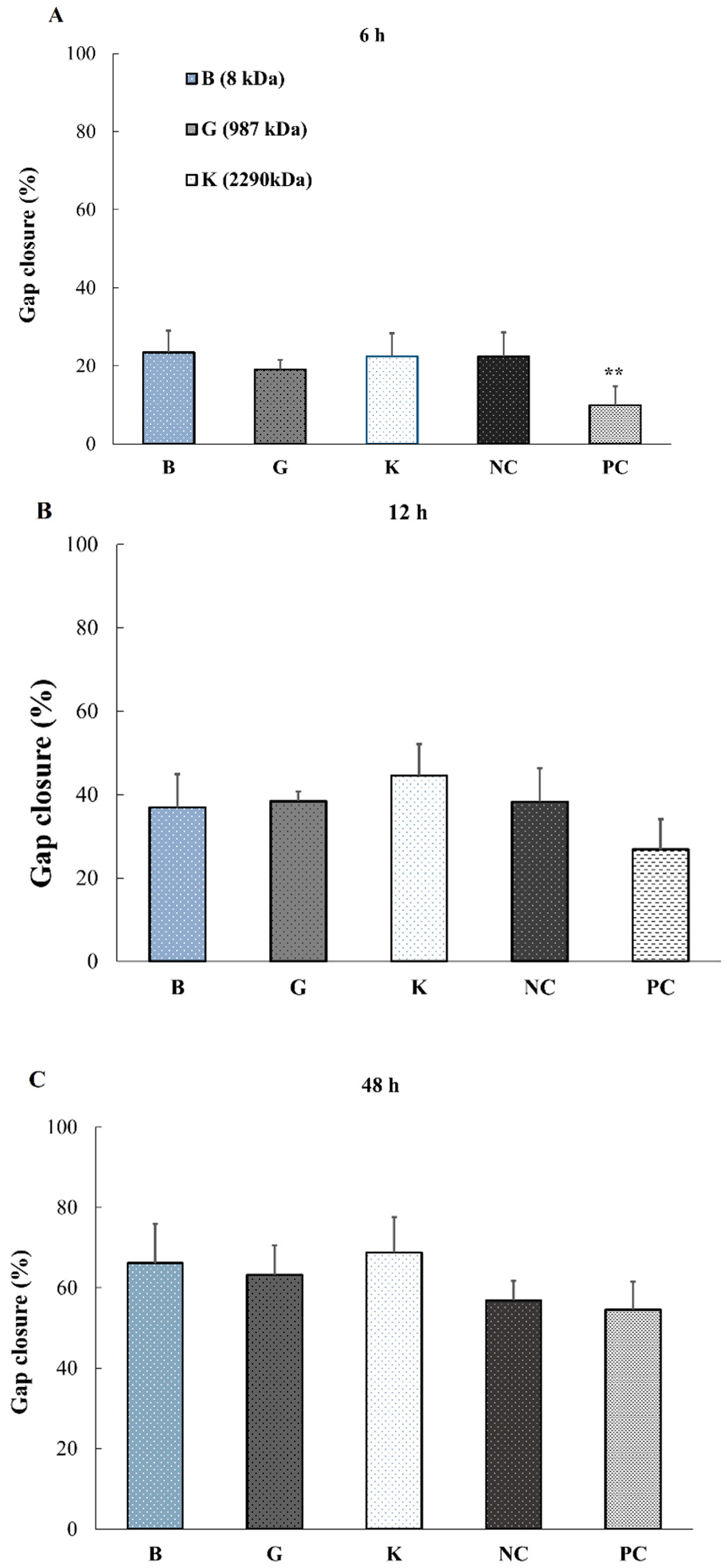
2.5. Effect of HA Addition on the Promotion of Wound Healing
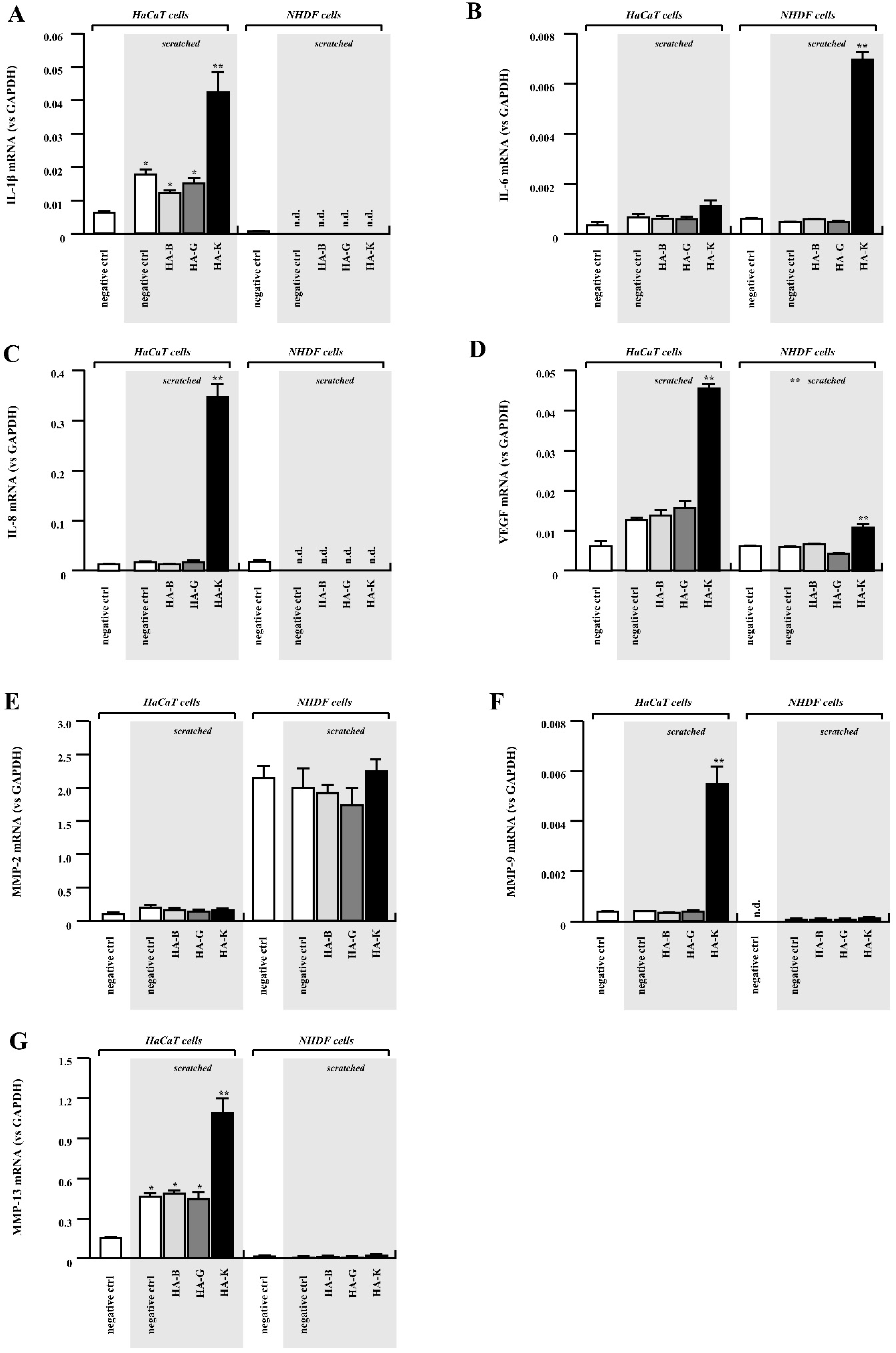
2.6. Gene Expression Analysis on Keratinocytes and Fibroblasts Exposed to HA
3. Discussion
4. Materials and Methods
4.1. Physicochemical Analysis: Assessment of Adhesiveness and Spreadability and Content Uniformity of HA Ointments
4.2. In Vitro Release Profile of HA from Ointment Formulations
4.3. Endotoxin Assay
4.4. Effects of HA Addition on Cell Proliferation In Vitro
4.4.1. In Vitro Cell Proliferation Assay (WST-8 Assay)
4.4.2. In Vitro Wound Scratch Assay
4.4.3. Realtime-PCR Analysis
4.5. In Vivo Wound Healing Experiments in Mice
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patrulea, V.; Ostafe, V.; Borchard, G.; Jordan, O. Chitosan as a starting material for wound healing applications. Eur. J. Pharm. Biopharm. 2015, 97, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrulea, V.; Younes, I.; Jordan, O.; Borchard, G. Chitosan-Based Systems for Controlled Delivery of Antimicrobial Peptides for Biomedical Application. In Functional Chitosan: Drug Delivery and Biomedical Applications; Sougata, J., Subrata, J., Eds.; Springer: Singapore, Singapore, 2019; pp. 415–455. [Google Scholar]
- Ribeiro, D.M.L.; Carvalho Júnior, A.R.; Vale De Macedo, G.H.R.; Chagas, V.L.; Silva, L.D.S.; Cutrim, B.D.S.; Santos, D.M.; Soares, B.L.L.; Zagmignan, A.; De Miranda, R.C.M.; et al. Polysaccharide-Based Formulations for Healing of Skin-Related Wound Infections: Lessons from Animal Models and Clinical Trials. Biomolecules 2019, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghatak, S.; Maytin, E.V.; Mack, J.A.; Hascall, V.C.; Atanelishvili, I.; Moreno Rodriguez, R.; Markwald, R.R.; Misra, S. Roles of Proteoglycans and Glycosaminoglycans in Wound Healing and Fibrosis. Int. J. Cell Biol. 2015, 2015, 834893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular Mediators of Angiogenesis. J. Burn. Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schanté, C.E.; Zuber, G.; Herlin, C.; Vandamme, T.F. Chemical modifications of hyaluronic acid for the synthesis of derivatives for a broad range of biomedical applications. Carbohydr. Polym. 2011, 85, 469–489. [Google Scholar] [CrossRef]
- Tavianatou, A.G.; Caon, I.; Franchi, M.; Piperigkou, Z.; Galesso, D.; Karamanos, N.K. Hyaluronan: Molecular size-dependent signaling and biological functions in inflammation and cancer. FEBS J. 2019, 286, 2883–2908. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.N.; Birkinshaw, C. Hyaluronic acid based scaffolds for tissue engineering—A review. Carbohydr. Polym. 2013, 92, 1262–1279. [Google Scholar] [CrossRef]
- Patrulea, V.; Laurent-Applegate, L.A.; Ostafe, V.; Borchard, G.; Jordan, O. Polyelectrolyte nanocomplexes based on chitosan derivatives for wound healing application. Eur. J. Pharm. Biopharm. 2019, 140, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xue, Y.; Jia, B.; Bai, Y.; Zuo, Y.; Wang, S.; Zhao, Y.; Yang, W.; Tang, H. The preparation of hyaluronic acid grafted pullulan polymers and their use in the formation of novel biocompatible wound healing film. Carbohydr. Polym. 2018, 188, 92–100. [Google Scholar] [CrossRef]
- Nyman, E.; Henricson, J.; Ghafouri, B.; Anderson, C.D.; Kratz, G. Hyaluronic Acid Accelerates Re-epithelialization and Alters Protein Expression in a Human Wound Model. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2221. [Google Scholar] [CrossRef] [PubMed]
- Iocono, J.A.; Ehrlich, H.P.; Keefer, K.A.; Krummel, T.M. Hyaluronan induces scarless repair in mouse limb organ culture. J. Pediatr. Surg. 1998, 33, 564–567. [Google Scholar] [CrossRef]
- Fallacara, A.; Baldini, E.; Manfredini, S.; Vertuani, S. Hyaluronic Acid in the Third Millennium. Polymers 2018, 10, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Yang, C.X.; Mo, W.; Liu, Y.W.; He, Y.Q. Hyaluronan oligosaccharides are potential stimulators to angiogenesis via RHAMM mediated signal pathway in wound healing. Clin. Investig. Med. 2008, 31, E106–E116. [Google Scholar] [CrossRef] [Green Version]
- Saettone, M.F.; Chetoni, P.; Tilde Torracca, M.; Burgalassi, S.; Giannaccini, B. Evaluation of muco-adhesive properties and in vivo activity of ophthalmic vehicles based on hyaluronic acid. Int. J. Pharm. 1989, 51, 203–212. [Google Scholar] [CrossRef]
- Snetkov, P.; Zakharova, K.; Morozkina, S.; Olekhnovich, R.; Uspenskaya, M. Hyaluronic Acid: The Influence of Molecular Weight on Structural, Physical, Physico-Chemical, and Degradable Properties of Biopolymer. Polymers 2020, 12, 1800. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, S.; Sun, Y.; Wang, J. Mitomycin C induces fibroblast apoptosis and reduces intra-articular scar adhesion by regulating miR-21 and its target Programmed cell death 4. Fitoterapia 2020, 142, 104392. [Google Scholar] [CrossRef] [PubMed]
- Zgheib, C.; Xu, J.; Liechty, K.W. Targeting Inflammatory Cytokines and Extracellular Matrix Composition to Promote Wound Regeneration. Adv. Wound Care 2014, 3, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnard, N.; Legeais, J.M.; Renard, G.; Robert, L. Effect of hyaluronan on MMP expression and activation. Cell Biol. Int. 2001, 25, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Bronzatto, E.; Riccetto, C.L.Z. Pro-inflammatory cytokines and metalloproteinase activation in polypropylene mesh implant in rat subcutaneous tissue. Int. Braz. J. Urol. 2018, 44, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Li, N.; Teng, W.; Wang, M.; Zhang, Y.; Xiao, Z. TGF-β1 promotes scar fibroblasts proliferation and transdifferentiation via up-regulating MicroRNA-21. Sci. Rep. 2016, 6, 32231. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Li, L.; Jiang, C.; Xu, W.; Lv, Y.; Zhong, L.; Cai, K.; Yang, L. The effect of hyaluronan on the motility of skin dermal fibroblasts in nanofibrous scaffolds. Int. J. Biol. Macromol. 2015, 79, 133–143. [Google Scholar] [CrossRef]
- Fingleton, B. Matrix metalloproteinases as regulators of inflammatory processes. BBA Mol. Cell Res. 2017, 1864, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, R.C.; Siebert, J.W.; Eidelman, Y.; Gold, L.I.; Longaker, M.T.; Garg, H.G. The in vivo effect of hyaluronan associated protein-collagen complex on wound repair. Biochem. Mol. Biol. Int. 1995, 37, 151–158. [Google Scholar]
- Takabe, P.; Kärnä, R.; Rauhala, L.; Tammi, M.; Tammi, R.; Pasonen-Seppänen, S. Melanocyte Hyaluronan Coat Fragmentation Enhances the UVB-Induced TLR-4 Receptor Signaling and Expression of Proinflammatory Mediators IL6, IL8, CXCL1, and CXCL10 via NF-κB Activation. J. Investig. Dermatol. 2019, 139, 1993–2003.e4. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.B.; Barroso, W.A.; Da Silva, N.N.; Silva, S.D.N.; Borges, A.C.R.; Abreu, I.C.; Borges, M. From Inflammation to Current and Alternative Therapies Involved in Wound Healing. Int. J. Inflam. 2017, 2017, 3406215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourguignon, L.Y.W.; Ramez, M.; Gilad, E.; Singleton, P.A.; Man, M.-Q.; Crumrine, D.A.; Elias, P.M.; Feingold, K.R. Hyaluronan–CD44 Interaction Stimulates Keratinocyte Differentiation, Lamellar Body Formation/Secretion, and Permeability Barrier Homeostasis. J. Investig. Dermatol. 2006, 126, 1356–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Cao, M.; Liu, H.; He, Y.; Xu, J.; Du, Y.; Liu, Y.; Wang, W.; Cui, L.; Hu, J.; et al. The high and low molecular weight forms of hyaluronan have distinct effects on CD44 clustering. J. Biol. Chem. 2012, 287, 43094–43107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Mack, J.A.; Maytin, E.V. CD44 inhibits α-SMA gene expression via a novel G-actin/MRTF-mediated pathway that intersects with TGFβR/p38MAPK signaling in murine skin fibroblasts. J. Biol. Chem. 2019, 294, 12779–12794. [Google Scholar] [CrossRef]
- Price, Z.K.; Lokman, N.A.; Ricciardelli, C. Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance. Cancers 2018, 10, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainio, A.; Järveläinen, H. Extracellular matrix-cell interactions: Focus on therapeutic applications. Cell. Signal. 2020, 66, 109487. [Google Scholar] [CrossRef] [PubMed]
- Pardue, E.L.; Ibrahim, S.; Ramamurthi, A. Role of hyaluronan in angiogenesis and its utility to angiogenic tissue engineering. Organogenesis 2008, 4, 203–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundschell, C.S.; Jakob, F.; Wagemans, A.M. Molecular weight dependent structure of the exopolysaccharide levan. Int. J. Biol. Macromol. 2020, 161, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Kawata, K.; Hanawa, T.; Endo, N.; Suzuki, M.; Oguchi, T. Formulation study on retinoic acid gel composed of iota-carrageenan, polyethylene oxide and Emulgen® 408. Chem. Pharm. Bull. 2012, 60, 825–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Spreadability (mm2) | Yield Value (dyne/cm2) | Adhesion Energy (J/m3) |
|---|---|---|---|
| PBS | 760.9 ± 0.06 | 925.0 ± 21.33 | 1960 ± 99.25 |
| HA-B | 780.3 ± 0.40 | 921.8 ± 52.02 | 1920 ± 271.8 |
| HA-G | 752.7 ± 0.56 | 881.4 ± 27.46 | 2030 ± 65.07 |
| HA-K | 791.4 ± 0.01 | 850.2 ± 7.35 | 2120 ± 58.31 |
| MMC+ Results | 6 h | 24 h | 48 h |
|---|---|---|---|
| B vs. G | ns | ** | * |
| B vs. K | * | ** | ** |
| G vs. K | ns | ns | ns |
| Genes Influencing Wound Healing | HA | ||
|---|---|---|---|
| B | G | K | |
| TGF-β1 | ↑↑ | ↑ | ↑ |
| TGF-β3 | = | = | ↓ |
| VEGF | = | ↑ | ↑ |
| IL-1β | ↓ | = | ↑ |
| IL-6 | = | = | ↑ |
| IL-8 | ↓ | = | ↑↑↑ |
| MMP-2 | = | = | = |
| MMP-9 | = | = | ↑↑↑ |
| MMP-13 | = | = | ↑ |
| Genes | Forward Primer | Reverse Primer |
|---|---|---|
| [Target] | ||
| TGF-β1 | GCCCTGGACACCAACTATTGC | GCACTTGCAGGAGCGCA |
| TGF-β3 | AAGYGGGYCCATGAACCTAA | GCTACATTTACAAGACTTCAC |
| IL-1β | AAAAGCTTGGTGATGTCTGG | TTTCAACACGCAGGACAGG |
| IL-6 | GACTGGAGATGTCTGAGGCTCAT | CCCAGGGAGAAGGCAACTG |
| IL-8 | ATGACTTCCAAGCTGGGCCGTG | TATGAATTCTCAGCCCTCTTCAAAA |
| VEGF | GAGGCCTTGCCTTGCTGCTCTA | CACCAGGGTCTCGATTGGAT |
| MMP-2 | AGATCTTCTTCTTCAAGGACCGGTT | GGCTGGTCAGTGGCTTGGGGTA |
| MMP-9 | ATTTCTGCCAGGACCGCTTCTACT | CAGTTTGTATCCGGCAAACTGGCT |
| MMP-13 | TCCCAGGAATTGGTGATAAAGTAGA | CTGGCATGACGCGAACAATA |
| [internal control] | ||
| GAPDH | CCCATGTTCGTCATGGGTGT | TGGTCATGAGTCCTTCCACGATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawano, Y.; Patrulea, V.; Sublet, E.; Borchard, G.; Iyoda, T.; Kageyama, R.; Morita, A.; Seino, S.; Yoshida, H.; Jordan, O.; et al. Wound Healing Promotion by Hyaluronic Acid: Effect of Molecular Weight on Gene Expression and In Vivo Wound Closure. Pharmaceuticals 2021, 14, 301. https://doi.org/10.3390/ph14040301
Kawano Y, Patrulea V, Sublet E, Borchard G, Iyoda T, Kageyama R, Morita A, Seino S, Yoshida H, Jordan O, et al. Wound Healing Promotion by Hyaluronic Acid: Effect of Molecular Weight on Gene Expression and In Vivo Wound Closure. Pharmaceuticals. 2021; 14(4):301. https://doi.org/10.3390/ph14040301
Chicago/Turabian StyleKawano, Yayoi, Viorica Patrulea, Emmanuelle Sublet, Gerrit Borchard, Takuya Iyoda, Rihoko Kageyama, Asa Morita, Satoshi Seino, Hideto Yoshida, Olivier Jordan, and et al. 2021. "Wound Healing Promotion by Hyaluronic Acid: Effect of Molecular Weight on Gene Expression and In Vivo Wound Closure" Pharmaceuticals 14, no. 4: 301. https://doi.org/10.3390/ph14040301
APA StyleKawano, Y., Patrulea, V., Sublet, E., Borchard, G., Iyoda, T., Kageyama, R., Morita, A., Seino, S., Yoshida, H., Jordan, O., & Hanawa, T. (2021). Wound Healing Promotion by Hyaluronic Acid: Effect of Molecular Weight on Gene Expression and In Vivo Wound Closure. Pharmaceuticals, 14(4), 301. https://doi.org/10.3390/ph14040301