
Valproic Acid Suppresses Autoimmune Recurrence and Allograft Rejection in Islet Transplantation through Induction of the Differentiation of Regulatory T Cells and Can Be Used in Cell Therapy for Type 1 Diabetes


Abstract
:1. Introduction
2. Results
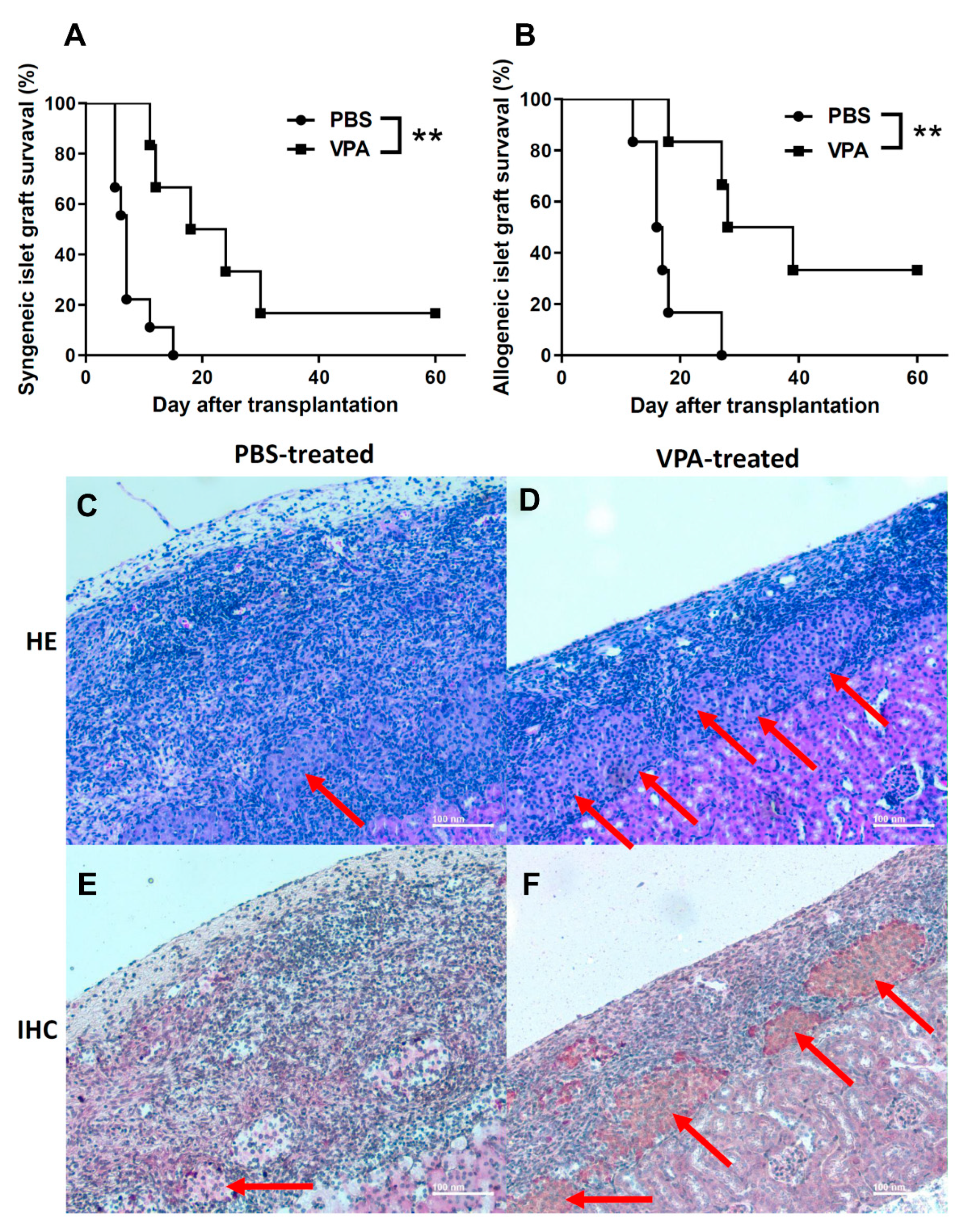
2.1. VPA Treatment Prolonged the Survival of Islet Grafts after Islet Transplantation
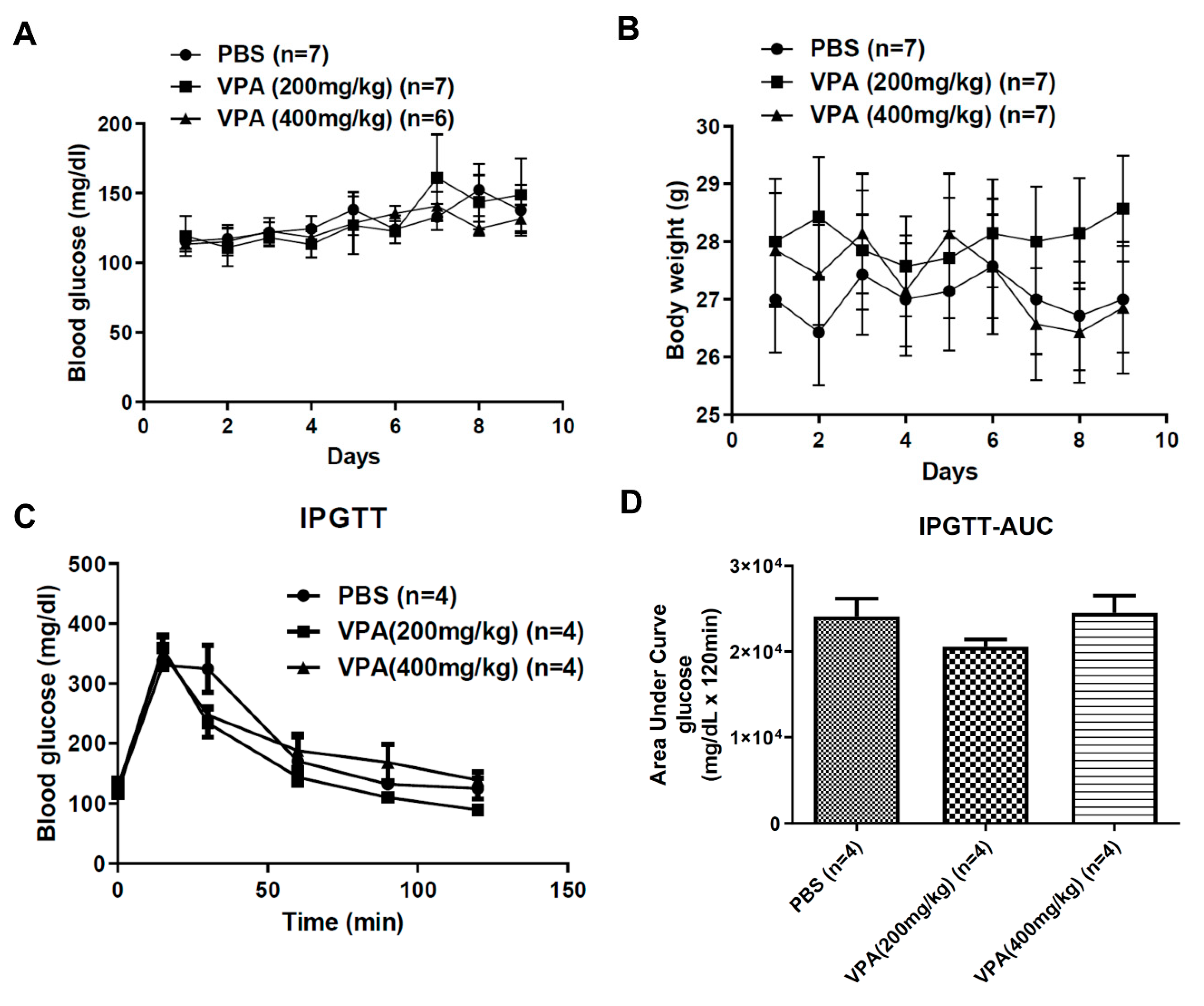
2.2. VPA Treatment Did Not Reverse the Phenotype of Hyperglycemia in Diabetic NOD Mice
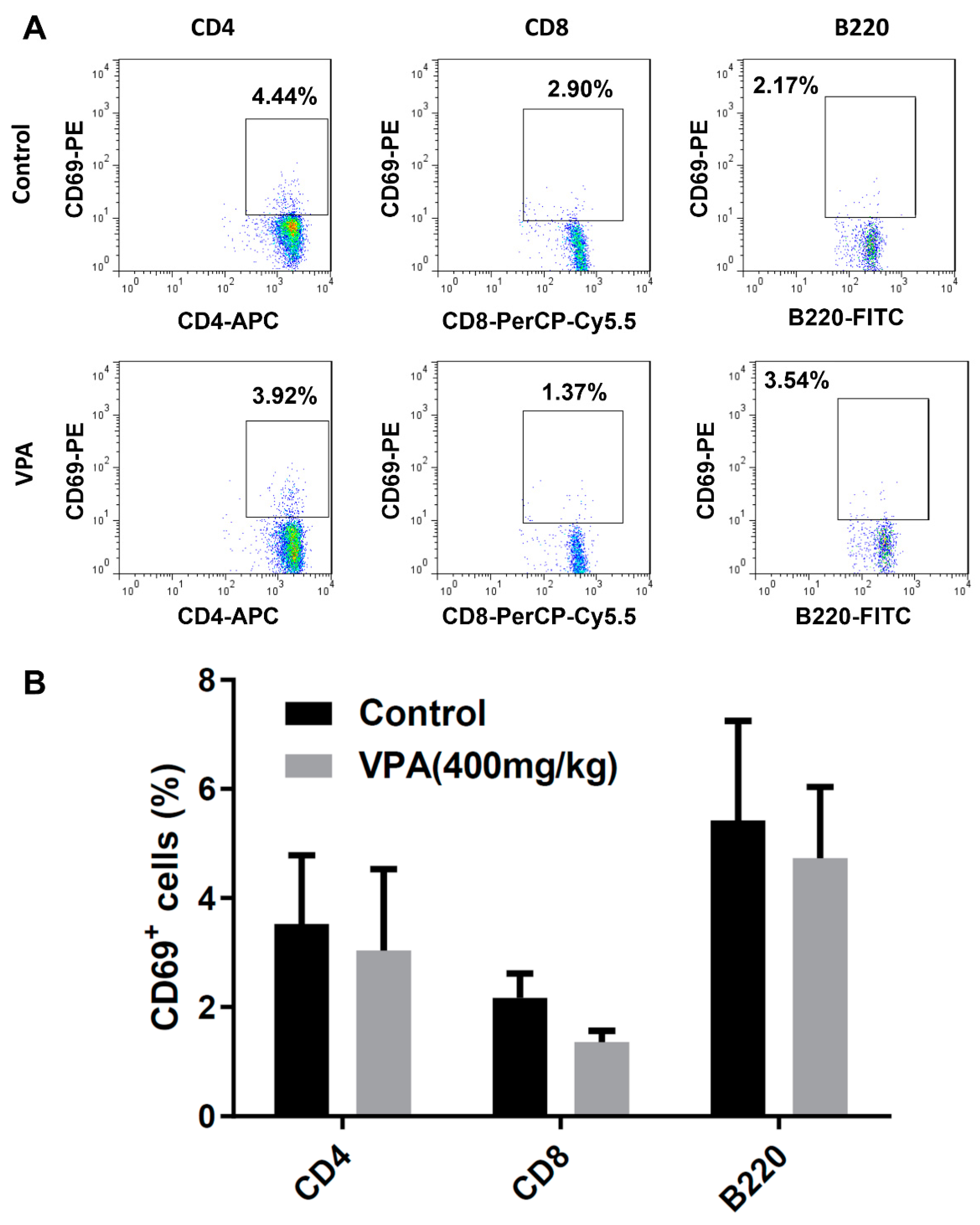
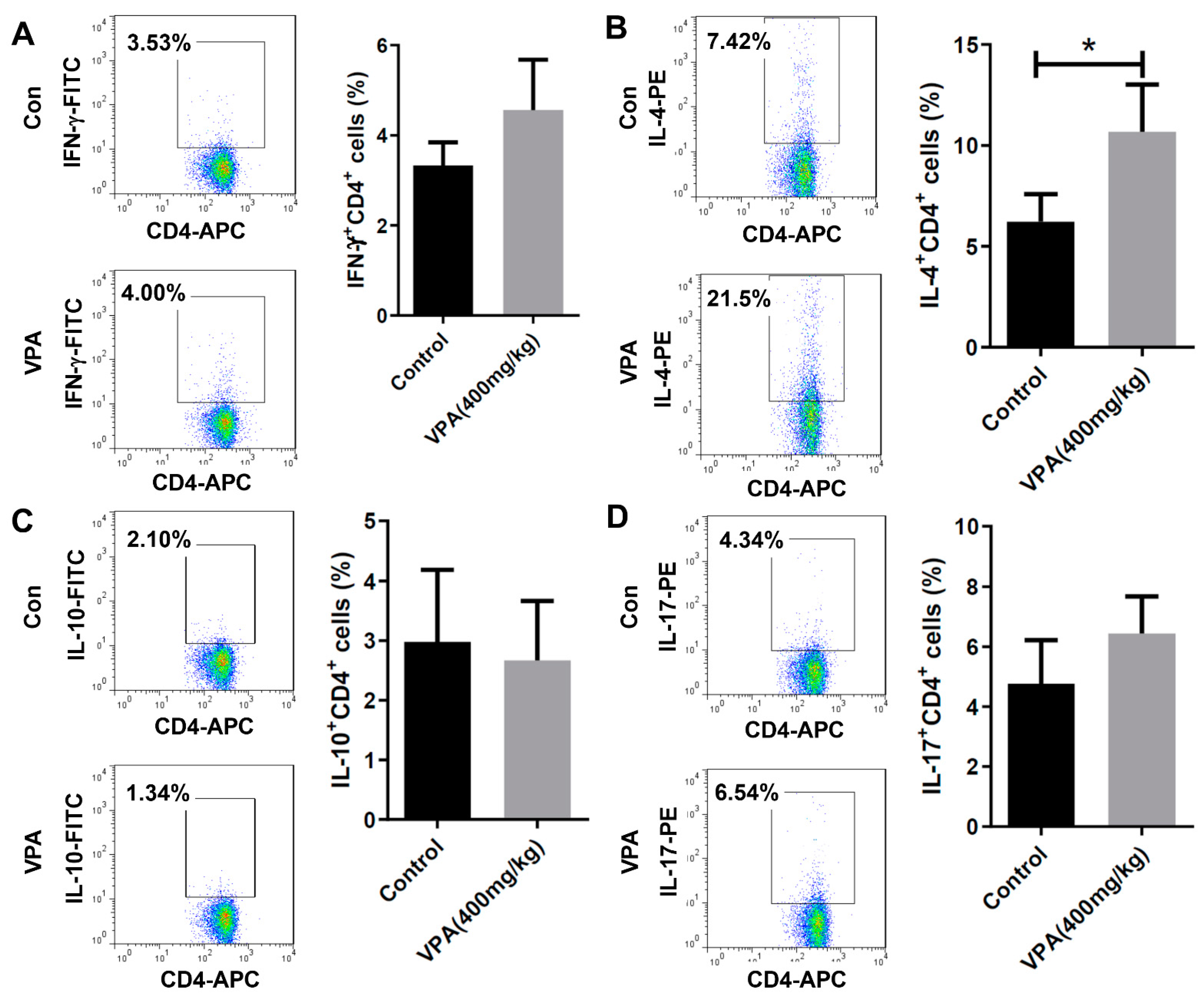
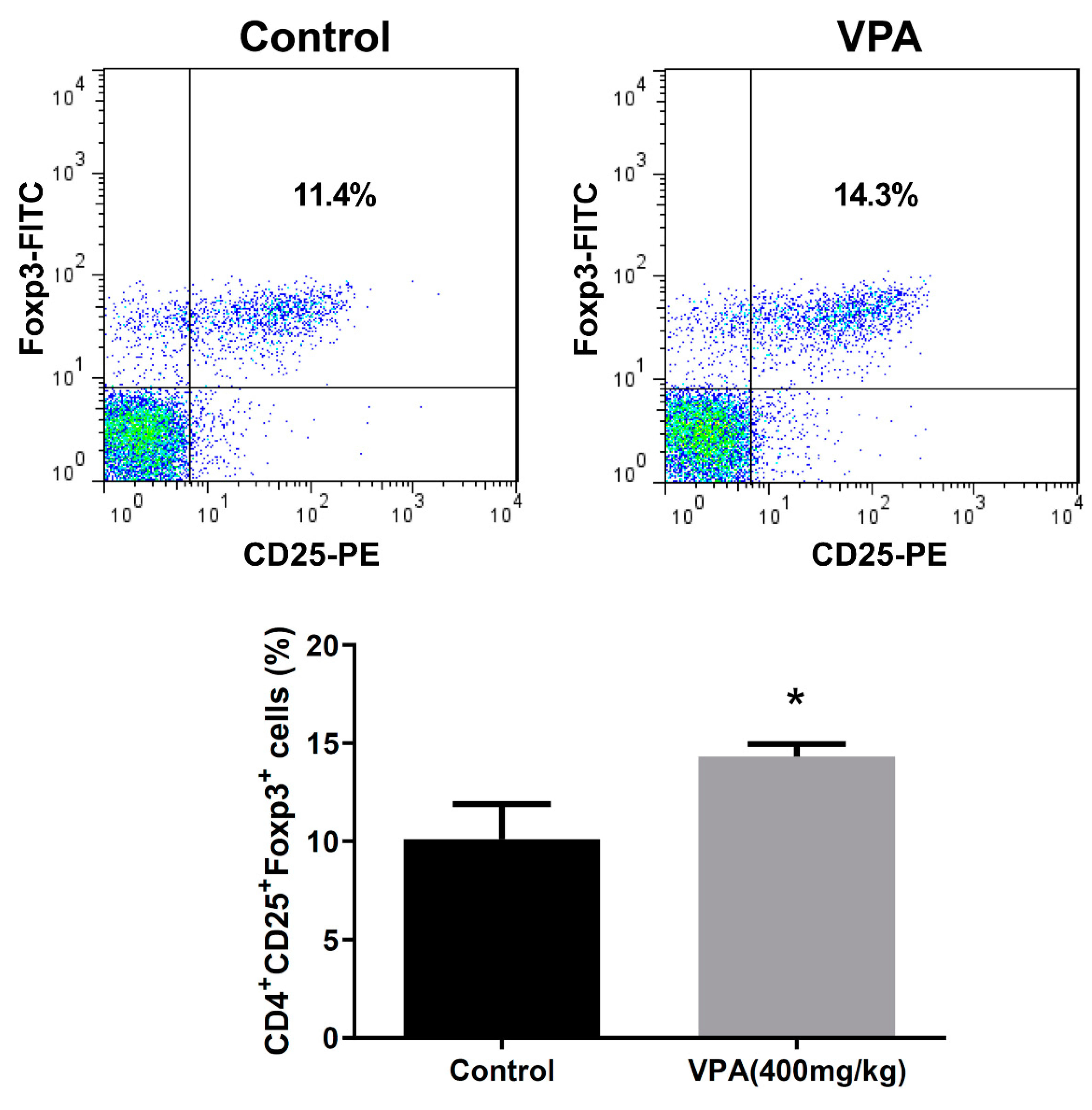
2.3. VPA Increased IL-4-Producing CD4 T Cell and Treg Cell Population in NOD Recipients
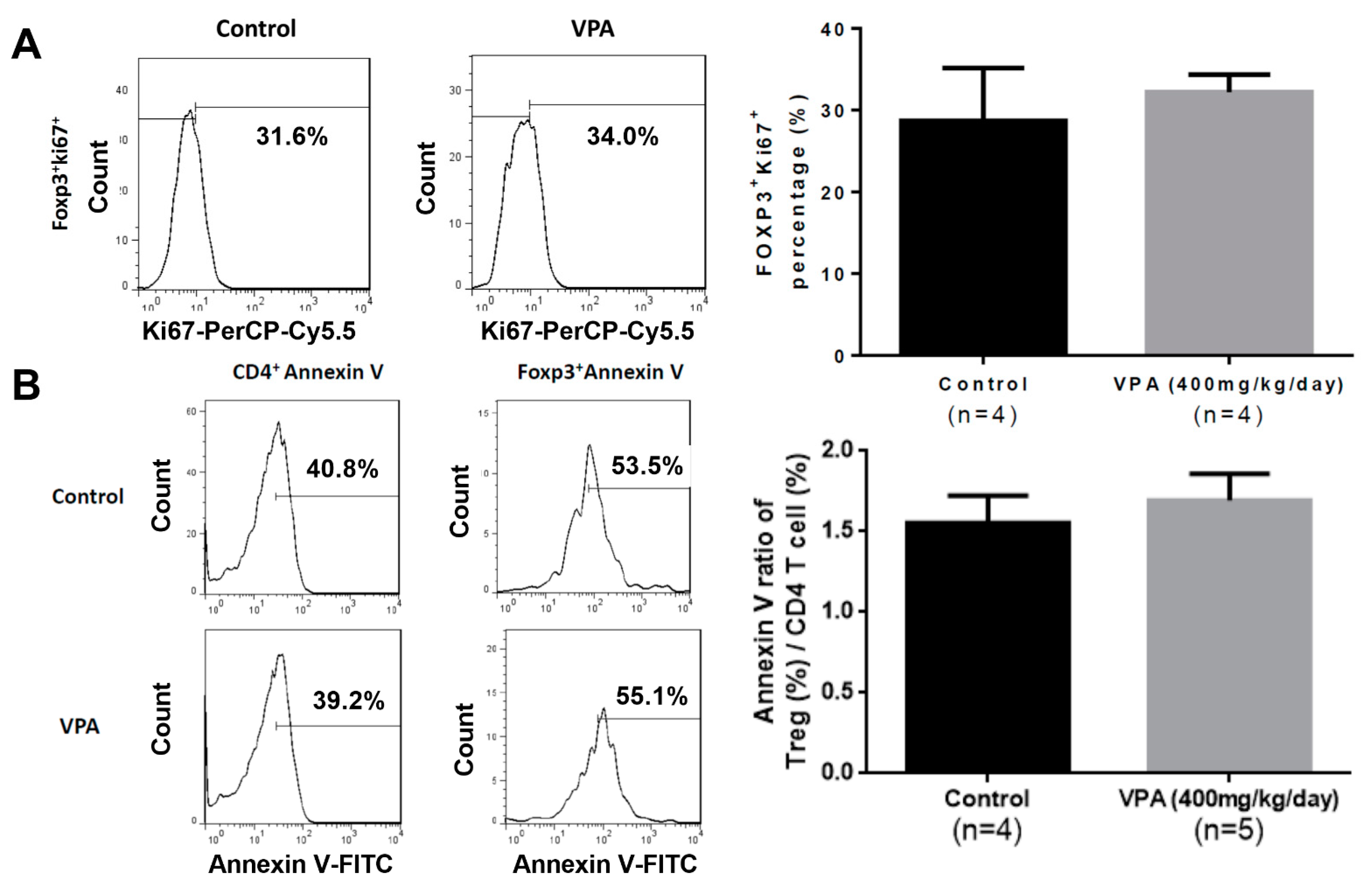
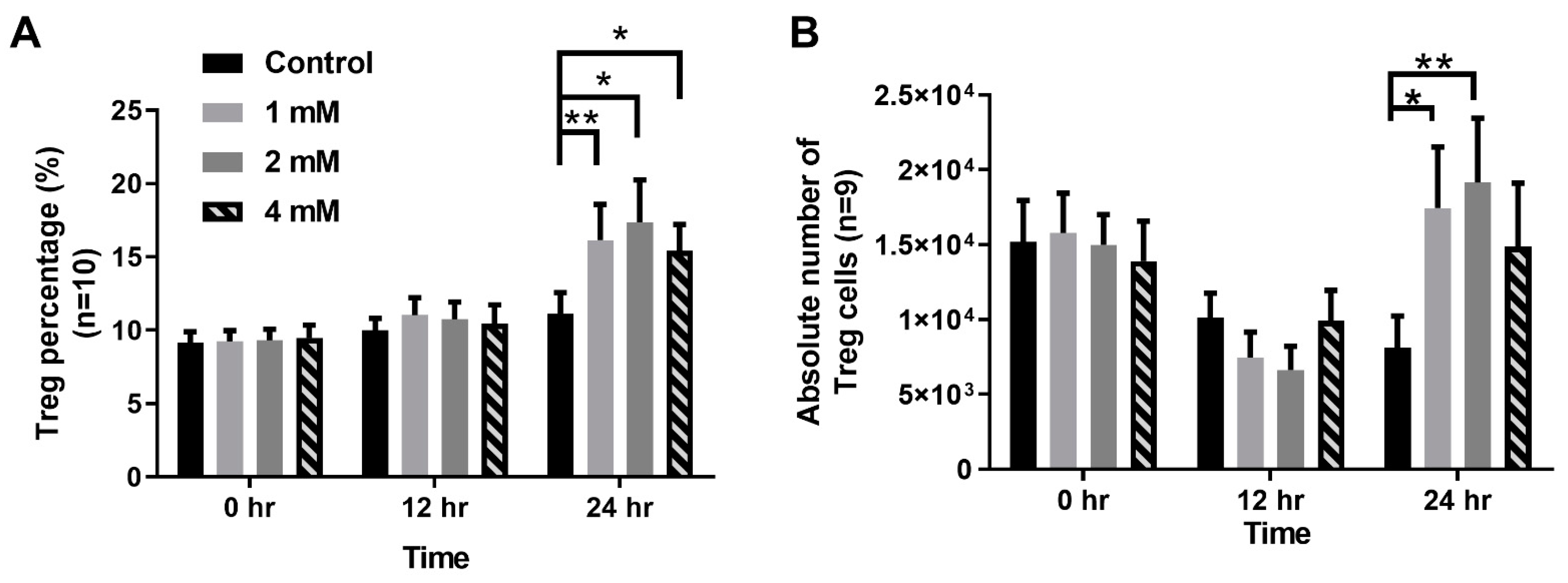
2.4. VPA Treatment Induced Treg Differentiation from Naive CD4 T Cells
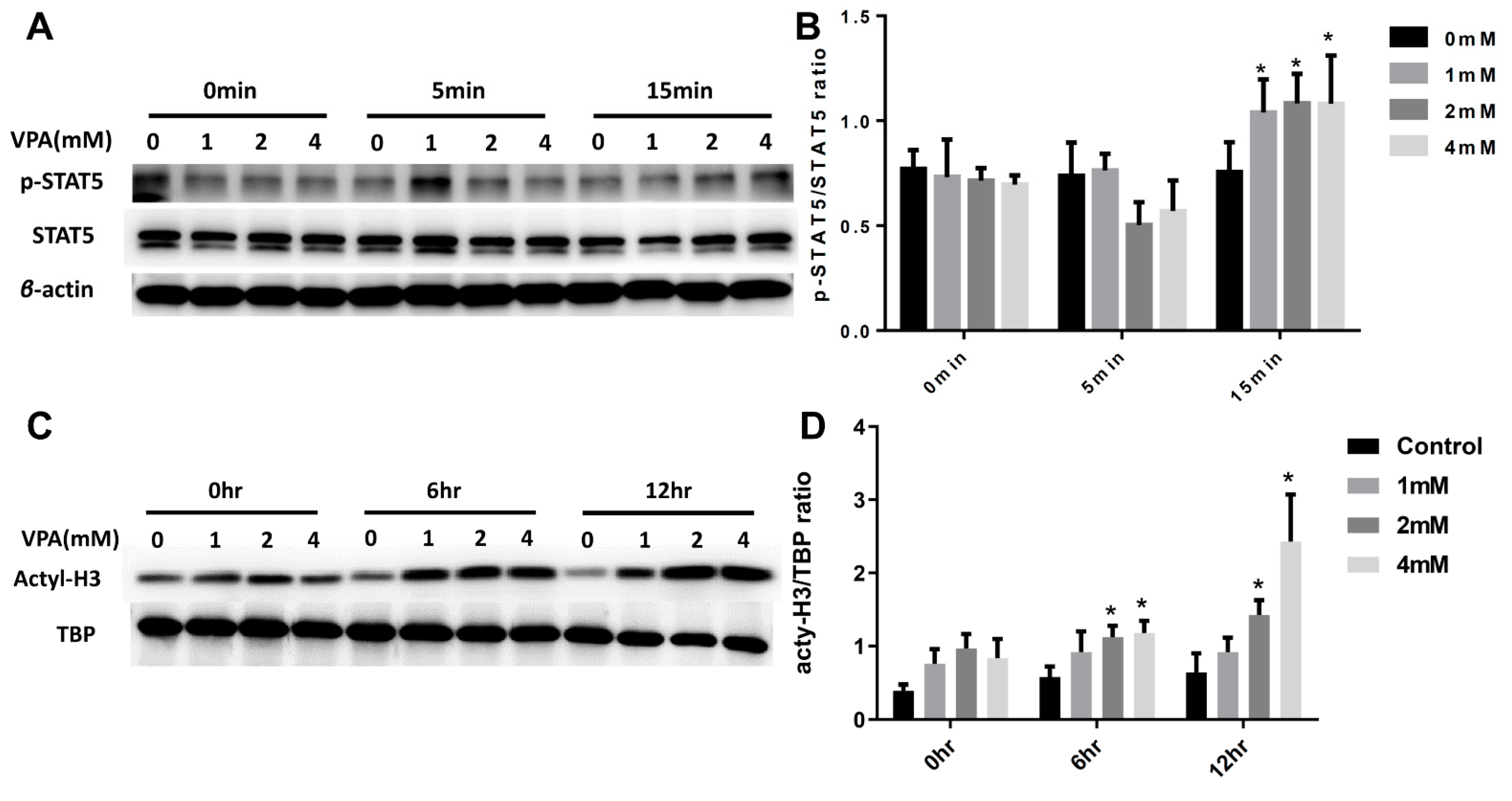
2.5. VPA Induced Treg Cell Differentiation from Naive CD4 T Cells by Increasing the Expression of the Transcription Factor STAT5 and Histone3(H3) Acetylation
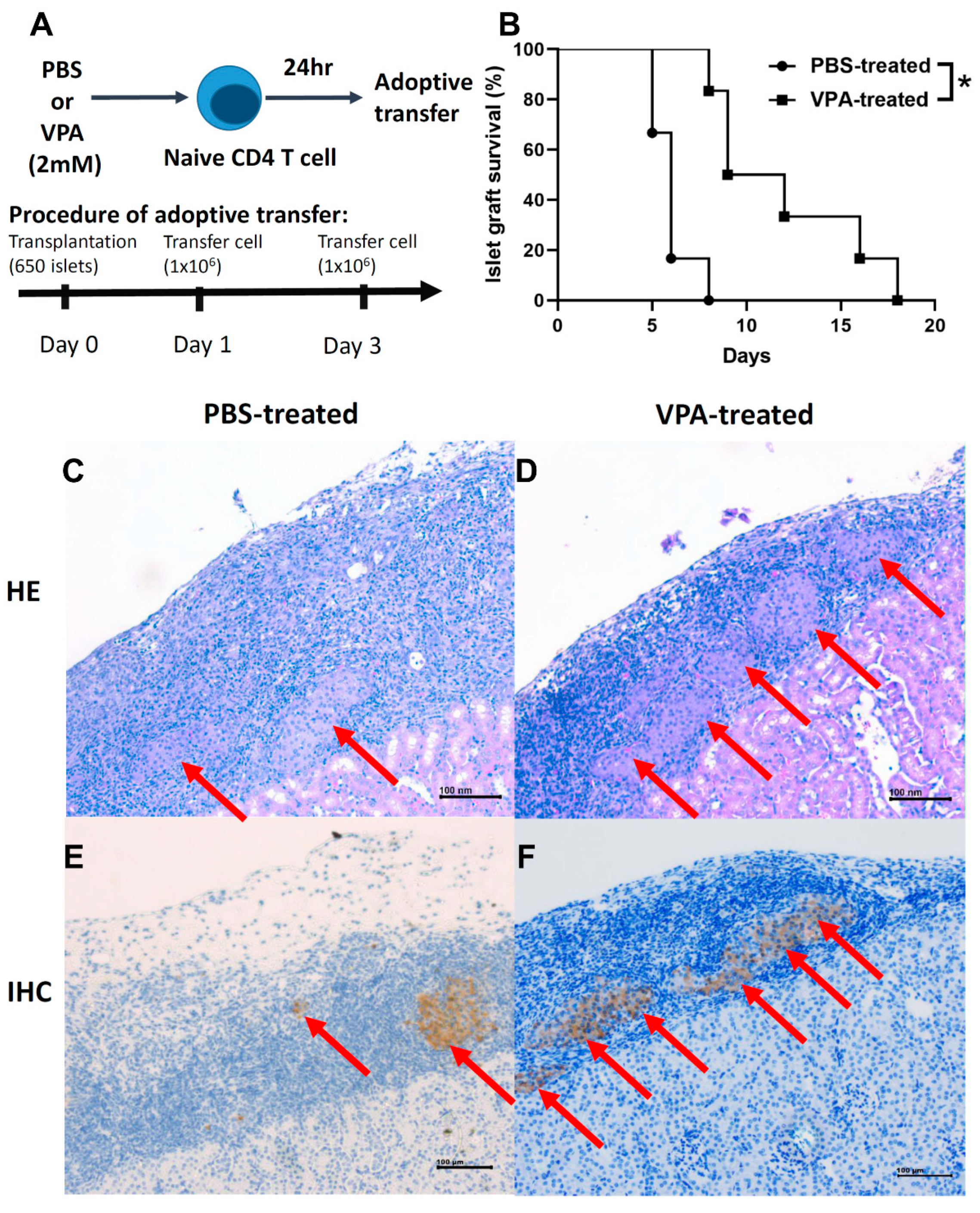
2.6. Adoptive Transfer of In Vitro VPA-Induced Regulatory T Cells Prolonged Islet Graft Survival
3. Discussion
4. Materials and Methods
4.1. Animal Model
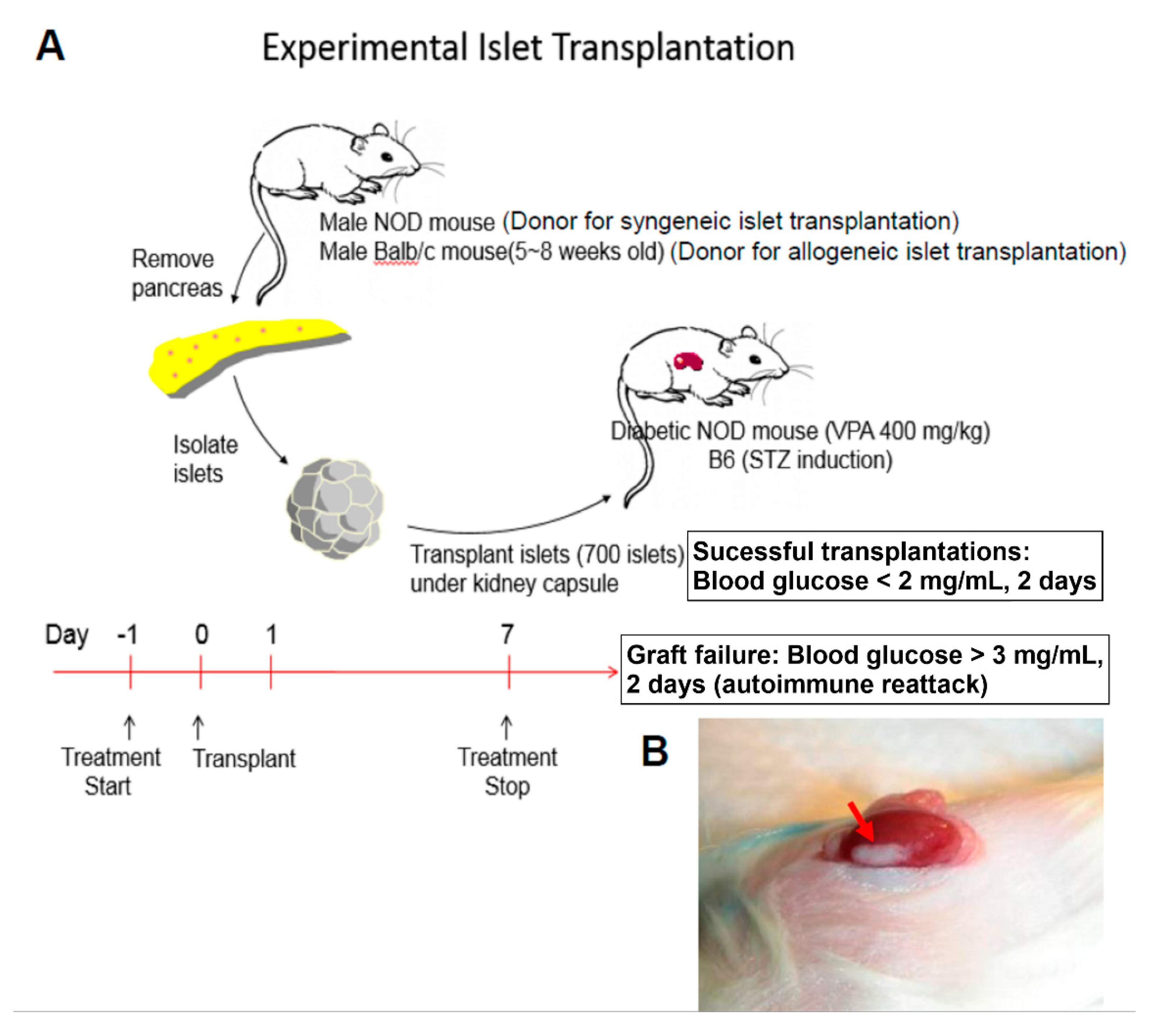
4.2. Islet Isolation and Transplantation
4.3. Blood Glucose Monitoring
4.4. Naive T Cell Sorting
4.5. In Vitro of Treg Cell Differentiation
4.6. T Cell Apoptosis Analysis
4.7. Flow Cytometry
4.8. Low-Dose Streptozotocin(STZ) Induction Diabetes
4.9. Histological and Immunohistochemical Assays
4.10. Adoptive Transfer of Regulatory T Cells
4.11. Protein Extraction and Western Blot
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kawasaki, E.; Abiru, N.; Eguchi, K. Prevention of type 1 diabetes: From the view point of beta cell damage. Diabetes Res. Clin. Pract. 2004, 66 (Suppl. 1), S27–S32. [Google Scholar] [CrossRef]
- Maahs, D.M.; West, N.A.; Lawrence, J.M.; Mayer-Davis, E.J. Epidemiology of type 1 diabetes. J. Clin. Endocrinol. Metab. 2010, 39, 481–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birk, O.S.; Douek, D.C.; Elias, D.; Takacs, K.; Dewchand, H.; Gur, S.L.; Walker, M.D.; van der Zee, R.; Cohen, I.R.; Altmann, D.M. A role of Hsp60 in autoimmune diabetes: Analysis in a transgenic model. Proc. Natl. Acad. Sci. USA 1996, 93, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, S.; Dasu, M.R.; Park, S.H.; Jialal, I. Increased levels of ligands of Toll-like receptors 2 and 4 in type 1 diabetes. Diabetologia 2009, 52, 1665–1668. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Boucher, D.W.; Notkins, A.L. Virus-induced diabetes mellitus. II. Relationship between beta cell damage and hyperglycemia in mice infected with encephalomyocarditis virus. Am. J. Pathol. 1974, 75, 91–102. [Google Scholar] [PubMed]
- Aoki, C.A.; Borchers, A.T.; Ridgway, W.M.; Keen, C.L.; Ansari, A.A.; Gershwin, M.E. NOD mice and autoimmunity. Autoimmun. Rev. 2005, 4, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, R.J.; Dilorenzo, T.P. An update on the use of NOD mice to study autoimmune (Type 1) diabetes. Expert. Rev. Clin. Immunol. 2010, 6, 939–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, J.A.; Wong, F.S.; Wen, L. The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes. J. Autoimmun. 2016, 66, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Salomon, B.; Lenschow, D.J.; Rhee, L.; Ashourian, N.; Singh, B.; Sharpe, A.; Bluestone, J.A. B7/CD28 Costimulation Is Essential for the Homeostasis of the CD4+CD25+ Immunoregulatory T Cells that Control Autoimmune Diabetes. Immunity 2000, 12, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar]
- Bacchetta, R.; Passerini, L.; Gambineri, E.; Dai, M.; Allan, S.E.; Perroni, L.; Dagna-Bricarelli, F.; Sartirana, C.; Matthes-Martin, S.; Lawitschka, A.; et al. Defective regulatory and effector T cell functions in patients with FOXP3 mutations. J. Clin. Investig. 2006, 116, 1713–1722. [Google Scholar] [CrossRef]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A.; Buckner, J.H.; Fitch, M.; Gitelman, S.E.; Gupta, S.; Hellerstein, M.K.; Herold, K.C.; Lares, A.; Lee, M.R.; Li, K.; et al. Type 1 diabetes immunotherapy using polyclonal regulatory T cells. Sci. Transl. Med. 2015, 7, 315ra189. [Google Scholar] [CrossRef] [Green Version]
- Serr, I.; Furst, R.W.; Achenbach, P.; Scherm, M.G.; Gokmen, F.; Haupt, F.; Sedlmeier, E.M.; Knopff, A.; Shultz, L.; Willis, R.A.; et al. Type 1 diabetes vaccine candidates promote human Foxp3(+)Treg induction in humanized mice. Nat. Commun. 2016, 7, 10991. [Google Scholar] [CrossRef]
- Perucca, E. Pharmacological and Therapeutic Properties of Valproate: A Summary After 35 Years of Clinical Experience. CNS Drugs 2002, 16, 695–714. [Google Scholar] [CrossRef] [PubMed]
- Meunier, H.; Carraz, G.; Neunier, Y.; Eymard, P.; Aimard, M. Pharmacodynamic properties of N-dipropylacetic acid. Therapie 1963, 18, 435–438. [Google Scholar] [PubMed]
- BS, B. On the propyl derivatives and decomposition products of ethylacetoacetate. Am. Chem. J. 1882, 3, 385–395. [Google Scholar]
- Chiu, C.T.; Wang, Z.; Hunsberger, J.G.; Chuang, D.M. Therapeutic potential of mood stabilizers lithium and valproic acid: Beyond bipolar disorder. Pharmacol. Rev. 2013, 65, 105–142. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, G.; Wang, L.; Song, C.; Leng, Y.; Wang, X.; Kang, J. VPA inhibits breast cancer cell migration by specifically targeting HDAC2 and down-regulating Survivin. Mol. Cell. Biochem. 2012, 361, 39–45. [Google Scholar] [CrossRef]
- Amirzargar, M.A.; Yaghubi, F.; Hosseinipanah, M.; Jafari, M.; Pourjafar, M.; Rezaeepoor, M.; Rezaei, H.; Roshanaei, G.; Hajilooi, M.; Solgi, G. Anti-inflammatory Effects of Valproic Acid in a Rat Model of Renal Ischemia/Reperfusion Injury: Alteration in Cytokine Profile. Inflammation 2017, 40, 1310–1318. [Google Scholar] [CrossRef]
- Lv, J.; Du, C.; Wei, W.; Wu, Z.; Zhao, G.; Li, Z.; Xie, X. The antiepileptic drug valproic acid restores T cell homeostasis and ameliorates pathogenesis of experimental autoimmune encephalomyelitis. J. Biol. Chem. 2012, 287, 28656–28665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, Z.Y.; Wu, Y.; Schluesener, H.J. Valproic acid ameliorates inflammation in experimental autoimmune encephalomyelitis rats. Neuroscience 2012, 221, 140–150. [Google Scholar] [CrossRef]
- Saouaf, S.J.; Li, B.; Zhang, G.; Shen, Y.; Furuuchi, N.; Hancock, W.W.; Greene, M.I. Deacetylase inhibition increases regulatory T cell function and decreases incidence and severity of collagen-induced arthritis. Exp. Mol. Pathol. 2009, 87, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Epidemiology of diabetes and diabetes-related complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, B.K.; Srivastava, A.K. Diabetes mellitus: Complications and therapeutics. Med. Sci. Monit. 2006, 12, Ra130–Ra147. [Google Scholar] [PubMed]
- Lin, Y.J.; Wang, C.Y.; Cheng, S.W.; Ko, Y. Patient preferences for diabetes-related complications in Taiwan. Curr. Med. Res. Opin. 2019, 35, 7–13. [Google Scholar] [CrossRef]
- Gaglia, J.L.; Shapiro, A.M.; Weir, G.C. Islet transplantation: Progress and challenge. Arch. Med. Res. 2005, 36, 273–280. [Google Scholar] [CrossRef]
- Gamble, A.; Pepper, A.R.; Bruni, A.; Shapiro, A.M.J. The journey of islet cell transplantation and future development. Islets 2018, 10, 80–94. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, A.N.; Bottino, R.; Giannoukakis, N.; Smetanka, C. Prospective and Challenges of Islet Transplantation for the Therapy of Autoimmune Diabetes. Pancreas 2006, 32, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Sibley, R.K.; Sutherland, D.E.; Goetz, F.; Michael, A.F. Recurrent diabetes mellitus in the pancreas iso- and allograft. A light and electron microscopic and immunohistochemical analysis of four cases. Lab. Investig. 1985, 53, 132–144. [Google Scholar]
- Tyden, G.; Reinholt, F.P.; Sundkvist, G.; Bolinder, J. Recurrence of autoimmune diabetes mellitus in recipients of cadaveric pancreatic grafts. N. Engl. J. Med. 1996, 335, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Haque, I.; Tayal, V.; Roy, V. Valproic acid-induced acute pancreatitis. Indian J. Psychol. Med. 2019, 61, 421–422. [Google Scholar] [CrossRef]
- Zhu, X.; Li, X.; Zhang, T.; Zhao, L. Risk Factors for Valproic Acid-induced Hyperammonaemia in Chinese Paediatric Patients with Epilepsy. Basic Clin. Pharmacol. Toxicol. 2018, 123, 628–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, Y.; Limberis, M.P.; Zhang, H. Modulation of Treg function improves adenovirus vector-mediated gene expression in the airway. Gene Ther. 2014, 21, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Passerini, L.; Allan, S.E.; Battaglia, M.; Di Nunzio, S.; Alstad, A.N.; Levings, M.K.; Roncarolo, M.G.; Bacchetta, R. STAT5-signaling cytokines regulate the expression of FOXP3 in CD4+CD25+ regulatory T cells and CD4+CD25- effector T cells. Int. Immunol. 2008, 20, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Nosaka, T.; Misawa, K.; Mui, A.L.; Gorman, D.; McMahon, M.; Miyajima, A.; Kitamura, T. Identification and characterization of a constitutively active STAT5 mutant that promotes cell proliferation. Mol. Cell. Biol. 1998, 18, 3871–3879. [Google Scholar] [CrossRef] [Green Version]
- Gruessner, A.C.; Sutherland, D.E. Pancreas transplant outcomes for United States (US) and non-US cases as reported to the United Network for Organ Sharing (UNOS) and the International Pancreas Transplant Registry (IPTR) as of June 2004. Clin. Transplant. 2005, 19, 433–455. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.Y.; Fauser, U.; Schluesener, H.J. Valproic acid attenuates inflammation in experimental autoimmune neuritis. Cell Mol. Life Sci. 2008, 65, 4055–4065. [Google Scholar] [CrossRef]
- Ye, J.; Li, J.; Zhou, M.; Xia, R.; Liu, R.; Yu, L. Modulation of Donor-Specific Antibody Production after Organ Transplantation by Valproic Acid: A Histone Deacetylase Inhibitor. Transplantation 2016, 100, 2342–2351. [Google Scholar] [CrossRef]
- Nanau, R.M.; Neuman, M.G. Adverse drug reactions induced by valproic acid. Clin. Biochem. 2013, 46, 1323–1338. [Google Scholar] [CrossRef]
- Kanemura, H.; Sano, F.; Maeda, Y.; Sugita, K.; Aihara, M. Valproate sodium enhances body weight gain in patients with childhood epilepsy: A pathogenic mechanisms and open-label clinical trial of behavior therapy. Seizure 2012, 21, 496–500. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.M.; Reiner, S.L. The lineage decisions of helper T cells. Nat. Rev. Immunol. 2002, 2, 933–944. [Google Scholar] [CrossRef]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006, 24, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Guo, X. Valproic acid ameliorates coxsackievirus-B3-induced viral myocarditis by modulating Th17/Treg imbalance. Virol. J. 2016, 13, 168. [Google Scholar] [CrossRef] [Green Version]
- Jerram, S.T.; Dang, M.N.; Leslie, R.D. The Role of Epigenetics in Type 1 Diabetes. Curr. Diab. Rep. 2017, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- San Jose-Eneriz, E.; Gimenez-Camino, N.; Agirre, X.; Prosper, F. HDAC Inhibitors in Acute Myeloid Leukemia. Cancers 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, B.; Farlik, M.; Sexl, V. It is a differentiation game: STAT5 in a new role. Cell Death Differ. 2017, 24, 953–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gittes, R.F.; Chu, E.W. Reversal of the effect of pinealectomy in female rats by multiple isogeneic pineal transplants. Endocrinology 1965, 77, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, C.; Lama, A.; Annunziata, C.; Cavaliere, G.; De Caro, C.; Citraro, R.; Russo, E.; Tallarico, M.; Iannone, M.; Ferrante, M.C.; et al. Butyrate prevents valproate-induced liver injury: In vitro and in vivo evidence. FASEB J 2020, 34, 676–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.L.; Jing, X.; Sun, J.Y.; Hu, Y.H.; Xu, Z.J.; Ni, M.M.; Chen, F.; Lu, X.P.; Qiu, J.C.; Wang, T. Valproic Acid and the Liver Injury in Patients with Epilepsy: An Update. Curr. Pharm. Des. 2019, 25, 343–351. [Google Scholar] [CrossRef]
- Juang, J.H.; Kuo, C.H.; Hsu, B.R. Effects of multiple site implantation on islet transplantation. Transplant. Proc. 2002, 34, 2698–2699. [Google Scholar] [CrossRef]
- Lin, G.J.; Wu, C.H.; Yu, C.C.; Lin, J.R.; Liu, X.D.; Chen, Y.W.; Chang, H.M.; Hong, Z.J.; Cheng, C.P.; Sytwu, H.K.; et al. Adoptive transfer of DMSO-induced regulatory T cells exhibits a similar preventive effect compared to an in vivo DMSO treatment for chemical-induced experimental encapsulating peritoneal sclerosis in mice. Toxicol. Appl. Pharmacol. 2019, 378, 114641. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Individual Graft Survival Time (Days) | Number | Average Survival Time |
|---|---|---|---|
| PBS | 5, 5, 5, 6, 7, 7, 7, 11, 15 | 9 | 7.55 |
| VPA | 11, 12, 18, 24, 30, 63 | 6 | 26.33 |
| Group | Individual Graft Survival Time (Days) | Number | Average Survival Time |
|---|---|---|---|
| PBS | 12, 16, 16, 17, 18, 27 | 6 | 17.66 |
| VPA | 18, 27, 28, 39, 67, 82 | 6 | 43.5 |
| Group | Individual Graft Survival Time (Days) | Number | Average Survival Time |
|---|---|---|---|
| PBS | 5, 5, 6, 6, 6, 8 | 6 | 6 |
| VPA | 8, 9, 9, 12, 16, 18 | 6 | 10.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, J.-R.; Huang, S.-H.; Wu, C.-H.; Chen, Y.-W.; Hong, Z.-J.; Cheng, C.-P.; Sytwu, H.-K.; Lin, G.-J. Valproic Acid Suppresses Autoimmune Recurrence and Allograft Rejection in Islet Transplantation through Induction of the Differentiation of Regulatory T Cells and Can Be Used in Cell Therapy for Type 1 Diabetes. Pharmaceuticals 2021, 14, 475. https://doi.org/10.3390/ph14050475
Lin J-R, Huang S-H, Wu C-H, Chen Y-W, Hong Z-J, Cheng C-P, Sytwu H-K, Lin G-J. Valproic Acid Suppresses Autoimmune Recurrence and Allograft Rejection in Islet Transplantation through Induction of the Differentiation of Regulatory T Cells and Can Be Used in Cell Therapy for Type 1 Diabetes. Pharmaceuticals. 2021; 14(5):475. https://doi.org/10.3390/ph14050475
Chicago/Turabian StyleLin, Jeng-Rong, Shing-Hwa Huang, Chih-Hsiung Wu, Yuan-Wu Chen, Zhi-Jie Hong, Chia-Pi Cheng, Huey-Kang Sytwu, and Gu-Jiun Lin. 2021. "Valproic Acid Suppresses Autoimmune Recurrence and Allograft Rejection in Islet Transplantation through Induction of the Differentiation of Regulatory T Cells and Can Be Used in Cell Therapy for Type 1 Diabetes" Pharmaceuticals 14, no. 5: 475. https://doi.org/10.3390/ph14050475
APA StyleLin, J. -R., Huang, S. -H., Wu, C. -H., Chen, Y. -W., Hong, Z. -J., Cheng, C. -P., Sytwu, H. -K., & Lin, G. -J. (2021). Valproic Acid Suppresses Autoimmune Recurrence and Allograft Rejection in Islet Transplantation through Induction of the Differentiation of Regulatory T Cells and Can Be Used in Cell Therapy for Type 1 Diabetes. Pharmaceuticals, 14(5), 475. https://doi.org/10.3390/ph14050475