
Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome

 , ,
, , 
Abstract
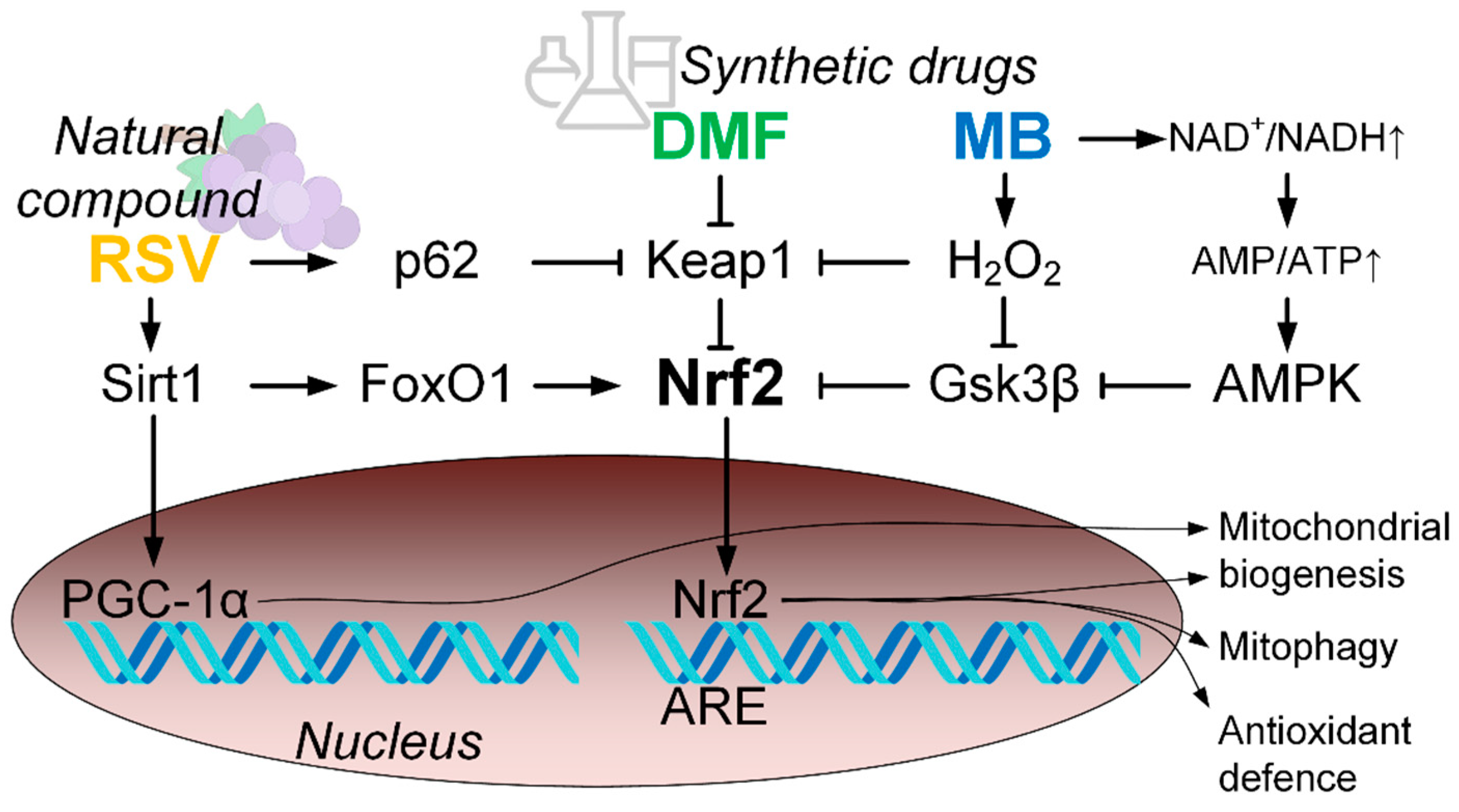
:1. Introduction
2. Results
2.1. Physiological Tests
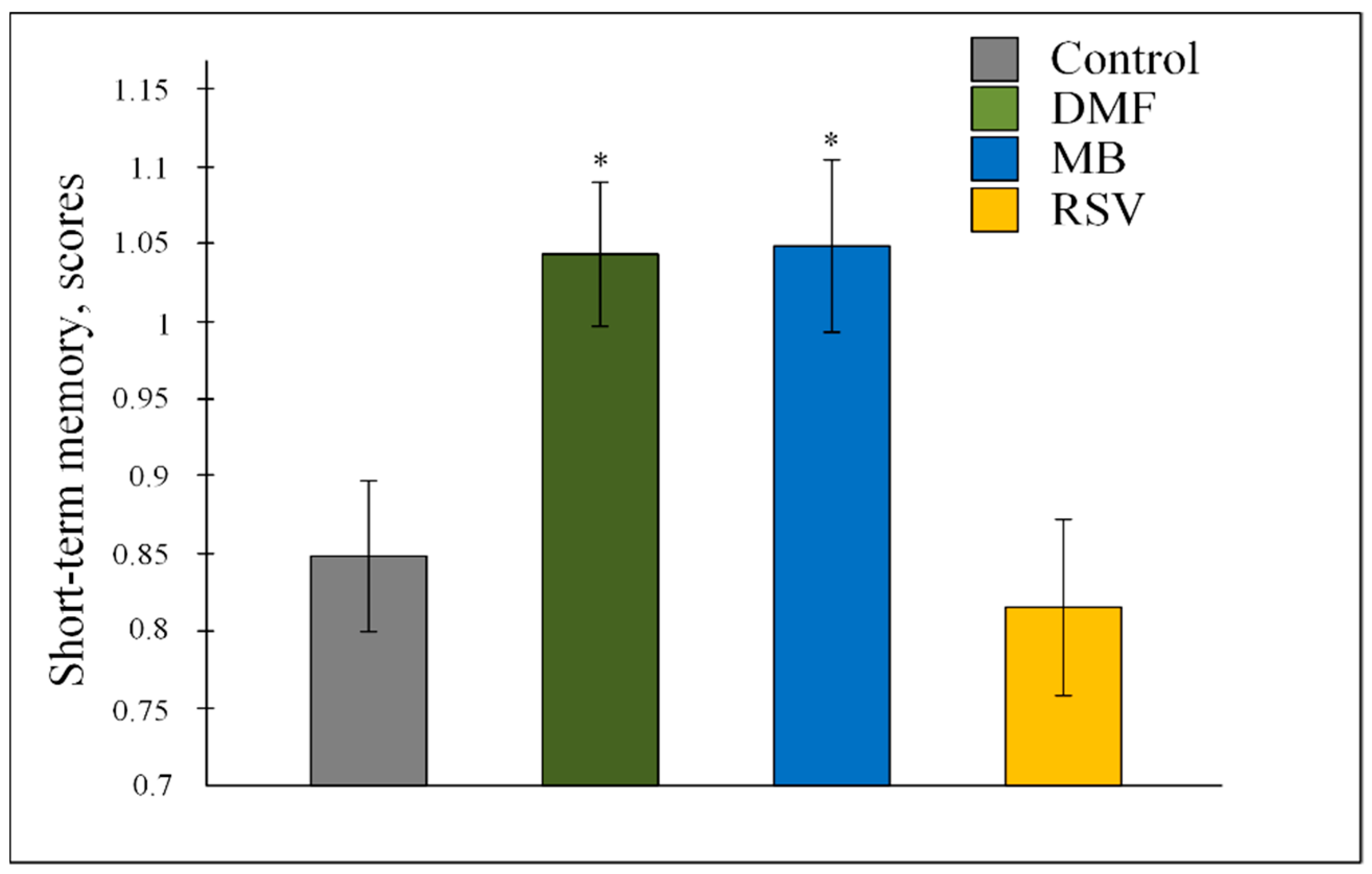
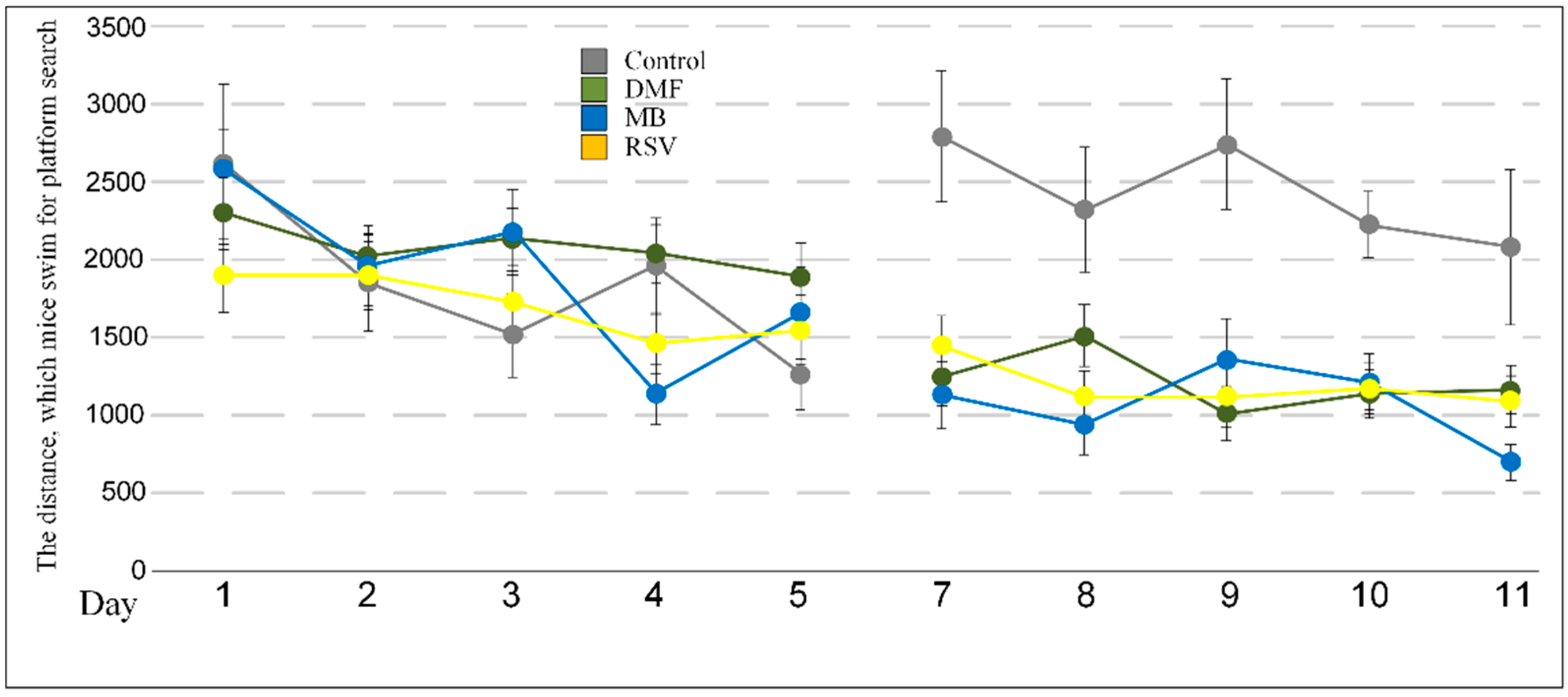
2.2. Memory
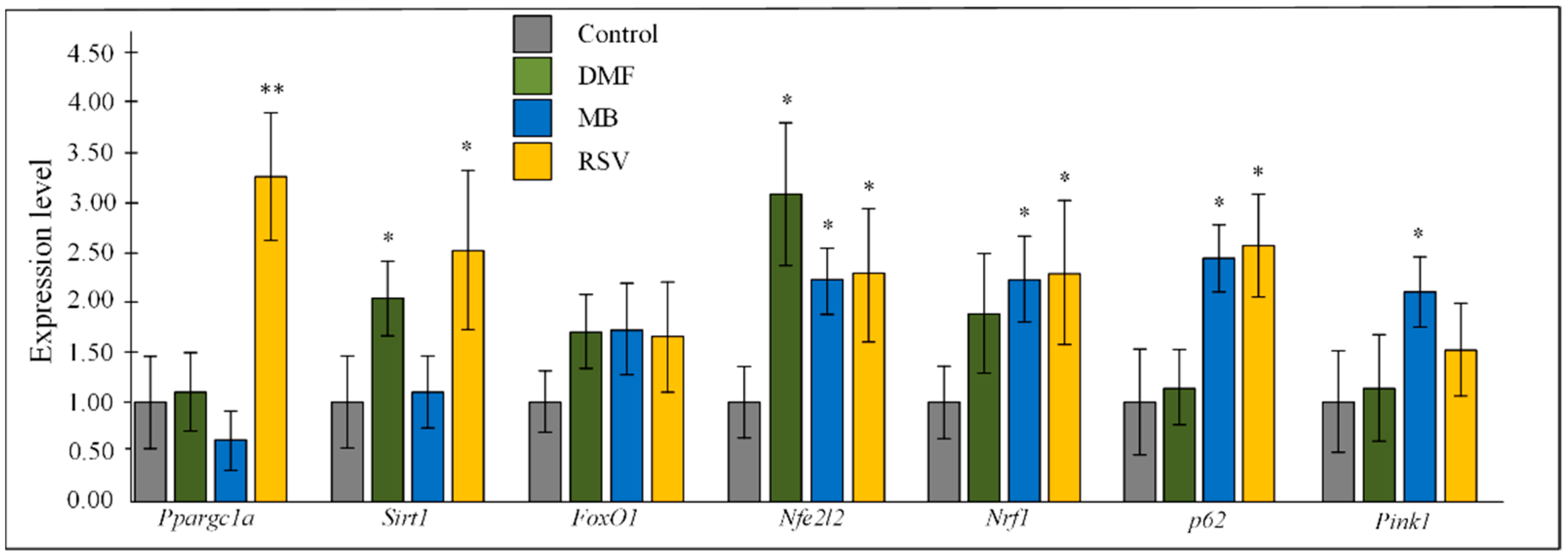
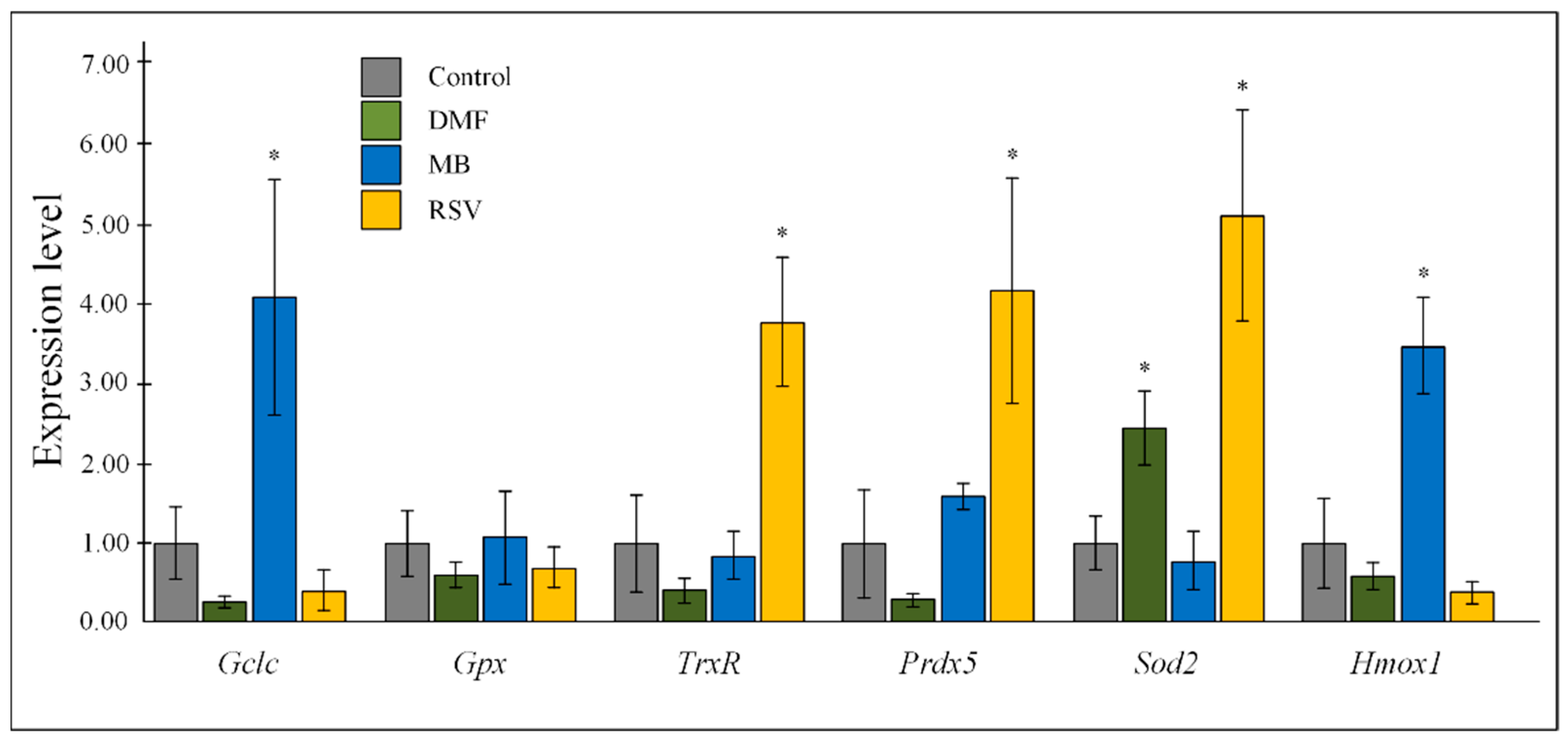
2.3. Gene Expression
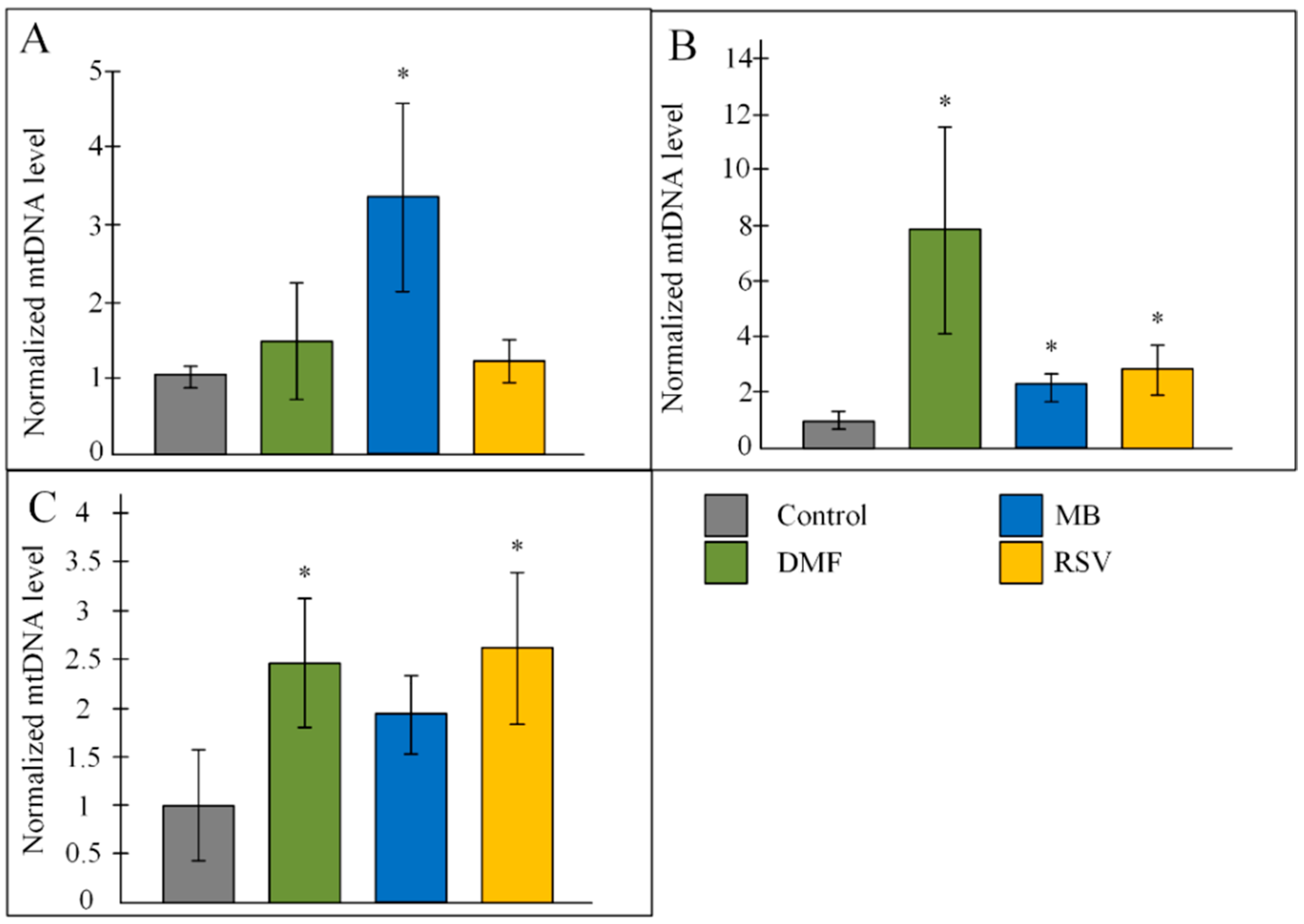
2.4. Copy Number of mtDNA
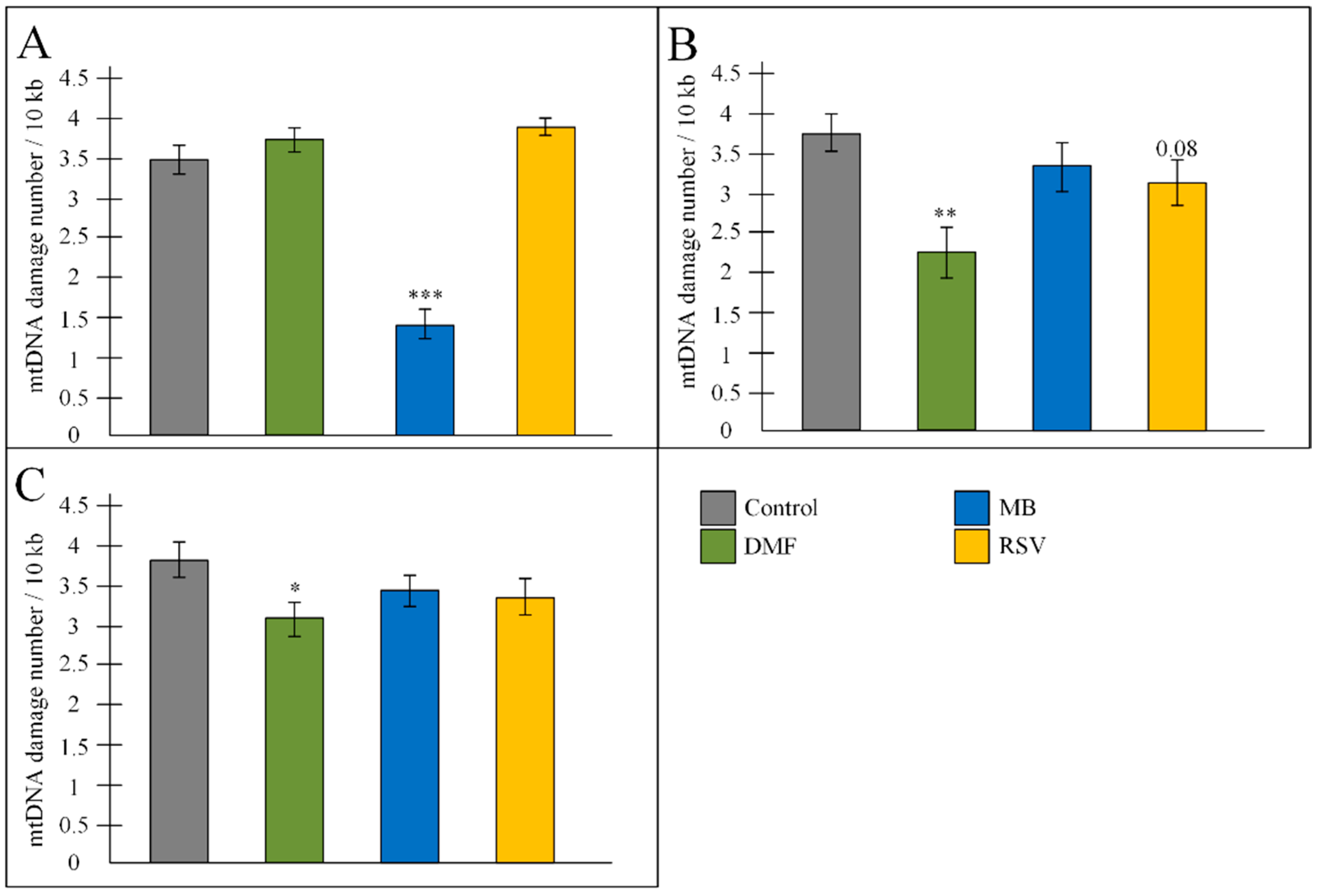
2.5. mtDNA Damage
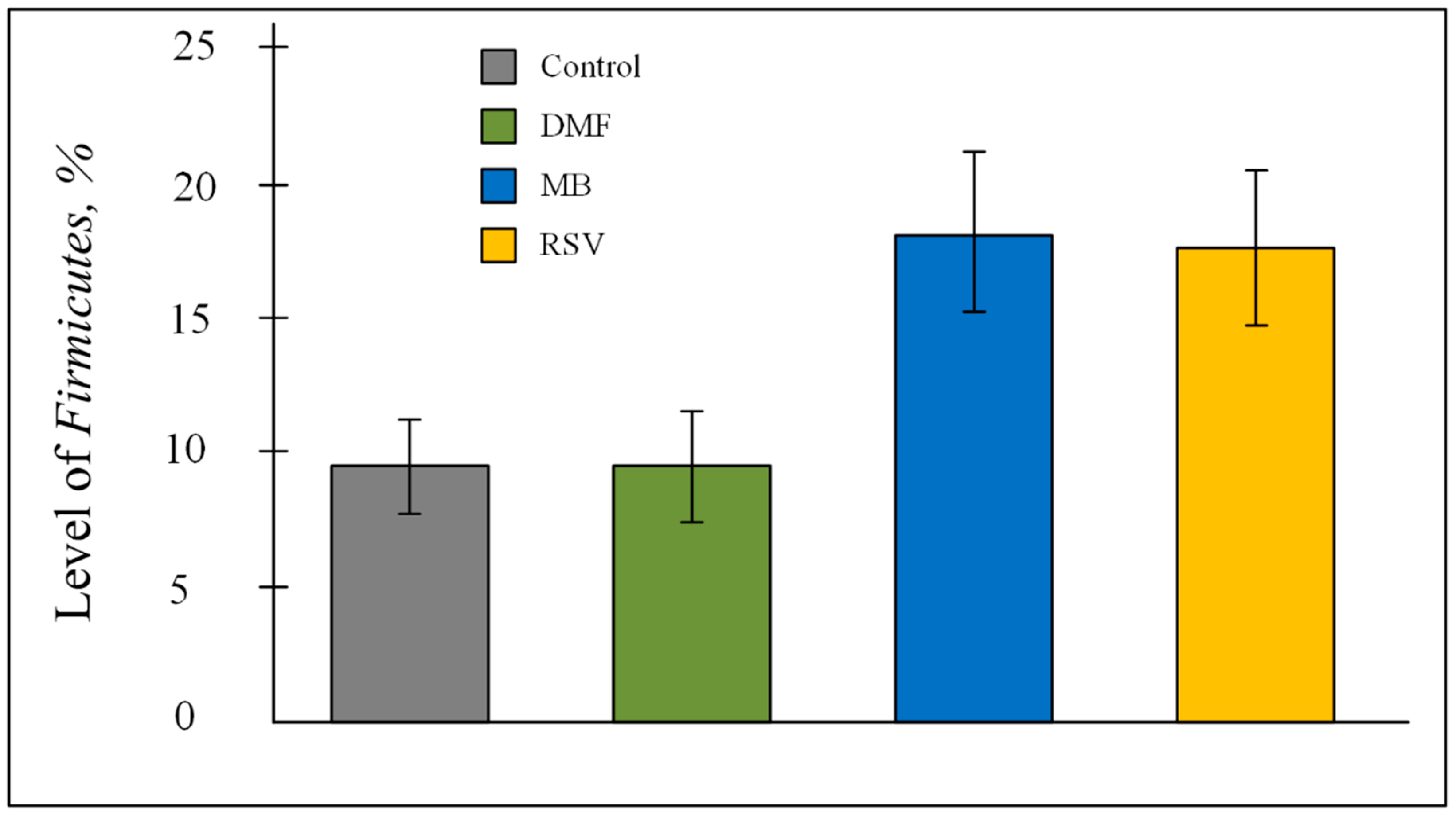
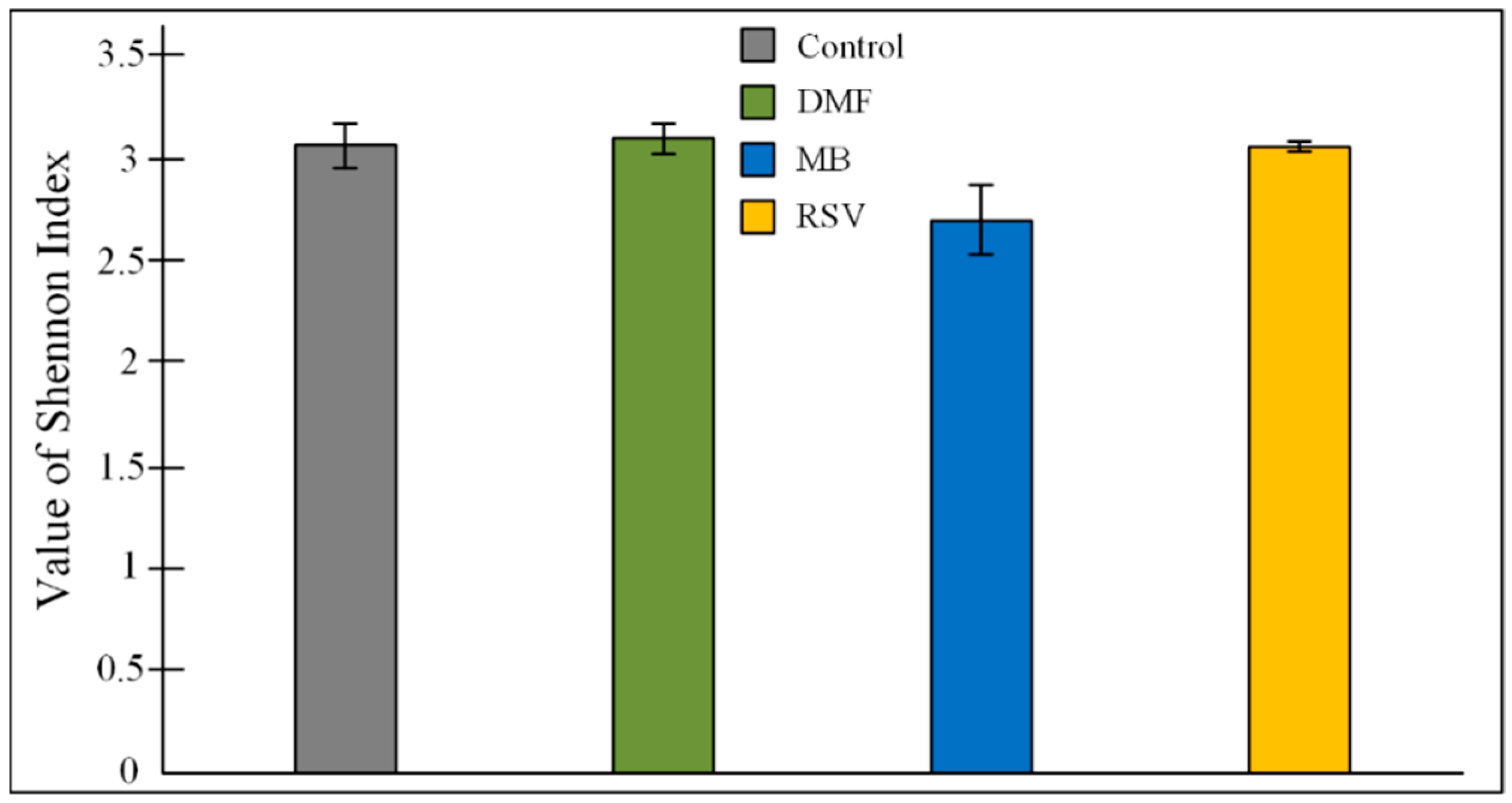
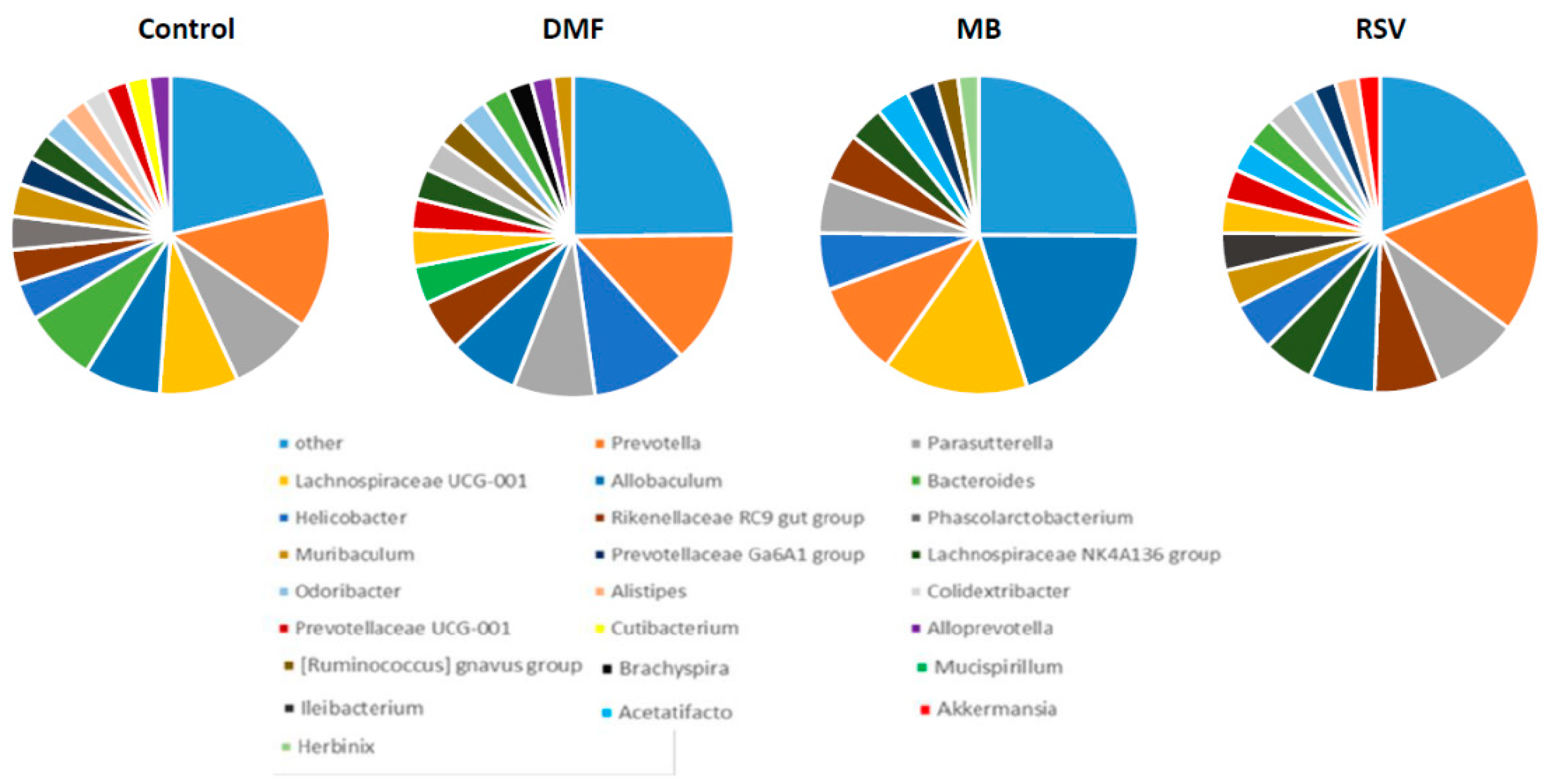
2.6. Bacterial Composition of Gut Microbiome
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Physiological Tests
4.3. Morris Water Maze
4.4. DNA and RNA Isolation
4.5. Measurement of mtDNA Copy Number
4.6. mtDNA Damage Measurement
4.7. Gene Expression Analysis
4.8. Analysis of Gut Microbiome Using PCR
4.9. Analysis of Gut Microbiome Using NGS
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Edwards, G.A., III; Gamez, N.; Escobedo, G., Jr.; Calderon, O.; Moreno-Gonzalez, I. Modifiable Risk Factors for Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Terada, S.; Nakashima, M.; Wakutani, Y.; Nakata, K.; Kutoku, Y.; Sunada, Y.; Kondo, K.; Ishizu, H.; Yokota, O.; Maki, Y.; et al. Social problems in daily life of patients with dementia. Geriatr. Gerontol. Int. 2019, 19, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Yeu, Y.; Yoon, Y.; Park, S. Protein localization vector propagation: A method for improving the accuracy of drug repositioning. Mol. Biosyst. 2015, 11, 2096–2102. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Li, J.; Xie, H.; Wang, Y. Review of Drug Repositioning Approaches and Resources. Int. J. Biol. Sci. 2018, 14, 1232–1244. [Google Scholar] [CrossRef] [Green Version]
- Mrowietz, U.; Morrison, P.J.; Suhrkamp, I.; Kumanova, M.; Clement, B. The Pharmacokinetics of Fumaric Acid Esters Reveal Their In Vivo Effects. Trends Pharmacol. Sci. 2018, 39, 1–12. [Google Scholar] [CrossRef]
- Cada, D.J.; Levien, T.L.; Baker, D.E. Dimethyl fumarate. Hosp. Pharm. 2013, 48, 668–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourakis, S.; Timpani, C.A.; de Haan, J.B.; Gueven, N.; Fischer, D.; Rybalka, E. Dimethyl Fumarate and Its Esters: A Drug with Broad Clinical Utility? Pharmaceuticals 2020, 13, 306. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, R.H.; Adler, H.; Pickhardt, M.; Mandelkow, E. “Lest we forget you--methylene blue…”. Neurobiol. Aging 2011, 32, 2325.e7–2325.e16. [Google Scholar] [CrossRef]
- Peter, C.; Hongwan, D.; Küpfer, A.; Lauterburg, B.H. Pharmacokinetics and organ distribution of intravenous and oral methylene blue. Eur. J. Clin. Pharmacol. 2000, 56, 247–250. [Google Scholar] [CrossRef]
- Skold, A.; Cosco, D.L.; Klein, R. Methemoglobinemia: Pathogenesis, diagnosis, and management. South Med. J. 2011, 104, 757–761. [Google Scholar] [CrossRef]
- Lu, G.; Nagbanshi, M.; Goldau, N.; Mendes Jorge, M.; Meissner, P.; Jahn, A.; Mockenhaupt, F.P.; Müller, O. Efficacy and safety of methylene blue in the treatment of malaria: A systematic review. BMC Med. 2018, 16, 59. [Google Scholar] [CrossRef]
- ClinicalTrials.gov Identifier: NCT01689246. Available online: https://clinicaltrials.gov/ct2/show/NCT01689246 (accessed on 29 April 2021).
- Hashweh, N.N.; Bartochowski, Z.; Khoury, R.; Grossberg, G.T. An evaluation of hydromethylthionine as a treatment option for Alzheimer’s disease. Expert Opin. Pharmacother. 2020, 21, 619–627. [Google Scholar] [CrossRef]
- ClinicalTrials.gov Identifier: NCT03446001. Available online: https://clinicaltrials.gov/ct2/show/NCT03446001 (accessed on 29 April 2021).
- ClinicalTrials.gov Identifier: NCT02380573. Available online: https://clinicaltrials.gov/ct2/show/NCT02380573 (accessed on 29 April 2021).
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef]
- Lin, H.S.; Ho, P.C. Preclinical pharmacokinetic evaluation of resveratrol trimethyl ether in sprague-dawley rats: The impacts of aqueous solubility, dose escalation, food and repeated dosing on oral bioavailability. J. Pharm. Sci. 2011, 100, 4491–4500. [Google Scholar] [CrossRef]
- ClinicalTrials.gov Identifier: NCT04314739. Available online: https://clinicaltrials.gov/ct2/show/NCT04314739 (accessed on 29 April 2021).
- Dinkova-Kostova, A.T.; Abramov, A.Y. The emerging role of Nrf2 in mitochondrial function. Free Radic. Biol. Med. 2015, 88 Pt B, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N. Regulation of Mitochondrial Biogenesis as a Way for Active Longevity: Interaction between the Nrf2 and PGC-1α Signaling Pathways. Front. Genet. 2019, 10, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saji, N.; Murotani, K.; Hisada, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Niida, S.; Toba, K.; Sakurai, T. The relationship between the gut microbiome and mild cognitive impairment in patients without dementia: A cross-sectional study conducted in Japan. Sci. Rep. 2019, 9, 19227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Jaber, V.; Lukiw, W.J. Secretory Products of the Human GI Tract Microbiome and Their Potential Impact on Alzheimer’s Disease (AD): Detection of Lipopolysaccharide (LPS) in AD Hippocampus. Front. Cell. Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef]
- Caputi, V.; Giron, M.C. Microbiome-Gut-Brain Axis and Toll-Like Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepeda, M.S.; Katz, E.G.; Blacketer, C. Microbiome-Gut-Brain Axis: Probiotics and Their Association with Depression. J. Neuropsychiatry Clin. Neurosci. 2017, 29, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Kanji, S.; Fonseka, T.M.; Marshe, V.S.; Sriretnakumar, V.; Hahn, M.K.; Müller, D.J. The microbiome-gut-brain axis: Implications for schizophrenia and antipsychotic induced weight gain. Eur. Arch Psychiatry Clin. Neurosci. 2018, 268, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, A.; Carpéné, C.; Mercader, J. Resveratrol, Metabolic Syndrome, and Gut Microbiota. Nutrients 2018, 10, 1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gureev, A.P.; Syromyatnikov, M.Y.; Ignatyeva, D.A.; Valuyskikh, V.V.; Solodskikh, S.A.; Panevina, A.V.; Gryaznova, M.V.; Kokina, A.V.; Popov, V.N. Effect of long-term methylene blue treatment on the composition of mouse gut microbiome and its relationship with the cognitive abilities of mice. PLoS ONE 2020, 15, e0241784. [Google Scholar] [CrossRef]
- Storm-Larsen, C.; Myhr, K.M.; Farbu, E.; Midgard, R.; Nyquist, K.; Broch, L.; Berg-Hansen, P.; Buness, A.; Holm, K.; Ueland, T.; et al. Gut microbiota composition during a 12-week intervention with delayed-release dimethyl fumarate in multiple sclerosis—A pilot trial. Mult. Scler. J. Exp. Transl. Clin. 2019, 5, 2055217319888767. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jia, Z.; Zhu, H. Regulation of Nrf2 Signaling. React. Oxyg Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Unni, S.; Deshmukh, P.; Krishnappa, G.; Kommu, P.; Padmanabhan, B. Structural insights into the multiple binding modes of Dimethyl Fumarate (DMF) and its analogs to the Kelch domain of Keap1. FEBS J. 2021, 288, 1599–1613. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, A.; Nagarajan, R.; Sundarraj, K.; Palanisamy, K.; Perumal, E. Identification of compounds that inhibit the binding of Keap1a/Keap1b Kelch DGR domain with Nrf2 ETGE/DLG motifs in zebrafish. Basic Clin. Pharmacol. Toxicol. 2019, 125, 259–270. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Folgado, S.L.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Samarghandian, S. The therapeutic effect of resveratrol: Focusing on the Nrf2 signaling pathway. Biomed. Pharmacother 2020, 127, 110234. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, W.; Wang, Z.; Wang, Z.; Jin, X.; Xu, J.; Bai, L.; Li, Y.; Cui, J.; Cai, L. Resveratrol attenuates testicular apoptosis in type 1 diabetic mice: Role of Akt-mediated Nrf2 activation and p62-dependent Keap1 degradation. Redox Biol. 2018, 14, 609–617. [Google Scholar] [CrossRef]
- Wen, Y.; Li, W.; Poteet, E.C.; Xie, L.; Tan, C.; Yan, L.J.; Ju, X.; Liu, R.; Qian, H.; Marvin, M.A.; et al. Alternative mitochondrial electron transfer as a novel strategy for neuroprotection. J. Biol. Chem. 2011, 286, 16504–16515. [Google Scholar] [CrossRef] [Green Version]
- Atamna, H.; Atamna, W.; Al-Eyd, G.; Shanower, G.; Dhahbi, J.M. Combined activation of the energy and cellular-defense pathways may explain the potent anti-senescence activity of methylene blue. Redox Biol. 2015, 6, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Tretter, L.; Horvath, G.; Hölgyesi, A.; Essek, F.; Adam-Vizi, V. Enhanced hydrogen peroxide generation accompanies the beneficial bioenergetic effects of methylene blue in isolated brain mitochondria. Free Radic. Biol. Med. 2014, 77, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Gureev, A.P.; Shaforostova, E.A.; Laver, D.A.; Khorolskaya, V.G.; Syromyatnikov, M.Y.; Popov, V.N. Methylene blue elicits non-genotoxic H2O2 production and protects brain mitochondria from rotenone toxicity. J. Appl. Biomed. 2019, 17, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Gureev, A.P.; Syromyatnikov, M.Y.; Gorbacheva, T.M.; Starkov, A.A.; Popov, V.N. Methylene blue improves sensorimotor phenotype and decreases anxiety in parallel with activating brain mitochondria biogenesis in mid-age mice. Neurosci. Res. 2016, 113, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; McKercher, S.R.; Lipton, S.A. Reprint of: Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic. Biol. Med. 2014, 66, 45–57. [Google Scholar] [CrossRef]
- Miller, C.J.; Gounder, S.S.; Kannan, S.; Goutam, K.; Muthusamy, V.R.; Firpo, M.A.; Symons, J.D.; Paine, R., 3rd; Hoidal, J.R.; Rajasekaran, N.S. Disruption of Nrf2/ARE signaling impairs antioxidant mechanisms and promotes cell degradation pathways in aged skeletal muscle. Biochim. Biophys. Acta 2012, 1822, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, W.Y.; Shahidi, F. Antioxidant activity of resveratrol ester derivatives in food and biological model systems. Food Chem 2018, 261, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Mankowski, R.T.; Burman, J.L.; Donisi, L.; Kim, J.S.; Marzetti, E.; Leeuwenburgh, C. Mitochondrial quality control mechanisms as molecular targets in cardiac ageing. Nat. Rev. Cardiol. 2018, 15, 543–554. [Google Scholar] [CrossRef]
- Scarpulla, R.C. Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol. Rev. 2008, 88, 611–638. [Google Scholar] [CrossRef] [Green Version]
- Islam, H.; Hood, D.A.; Gurd, B.J. Looking beyond PGC-1α: Emerging regulators of exercise-induced skeletal muscle mitochondrial biogenesis and their activation by dietary compounds. Appl. Physiol. Nutr. Metab. 2020, 45, 11–23. [Google Scholar] [CrossRef]
- Kiyama, T.; Chen, C.K.; Wang, S.W.; Pan, P.; Ju, Z.; Wang, J.; Takada, S.; Klein, W.H.; Mao, C.A. Essential roles of mitochondrial biogenesis regulator Nrf1 in retinal development and homeostasis. Mol. Neurodegener. 2018, 13, 56. [Google Scholar] [CrossRef]
- Han, S.; Jeong, Y.Y.; Sheshadri, P.; Su, X.; Cai, Q. Mitophagy regulates integrity of mitochondria at synapses and is critical for synaptic maintenance. EMBO Rep. 2020, 21, e49801. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Wang, Y.; Wang, G.; Mao, L.; Zhang, D.; Wang, J. Effects of resveratrol on learning and memory in rats with vascular dementia. Mol. Med. Rep. 2019, 20, 4587–4593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabrizian, K.; Musavi, S.S.; Rigi, M.; Hosseindadi, F.; Kordi, S.; Shamshirgaran, F.; Bazi, A.; Shahraki, J.; Rezaee, R.; Hashemzaei, M. Behavioral and molecular effects of intrahippocampal infusion of auraptene, resveratrol, and curcumin on H-89-induced deficits on spatial memory acquisition and retention in Morris water maze. Hum. Exp. Toxicol. 2019, 38, 775–784. [Google Scholar] [CrossRef]
- Shen, D.; Tian, X.; Sang, W.; Song, R. Effect of Melatonin and Resveratrol against Memory Impairment and Hippocampal Damage in a Rat Model of Vascular Dementia. Neuroimmunomodulation 2016, 23, 318–331. [Google Scholar] [CrossRef]
- Xu, B.L.; Zhang, H.; Ma, L.N.; Dong, W.; Zhao, Z.W.; Zhang, J.S.; Wang, Y.L.; Zhang, X.; Wang, R. Resveratrol prevents high-calorie diet-induced learning and memory dysfunction in juvenile C57BL/6J mice. Neurol. Res. 2018, 40, 709–715. [Google Scholar] [CrossRef]
- Labban, S.; Alghamdi, B.S.; Alshehri, F.S.; Kurdi, M. Effects of melatonin and resveratrol on recognition memory and passive avoidance performance in a mouse model of Alzheimer’s disease. Behav. Brain Res. 2021, 402, 113100. [Google Scholar] [CrossRef]
- Kodali, M.; Parihar, V.K.; Hattiangady, B.; Mishra, V.; Shuai, B.; Shetty, A.K. Resveratrol prevents age-related memory and mood dysfunction with increased hippocampal neurogenesis and microvasculature, and reduced glial activation. Sci. Rep. 2015, 5, 8075. [Google Scholar] [CrossRef] [Green Version]
- Farzaei, M.H.; Rahimi, R.; Nikfar, S.; Abdollahi, M. Effect of resveratrol on cognitive and memory performance and mood: A meta-analysis of 225 patients. Pharmacol. Res. 2018, 128, 338–344. [Google Scholar] [CrossRef]
- Paraiso, H.C.; Kuo, P.C.; Curfman, E.T.; Moon, H.J.; Sweazey, R.D.; Yen, J.H.; Chang, F.L.; Yu, I.C. Dimethyl fumarate attenuates reactive microglia and long-term memory deficits following systemic immune challenge. J. Neuroinflammation 2018, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Majkutewicz, I.; Kurowska, E.; Podlacha, M.; Myślińska, D.; Grembecka, B.; Ruciński, J.; Plucińska, K.; Jerzemowska, G.; Wrona, D. Dimethyl fumarate attenuates intracerebroventricular streptozotocin-induced spatial memory impairment and hippocampal neurodegeneration in rats. Behav. Brain Res. 2016, 308, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L.; Jensen, R.A.; Vasquez, B.J.; McGuinness, T.; McGaugh, J.L. Methylene blue alters retention of inhibitory avoidance responses. Physiol. Psychol. 1978, 6, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Callaway, N.L.; Riha, P.D.; Wrubel, K.M.; McCollum, D.; Gonzalez-Lima, F. Methylene blue restores spatial memory retention impaired by an inhibitor of cytochrome oxidase in rats. Neurosci. Lett. 2002, 332, 83–86. [Google Scholar] [CrossRef]
- Rodriguez, P.; Zhou, W.; Barrett, D.W.; Altmeyer, W.; Gutierrez, J.E.; Li, J.; Lancaster, J.L.; Gonzalez-Lima, F.; Duong, T.Q. Multimodal Randomized Functional MR Imaging of the Effects of Methylene Blue in the Human Brain. Radiology 2016, 281, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gareau, M.G. Microbiota-gut-brain axis and cognitive function. Adv. Exp. Med. Biol. 2014, 817, 357–371. [Google Scholar] [CrossRef]
- Manderino, L.; Carroll, I.; Azcarate-Peril, M.A.; Rochette, A.; Heinberg, L.; Peat, C.; Steffen, K.; Mitchell, J.; Gunstad, J. Preliminary Evidence for an Association Between the Composition of the Gut Microbiome and Cognitive Function in Neurologically Healthy Older Adults. J. Int. Neuropsychol. Soc. 2017, 23, 700–705. [Google Scholar] [CrossRef]
- Wang, L.; Lu, J.; Zeng, Y.; Guo, Y.; Wu, C.; Zhao, H.; Zheng, H.; Jiao, J. Improving Alzheimer’s disease by altering gut microbiota in tree shrews with ginsenoside Rg1. FEMS Microbiol. Lett. 2020, 367, fnaa011. [Google Scholar] [CrossRef]
- Higarza, S.G.; Arboleya, S.; Arias, J.L.; Gueimonde, M.; Arias, N. Akkermansia muciniphila and environmental enrichment reverse cognitive impairment associated with high-fat high-cholesterol consumption in rats. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Wu, F.; Guo, X.; Zhang, M.; Ou, Z.; Wu, D.; Deng, L.; Lu, Z.; Zhang, J.; Deng, G.; Chen, S.; et al. An Akkermansia muciniphila subtype alleviates high-fat diet-induced metabolic disorders and inhibits the neurodegenerative process in mice. Anaerobe 2020, 61, 102138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, S.; Yang, Y.; Wang, Z.; Wang, B.; Zhang, B.; Yu, J.; Lu, W.; Pan, M.; Zhao, J.; et al. The diversity of gut microbiota in type 2 diabetes with or without cognitive impairment. Aging Clin. Exp. Res. 2021, 33, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Jeon, W.J.; Dong, H.J.; Shin, J.H.; Kim, I.Y.; Ho, H.; Oh, S.H.; Yoon, Y.M.; Choi, Y.K.; Suh, J.G.; Nam, K.H.; et al. Helicobacter apodemus sp. nov., a new Helicobacter species identified from the gastrointestinal tract of striped field mice in Korea. J. Vet. Sci. 2015, 16, 475–481. [Google Scholar] [CrossRef]
- Gu, C.; Zhou, W.; Wang, W.; Xiang, H.; Xu, H.; Liang, L.; Sui, H.; Zhan, L.; Lu, X. ZiBuPiYin recipe improves cognitive decline by regulating gut microbiota in Zucker diabetic fatty rats. Oncotarget 2017, 8, 27693–27703. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Dong, J.; Yin, Z.; Wu, Q.; Zhou, Y.; Zhou, X. Procyanidin B2 protects against d-galactose-induced mimetic aging in mice: Metabolites and microbiome analysis. Food Chem. Toxicol. 2018, 119, 141–149. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.H.; Luo, L.; Qin, L.Q.; Yin, J.; Yu, Z.; Zhang, L.; Wan, Z. Voluntary wheel running is capable of improving cognitive function only in the young but not the middle-aged male APPSwe/PS1De9 mice. Neurochem. Int. 2021, 145, 105010. [Google Scholar] [CrossRef]
- D’Amato, A.; Di Cesare Mannelli, L.; Lucarini, E.; Man, A.L.; Le Gall, G.; Branca, J.; Ghelardini, C.; Amedei, A.; Bertelli, E.; Regoli, M.; et al. Faecal microbiota transplant from aged donor mice affects spatial learning and memory via modulating hippocampal synaptic plasticity- and neurotransmission-related proteins in young recipients. Microbiome 2020, 8, 140. [Google Scholar] [CrossRef]
- Liu, J.; Yu, C.; Li, R.; Liu, K.; Jin, G.; Ge, R.; Tang, F.; Cui, S. High-altitude Tibetan fermented milk ameliorated cognitive dysfunction by modified gut microbiota in Alzheimer’s disease transgenic mice. Food Funct. 2020, 11, 5308–5319. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, L.; Ji, H.F. Alzheimer’s Disease Histological and Behavioral Manifestations in Transgenic Mice Correlate with Specific Gut Microbiome State. J. Alzheimers Dis. 2017, 56, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.A.; Ida, M.; Peterson, V.L.; Prenderville, J.A.; Moloney, G.M.; Izumo, T.; Murphy, K.; Murphy, A.; Ross, R.P.; Stanton, C.; et al. Revisiting Metchnikoff: Age-related alterations in microbiota-gut-brain axis in the mouse. Brain Behav. Immun. 2017, 65, 20–32. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.F.; Zhang, P.; Wang, H.; Zhang, Q.; Yu, S.; Yu, Y. Chronic rhein treatment improves recognition memory in high-fat diet-induced obese male mice. J. Nutr. Biochem. 2016, 36, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Leigh, S.J.; Kaakoush, N.O.; Westbrook, R.F.; Morris, M.J. Minocycline-induced microbiome alterations predict cafeteria diet-induced spatial recognition memory impairments in rats. Transl. Psychiatry 2020, 10, 92. [Google Scholar] [CrossRef]
- Beilharz, J.E.; Kaakoush, N.O.; Maniam, J.; Morris, M.J. Cafeteria diet and probiotic therapy: Cross talk among memory, neuroplasticity, serotonin receptors and gut microbiota in the rat. Mol. Psychiatry 2018, 23, 351–361. [Google Scholar] [CrossRef]
- Li, W.; Zhao, Q.; Wang, J.; Wang, Y.; Wen, T. Dcf1 deletion presents alterations in gut microbiota of mice similar to Parkinson’s disease. Biochem. Biophys. Res. Commun. 2020, 529, 1137–1144. [Google Scholar] [CrossRef]
- Sun, Y.; Tang, Y.; Xu, X.; Hu, K.; Zhang, Z.; Zhang, Y.; Yi, Z.; Zhu, Q.; Xu, R.; Zhang, Y.; et al. Lead exposure results in defective behavior as well as alteration of gut microbiota composition in flies and their offsprings. Int. J. Dev. Neurosci. 2020, 80, 699–708. [Google Scholar] [CrossRef]
- Leigh, S.J.; Kaakoush, N.O.; Bertoldo, M.J.; Westbrook, R.F.; Morris, M.J. Intermittent cafeteria diet identifies fecal microbiome changes as a predictor of spatial recognition memory impairment in female rats. Transl. Psychiatry 2020, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, T.; Harvey, B.H.; Harden, L.M.; Laburn, H.P.; Mitchell, D. Simulated systemic recurrent Mycoplasma infection in rats induces recurrent sickness responses without residual impairment in spatial learning and memory. Physiol. Behav. 2012, 105, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, T.; Sabbar, M.; Baartman, T.L.; Laburn, H.P.; Mitchell, D.; Dukhan, T.; Harden, L.M. Simulated acute central Mycoplasma infections in rats induce fever, anorexia, body mass stunting and lethargy but spare memory. Physiol. Behav. 2016, 163, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, T.; Harvey, B.H.; Harden, L.M.; Laburn, H.P.; Mitchell, D. Dissociation between learning and memory impairment and other sickness behaviours during simulated Mycoplasma infection in rats. Brain Behav. Immun. 2011, 25, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Le Bastard, Q.; Al-Ghalith, G.A.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic review: Human gut dysbiosis induced by non-antibiotic prescription medications. Aliment. Pharmacol. Ther. 2018, 47, 332–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Wan, Y.D.; Pan, X.T.; Zhou, C.Y.; Lin, N.; Ma, C.T.; Yao, J.; Su, Z.; Wan, C.; Yu, Y.W.; et al. Association Between Gut Bacterial Diversity and Mortality in Septic Shock Patients: A Cohort Study. Med. Sci. Monit. 2019, 25, 7376–7382. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drugs FDA: FDA-Approved Drugs. New Drug Application (NDA): 204630. Available online: https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm?event=overview.process&varApplNo=204630 (accessed on 29 April 2021).
- Danilova, N.A.; Abdulkhakov, S.R.; Grigoryeva, T.V.; Markelova, M.I.; Vasilyev, I.Y.; Boulygina, E.A.; Ardatskaya, M.D.; Pavlenko, A.V.; Tyakht, A.V.; Odintsova, A.K.; et al. Markers of dysbiosis in patients with ulcerative colitis and Crohn’s disease. Ter. Arkh. 2019, 91, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.J.; Lee, J.; Shin, N.R.; Kim, M.S.; Hyun, D.W.; Yun, J.H.; Kim, P.S.; Whon, T.W.; Bae, J.W. Chronic Repression of mTOR Complex 2 Induces Changes in the Gut Microbiota of Diet-induced Obese Mice. Sci. Rep. 2016, 6, 30887. [Google Scholar] [CrossRef] [Green Version]
- Gureev, A.P.; Shaforostova, E.A.; Starkov, A.A.; Popov, V.N. Simplified qPCR method for detecting excessive mtDNA damage induced by exogenous factors. Toxicology 2017, 382, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.W.; Chen, M.K.; Yang, B.Y.; Huang, X.J.; Zhang, X.R.; He, L.Q.; Zhang, J.; Hua, Z.C. Use of 16S rRNA Gene-Targeted Group-Specific Primers for Real-Time PCR Analysis of Predominant Bacteria in Mouse Feces. Appl. Environ. Microbiol. 2015, 81, 6749–6756. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Flyvbjerg, H. Error filtering, pair assembly and error correction for next-generation sequencing reads. Bioinformatics 2015, 31, 3476–3482. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Open Field | |||||
|---|---|---|---|---|---|---|
| Indicator | Horizontal Activity (s) | Time in the Center (s) | Entering in the Center (Number) | Rearing (Number) | Hole-Poking (Number) | Grooming (s) |
| Control | 114.62 ± 14.49 | 19.87 ± 7.13 | 5.87 ± 1.78 | 15.25 ± 2.25 | 4.5 ± 0.91 | 10.62 ± 1.44 |
| DMF | 124.00 ± 7.41 | 16.00 ± 4.07 | 5.12 ± 0.67 | 12.75 ± 1.23 | 3.12 ± 0.69 | 9.25 ± 2.61 |
| MB | 83.67 ± 14.27 | 10.5 ± 2.66 | 4.83 ± 1.51 | 14.00 ± 2.67 | 2.66 ± 0.80 | 9.00 ± 3.38 |
| RSV | 90.75 ± 12.89 | 10.66 ± 3.38 | 4.5 ± 1.18 | 7.62 ± 2.09 | 5.5 ± 2.38 | 24.25 ± 4.38* |
| Test | Open Field | Dark-Light Box | EPM | String | ||
| Indicator | Grooming (Number) | Defecation | Time in the Open Compartment (s) | Transition between Compartments (Number) | Time in the Open Arm (s) | Scores (Number) |
| Control | 1.75 ± 0.16 | 1.62 ± 0.86 | 235.25 ± 8.64 | 15.75 ± 2.68 | 4.37 ± 1.61 | 2.8 ± 0.33 |
| DMF | 4.12 ± 2.42 | 0.25 ± 0.25 | 253.22 ± 9.16 | 7.67 ± 1.43 * | 8.44 ± 2.53 | 1.82 ± 0.39 |
| MB | 1.5 ± 0.22 | 1.5 ± 0.5 | 231.83 ± 7.50 | 16.83 ± 1.42 | 6.83 ± 2.09 | 3.5 ± 0.43 |
| RSV | 2.75 ± 0.65 | 2.5 ± 1.44 | 64.37 ± 12.64 *** | 8.37 ± 2.14 | 5.75 ± 4.34 | 3.2 ± 0.40 |
| Acquisition | Learning | Probe | ||||
|---|---|---|---|---|---|---|
| Start SW | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 |
| N; E; SE; NW | SE; N; NW; E | NW; SE; E; N | E; NW; N; SE | N; SE; E; NW | NW | |
| Control | 2611 ± 514 | 1852 ± 306 | 1511 ± 266 | 1956 ± 313 | 1265 ± 232 | 1401 ± 410 |
| DMF | 2512 ± 230 | 2018 ± 151 | 2130 ± 201 | 2035 ± 187 | 1887 ± 222 | 2897 ± 359 |
| MB | 2573 ± 266 | 1960 ± 257 | 2174 ± 276 | 1134 ± 192 | 1654 ± 295 | 1682 ± 464 |
| RSV | 1896 ± 238 | 1894 ± 220 | 1724 ± 236 | 1461 ± 195 | 1544 ± 223 | 1376 ± 385 |
| Reversal | Learning | Probe | ||||
| Start NE | Day 7 | Day 8 | Day 9 | Day 10 | Day 11 | Day 12 |
| S; W; NW; SE | NW; S; SE; W | SE; NW; W; S | W; SE; S; NW | S; NW; W; SE | SE | |
| Control | 2774 ± 423 | 2306 ± 398 | 2726 ± 420 | 2212 ± 213 | 2064 ± 498 | 1324 ± 443 |
| DMF | 1228 ± 184 ** | 1491 ± 195 | 996 ± 174 *** | 1122 ± 153 ** | 1142 ± 154 * | 1062 ± 401 |
| MB | 1113 ± 217 ** | 920 ± 188 ** | 1335 ± 266* | 1195 ± 179 * | 682 ± 116 *** | 1838 ± 334 |
| RSV | 1430 ± 195 * | 1107 ± 165 * | 1229 ± 201 ** | 1160 ± 163 ** | 1076 ± 168 * | 1691 ± 338 |
| Acquisition | Learning | Probe | ||||
|---|---|---|---|---|---|---|
| Start SW | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 |
| N; E; SE; NW | SE; N; NW; E | NW; SE; E; N | E; NW; N; SE | N; SE; E; NW | NW | |
| Control | 38.3 ± 6.2 | 28.4 ± 5.4 | 35.5 ± 10.0 | 29.5 ± 4.5 | 34.5 ± 6.5 | 24.5 ± 8.5 |
| DMF | 47.3 ± 3.3 | 49.9 ± 2.6 | 46.7 ± 3.4 | 44.2 ± 3.3 | 36.9 ± 3.8 | 49.2 ± 5.6 |
| MB | 45.9 ± 4.0 | 34.3 ± 4.6 | 37.3 ± 4.5 | 24.3 ± 4.1 | 30.7 ± 5.6 | 29.3 ± 9.7 |
| RSV | 41.2 ± 4.3 | 37.6 ± 3.8 | 31.6 ± 4.3 | 29.2 ± 3.9 | 24.5 ± 3.6 | 29.4 ± 8.9 |
| Reversal | Learning | Probe | ||||
| Start NE | Day 7 | Day 8 | Day 9 | Day 10 | Day 11 | Day 12 |
| S; W; NW; SE | NW; S; SE; W | SE; NW; W; S | W; SE; S; NW | S; NW; W; SE | SE | |
| Control | 19.7 ± 3.6 | 17.6 ± 3.5 | 20.9 ± 3.3 | 21.5 ± 2.6 | 19.8 ± 5.5 | 22.1 ± 7.4 |
| DMF | 21.7 ± 3.3 | 31.7 ± 3.9 | 17.4 ± 3.2 | 21.6 ± 3.4 | 23.0 ± 3.5 | 16.8 ± 6.6 |
| MB | 17.7 ± 3.1 | 15.1 ± 3.0 | 19.1 ± 3.8 | 18.4 ± 2.8 | 13.1 ± 2.5 | 31.0 ± 3.9 |
| RSV | 26 ± 3.8 | 22.4 ± 3.6 | 26.1 ± 4.3 | 23.1 ± 3.8 | 25.9 ± 3.7 | 33.0 ± 8.6 |
| Acquisition | Learning | Probe | ||||
|---|---|---|---|---|---|---|
| Start SW | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 |
| N; E; SE; NW | SE; N; NW; E | NW; SE; E; N | E; NW; N; SE | N; SE; E; NW | NW | |
| Control | 10.3 ± 1.62 | 9.3 ± 1.38 | 11.4 ± 2.95 | 7.5 ± 0.98 | 5.9 ± 0.87 | 8.5 ± 3.09 |
| DMF | 11 ± 0.99 | 17.4 ± 1.52 | 10.9 ± 0.98 | 14.2 ± 1.55 | 10.8 ± 1.30 | 10.6 ± 1.83 |
| MB | 11.8 ± 1.72 | 13.9 ± 2.29 | 12.2 ± 1.42 | 13.9 ± 2.68 | 10.1 ± 1.81 | 7.8 ± 2.40 |
| RSV | 9.7 ± 0.97 | 12.7 ± 1.66 | 10.9 ± 1.61 | 9.5 ± 1.47 | 8.3 ± 1.27 | 10.9 ± 3.79 |
| Reversal | Learning | Probe | ||||
| Start NE | Day 7 | Day 8 | Day 9 | Day 10 | Day 11 | Day 12 |
| S; W; NW; SE | NW; S; SE; W | SE; NW; W; S | W; SE; S; NW | S; NW; W; SE | SE | |
| Control | 5.7 ± 0.90 | 5.2 ± 0.43 | 7.2 ± 0.94 | 6.5 ± 0.51 | 8.3 ± 1.23 | 4.6 ± 1.05 |
| DMF | 7.9 ± 1.14 | 7.6 ± 0.95 | 5.7 ± 0.79 | 4.9 ± 0.79 | 6.0 ± 0.80 | 5.6 ± 1.83 |
| MB | 6.3 ± 1.08 | 5.3 ± 0.86 | 5.3 ± 0.76 | 5.8 ± 0.84 | 3.9 ± 0.56 | 10.7 ± 2.16 |
| RSV | 7.6 ± 1.18 | 6.4 ± 1.19 | 5.2 ± 1.01 | 5.4 ± 1.15 | 5.1 ± 0.73 | 9.5 ± 3.37 |
| Hippocampus | ||||
| Control | DMF | MB | RSV | |
| 1 fragment | 2.05 ± 0.30 | 2.97 ± 0.30 | 1.20 ± 0.34 | 3.68 ± 0.19 *** |
| 2 fragment | 2.33 ± 0.24 | 3.83 ± 0.71 | 0.80 ± 0.34 ** | 4.63 ± 0.26 *** |
| 3 fragment | 2.19 ± 0.54 | 3.46 ± 0.21 * | 1.50 ± 0.48 | 3.29 ± 0.28 |
| 4 fragment | 3.84 ± 0.28 | 4.03 ± 0.20 | 1.72 ± 0.61 ** | 3.82 ± 0.27 |
| 5 fragment | 3.84 ± 0.23 | 4.09 ± 0.18 | 1.39 ± 0.32 *** | 4.22 ± 0.24 |
| 6 fragment | 4.75 ± 0.30 | 5.17 ± 0.17 | 1.82 ± 0.59 *** | 4.39 ± 0.21 |
| Forebrain | ||||
| 1 fragment | 2.26 ± 0.35 | 1.86 ± 0.47 | 3.16 ± 0.35 | 3.03 ± 0.42 |
| 2 fragment | 5.35 ± 0.34 | 3.60 ± 0.79 * | 6.23 ± 0.24 | 5.51 ± 0.59 |
| 3 fragment | 4.74 ± 0.14 | 2.94 ± 0.51 *** | 4.72 ± 0.16 | 4.32 ± 0.44 |
| 4 fragment | 1.71 ± 0.47 | 0.47 ± 0.69 | 1.26 ± 0.53 | 1.23 ± 0.48 |
| 5 fragment | 2.50 ± 0.74 | 0.97 ± 0.67 | 3.00 ± 0.62 | 3.17 ± 0.53 |
| 6 fragment | 5.92 ± 0.45 | 3.59 ± 1.05 * | 1.55 ± 1.24 ** | 1.47 ± 0.98 *** |
| Mid-Brain | ||||
| 1 fragment | 2.30 ± 0.37 | 2.90 ± 0.29 | 3.35 ± 0.35 * | 3.16 ± 0.46 |
| 2 fragment | 7.54 ± 0 | 2.27 ± 0.72 *** | 2.48 ± 0.88 *** | 2.43 ± 0.88 *** |
| 3 fragment | 4.15 ± 0.45 | 2.88 ± 0.55 | 2.98 ± 0.59 | 2.62 ± 0.52 * |
| 4 fragment | 2.71 ± 0.38 | 3.45 ± 0.29 | 3.14 ± 0.17 | 3.39 ± 0.24 |
| 5 fragment | 3.1 ± 0.53 | 2.71 ± 0.57 | 3.60 ± 0.35 | 2.74 ± 0.45 |
| 6 fragment | 3.1 ± 0.65 | 4.28 ± 0.59 | 5.05 ± 0.45 * | 5.84 ± 0.48 ** |
| Control | DMF | MB | RSV | |
|---|---|---|---|---|
| Bacteroidetes | 85.20 ± 2.74 | 84.19 ± 2.31 | 75.44 ± 4.31 | 79.70 ± 3.01 |
| Firmicutes | 9.45 ± 1.76 | 9.48 ± 2.07 | 18.18 ± 2.97 | 17.60 ± 2.86 |
| Actinobacteria | 0.26 ± 0.04 | 0.25 ± 0.08 | 0.26 ± 0.08 | 0.11 ± 0.03 |
| Betaproteobacteria | 2.24 ± 1.05 | 1.20 ± 0.36 | 0.90 ± 0.24 | 0.75 ± 0.17 |
| Epsilonproteobacteria | 0.46 ± 0.29 | 0.30 ± 0.07 | 0.20 ± 0.07 | 0.60 ± 0.21 |
| Delta- and Gammaproteobacteria | 1.26 ± 0.30 | 2.24 ± 0.59 | 2.78 ± 0.69 | 2.05 ± 0.90 |
| «Candidatus Saccharibacteria» | 0.42 ± 0.12 | 0.80 ± 0.29 | 0.55 ± 0.15 | 0.47 ± 0.11 |
| Deferribacteres | 0.29 ± 0.11 | 0.69 ± 0.36 | 1.60 ± 0.88 | 0.52 ± 0.23 |
| Tenericutes | 0.00 ± 0.00 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.00 ± 0.00 |
| Verrucomicrobia | 0.41 ± 0.15 | 0.82 ± 0.67 | 0.06 ± 0.01 | 0.17 ± 0.04 |
| Phylum | Class | Family | Genus | Control | DMF | MB | RSV |
|---|---|---|---|---|---|---|---|
| Abditibacteriota | Abditibacteriaceae | Abditibacteriaceae | Abditibacterium | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Actinobacteriota | Actinobacteria | Propionibacteriaceae | Cutibacterium | 0.022 ± 0.022 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 |
| Bifidobacteriaceae | Bifidobacterium | 0.005 ± 0.001 | 0.004 ± 0.003 | 0.003 ± 0.001 | 0.013 ± 0.007 | ||
| Corynebacteriaceae | Corynebacterium | 0.002 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | ||
| Coriobacteriia | Atopobiaceae | Olsenella | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | |
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Methylobacterium-Methylorubrum | 0.005 ± 0.005 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Caulobacterales | Asticcacaulis | 0.002 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | ||
| Gammaproteobacteria | Burkholderiales | Parasutterella | 0.085 ± 0.025 | 0.082 ± 0.055 | 0.054 ± 0.013 | 0.088 ± 0.021 | |
| Burkholderia-Caballeronia-Paraburkholderia | 0.006 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | |||
| Enterobacteriaceae | Escherichia-Shigella | 0.002 ± 0.002 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | ||
| Xanthomonadaceae | Pseudoxanthomonas | 0.004 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | ||
| Moraxellaceae | Acinetobacter | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | ||
| Rhodanobacteraceae | Rudaea | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | ||
| Bacteroidota | Bacteroidia | Prevotellaceae | Prevotella | 0.135 ± 0.031 | 0.135 ± 0.058 | 0.096 ± 0.018 | 0.160 ± 0.027 |
| Prevotellaceae UCG-001 | 0.023 ± 0.005 | 0.031 ± 0.014 | 0.014 ± 0.004 | 0.032 ± 0.008 | |||
| Prevotellaceae Ga6A1 group | 0.029 ± 0.008 | 0.015 ± 0.009 | 0.030 ± 0.011 | 0.023 ± 0.010 | |||
| Alloprevotella | 0.022 ± 0.007 | 0.022 ± 0.021 | 0.010 ± 0.002 | 0.012 ± 0.003 | |||
| Marinifilaceae | Odoribacter | 0.026 ± 0.006 | 0.028 ± 0.013 | 0.014 ± 0.004 | 0.025 ± 0.004 | ||
| Bacteroidaceae | Bacteroides | 0.074 ± 0.025 | 0.027 ± 0.009 | 0.018 ± 0.003 | 0.029 ± 0.004 | ||
| Muribaculaceae | Muribaculum | 0.033 ± 0.006 | 0.020 ± 0.010 | 0.017 ± 0.004 | 0.038 ± 0.013 | ||
| Tannerellaceae | Parabacteroides | 0.008 ± 0.002 | 0.005 ± 0.003 | 0.004 ± 0.001 | 0.005 ± 0.001 | ||
| Rikenellaceae | Rikenella | 0.006 ± 0.001 | 0.006 ± 0.004 | 0.007 ± 0.001 | 0.006 ± 0.001 | ||
| Alistipes | 0.025 ± 0.005 | 0.019 ± 0.011 | 0.011 ± 0.003 | 0.023 ± 0.006 | |||
| Rikenellaceae RC9 gut group | 0.035 ± 0.004 | 0.052 ± 0.029 | 0.050 ± 0.027 | 0.067 ± 0.025 | |||
| Campilobacterota | Campylobacteria | Helicobacteraceae | Helicobacter | 0.038 ± 0.007 | 0.095 ± 0.070 | 0.058 ± 0.021 | 0.050 ± 0.007 |
| Firmicutes | Clostridia | Lachnospiraceae | Lachnospiraceae UCG-001 | 0.080 ± 0.025 | 0.037 ± 0.017 | 0.147 ± 0.062 | 0.034 ± 0.014 |
| Herbinix | 0.014 ± 0.004 | 0.012 ± 0.016 | 0.021 ± 0.015 | 0.007 ± 0.001 | |||
| [Ruminococcus] gnavus group | 0.013 ± 0.002 | 0.029 ± 0.022 | 0.023 ± 0.008 | 0.016 ± 0.005 | |||
| Acetatifactor | 0.006 ± 0.001 | 0.010 ± 0.014 | 0.034 ± 0.019 | 0.031 ± 0.012 | |||
| Stomatobaculum | 0.002 ± 0.001 | 0.004 ± 0.002 | 0.003 ± 0.001 | 0.008 ±0.003 | |||
| Lachnospiraceae NK4A136 group | 0.027 ± 0.005 | 0.031 ± 0.020 | 0.036 ± 0.006 | 0.052 ± 0.011 | |||
| Tuzzerella | 0.004 ± 0.001 | 0.006 ± 0.006 | 0.002 ± 0.000 | 0.002 ± 0.001 | |||
| Tyzzerella | 0.003 ± 0.001 | 0.003 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Roseburia | 0.008 ± 0.006 | 0.003 ± 0.002 | 0.003 ± 0.001 | 0.007 ± 0.004 | |||
| GCA-900066575 | 0.003 ± 0.001 | 0.002 ± 0.001 | 0.001 ± 0.000 | 0.002 ± 0.001 | |||
| ASF356 | 0.002 ± 0.000 | 0.002 ± 0.002 | 0.002 ± 0.000 | 0.001 ± 0.000 | |||
| Blautia | 0.001 ± 0.000 | 0.002 ± 0.002 | 0.001 ± 0.000 | 0.001 ± 0.001 | |||
| [Eubacterium] hallii group | 0.001 ± 0.000 | 0.002 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| A2 | 0.002 ± 0.000 | 0.002 ± 0.001 | 0.004 ± 0.001 | 0.002 ± 0.001 | |||
| Lachnospiraceae UCG-003 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.003 ± 0.001 | 0.001 ± 0.000 | |||
| [Eubacterium] xylanophilum group | 0.003 ± 0.002 | 0.003 ± 0.002 | 0.002 ± 0.000 | 0.003 ± 0.001 | |||
| [Eubacterium] fissicatena group | 0.002 ± 0.000 | 0.002 ± 0.001 | 0.003 ± 0.001 | 0.002 ± 0.000 | |||
| Mobilitalea | 0.001 ± 0.000 | 0.003 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Lachnoanaerobaculum | 0.004 ± 0.001 | 0.007 ± 0.006 | 0.005 ± 0.002 | 0.007 ± 0.001 | |||
| Oscillospiraceae | Oscillibacter | 0.014 ± 0.003 | 0.018 ± 0.013 | 0.007 ± 0.001 | 0.011 ± 0.001 | ||
| Colidextribacter | 0.024 ± 0.005 | 0.030 ± 0.012 | 0.016 ± 0.001 | 0.029 ± 0.002 | |||
| UCG-003 | 0.004 ± 0.001 | 0.009 ± 0.005 | 0.003 ± 0.001 | 0.007 ± 0.001 | |||
| Flavonifractor | 0.003 ± 0.001 | 0.003 ± 0.002 | 0.002 ± 0.000 | 0.003 ± 0.001 | |||
| UCG-002 | 0.002 ± 0.001 | 0.002 ± 0.001 | 0.000 ± 0.000 | 0.001 ± 0.000 | |||
| Intestinimonas | 0.009 ± 0.002 | 0.010 ± 0.007 | 0.006 ± 0.001 | 0.009 ± 0.002 | |||
| Ruminococcaceae | Ruminococcus | 0.018 ± 0.004 | 0.019 ± 0.015 | 0.013 ± 0.004 | 0.011 ± 0.005 | ||
| Incertae Sedis | 0.000 ± 0.001 | 0.003 ± 0.002 | 0.004 ± 0.001 | 0.003 ± 0.002 | |||
| Negativibacillus | 0.004 ± 0.001 | 0.002 ± 0.001 | 0.002 ± 0.000 | 0.003 ± 0.001 | |||
| Angelakisella | 0.002 ± 0.001 | 0.003 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Paludicola | 0.002 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Anaerotruncus | 0.004 ± 0.001 | 0.004 ± 0.002 | 0.004 ± 0.001 | 0.001 ± 0.000 | |||
| [Eubacterium] siraeum group | 0.003 ± 0.001 | 0.002 ± 0.001 | 0.002 ± 0.001 | 0.008 ± 0.004 | |||
| Clostridiaceae | Clostridium sensu stricto 1 | 0.001 ± 0.000 | 0.002 ± 0.001 | 0.003 ± 0.001 | 0.001 ± 0.000 | ||
| Anaerovoracaceae | [Eubacterium] nodatum group | 0.002 ± 0.001 | 0.002 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | ||
| Family XIII UCG-001 | 0.001 ± 0.000 | 0.003 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Monoglobaceae | Monoglobus | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.003 ± 0.002 | 0.002 ± 0.002 | ||
| Christensenellaceae | Christensenellaceae R-7 group | 0.001 ± 0.000 | 0.001 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | ||
| Peptococcaceae | Peptococcus | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.001 | 0.000 ± 0.000 | ||
| Bacilli | Erysipelotrichaceae | Allobaculum | 0.077 ± 0.020 | 0.070 ± 0.069 | 0.200 ± 0.079 | 0.067 ± 0.017 | |
| Ileibacterium | 0.017 ± 0.006 | 0.019 ± 0.030 | 0.011 ± 0.004 | 0.038 ± 0.016 | |||
| Dubosiella | 0.003 ± 0.002 | 0.004 ± 0.002 | 0.004 ± 0.002 | 0.007 ± 0.002 | |||
| Faecalibaculum | 0.001 ± 0.000 | 0.004 ± 0.003 | 0.001 ± 0.000 | 0.002 ± 0.001 | |||
| Lactobacillaceae | Lactobacillus | 0.012 ± 0.005 | 0.008 ± 0.002 | 0.015 ± 0.003 | 0.008 ± 0.003 | ||
| Atopostipes | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Streptococcaceae | Streptococcus | 0.004 ± 0.003 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.003 ± 0.001 | ||
| Acholeplasmataceae | Anaeroplasma | 0.001 ± 0.000 | 0.002 ± 0.003 | 0.003 ± 0.003 | 0.001 ± 0.000 | ||
| Staphylococcaceae | Staphylococcus | 0.002 ± 0.002 | 0.000 ± 0.000 | 0.000± 0.000 | 0.000 ± 0.000 | ||
| Jeotgalicoccus | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.003 ± 0.002 | 0.001 ± 0.000 | |||
| Exiguobacteraceae | Exiguobacterium | 0.004 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | ||
| Erysipelatoclostridiaceae | Candidatus Stoquefichus | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.004 ± 0.003 | ||
| Erysipelatoclostridium | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | |||
| Streptococcaceae | Lactococcus | 0.001 ± 0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.002 ± 0.001 | ||
| Mycoplasmataceae | Ureaplasma | 0.000 ± 0.000 | 0.002 ± 0.002 | 0.001 ± 0.000 | 0.002 ± 0.001 | ||
| Mycoplasma | 0.002 ± 0.000 | 0.001± 0.000 | 0.000 ± 0.000 | 0.001 ± 0.000 | |||
| Negativicutes | Acidaminococcaceae | Phascolarctobacterium | 0.034 ± 0.021 | 0.019 ± 0.035 | 0.002 ± 0.001 | 0.001 ± 0.000 | |
| Sporomusaceae | Pelosinus | 0.001 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | ||
| Deferribacterota | Deferribacteres | Deferribacteraceae | Mucispirillum | 0.010 ± 0.003 | 0.038 ± 0.052 | 0.014 ± 0.011 | 0.006 ± 0.001 |
| Desulfobacterota | Desulfovibrionia | Desulfovibrionaceae | Desulfovibrio | 0.001 ± 0.000 | 0.002 ± 0.002 | 0.001 ± 0.001 | 0.001 ± 0.000 |
| Bilophila | 0.001 ± 0.000 | 0.002 ± 0.001 | 0.001 ± 0.000 | 0.001 ± 0.000 | |||
| Spirochaetota | Leptospirae | Leptospiraceae | Leptospira | 0.002 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Brachyspirae | Brachyspiraceae | Brachyspira | 0.004 ± 0.001 | 0.025 ± 0.040 | 0.004 ± 0.003 | 0.002 ± 0.001 | |
| Patescibacteria | Saccharimonadia | Saccharimonadaceae | Candidatus Saccharimonas | 0.005 ± 0.001 | 0.008 ± 0.005 | 0.003 ± 0.001 | 0.008 ± 0.001 |
| Verrucomicrobiota | Verrucomicrobiae | Akkermansiaceae | Akkermansia | 0.012 ± 0.006 | 0.009 ± 0.013 | 0.010 ± 0.006 | 0.023 ± 0.014 |
| Gene | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|
| 18s | CGGCTACCACATCCAAGGAA | GCTGGAATTACTGTGGCT |
| Gapdh | GGCTCCCTAGGCCCCTCCTG | TCCCAACTCGGCCCCCAACA |
| Ppargc1a | ATGTGTCGCCTTCTTGCTCT | CACGACCTGTGTCGAGAAAA |
| Sirt1 | CTGTTTCCTGTGGGATACCTGACT | ATCGAACATGGCTTGAGGATCT |
| FoxO1 | GGGTCTGTCTCCCTTTCCTC | TCAGTGGCATTCAGCAGGTA |
| Nfe2l2 | CTCTCTGAACTCCTGGACGG | GGGTCTCCGTAAATGGAAG |
| Nrf1 | AGCACGGAGTGACCCAAA | TGTACGTGGCTACATGGACCT |
| P62 | GCCAGAGGAACAGATGGAGT | TCCGATTCTGGCATCTGTAG |
| Pink1 | GAGCAGACTCCCAGTTCTCG | GTCCCACTCCACAAGGATGT |
| Gclc | GCAGCTTTGGGTCGCAAGTAG | TGGGTCTCTTCCCAGCTCAGT |
| Gpx | AGTCCACCGTGTATGCCTTCT | GAGACGCGACATTCTCAATGA |
| Txnr2 | GATCCGGTGGCCTAGCTTG | TCGGGGAGAAGGTTCCACAT |
| Prdx5 | GGCTGTTCTAAGACCCACCTG | GGAGCCGAACCTTGCCTTC |
| Sod2 | CAGACCTGCCTTACGACTATGG | CTCGGTGGCGTTGAGATTGTT |
| Hmox1 | CACGCATATACCCGCTACCT | CCAGAGTGTTCATTCGAGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadovnikova, I.S.; Gureev, A.P.; Ignatyeva, D.A.; Gryaznova, M.V.; Chernyshova, E.V.; Krutskikh, E.P.; Novikova, A.G.; Popov, V.N. Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome. Pharmaceuticals 2021, 14, 607. https://doi.org/10.3390/ph14070607
Sadovnikova IS, Gureev AP, Ignatyeva DA, Gryaznova MV, Chernyshova EV, Krutskikh EP, Novikova AG, Popov VN. Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome. Pharmaceuticals. 2021; 14(7):607. https://doi.org/10.3390/ph14070607
Chicago/Turabian StyleSadovnikova, Irina S., Artem P. Gureev, Daria A. Ignatyeva, Maria V. Gryaznova, Ekaterina V. Chernyshova, Ekaterina P. Krutskikh, Anastasia G. Novikova, and Vasily N. Popov. 2021. "Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome" Pharmaceuticals 14, no. 7: 607. https://doi.org/10.3390/ph14070607
APA StyleSadovnikova, I. S., Gureev, A. P., Ignatyeva, D. A., Gryaznova, M. V., Chernyshova, E. V., Krutskikh, E. P., Novikova, A. G., & Popov, V. N. (2021). Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome. Pharmaceuticals, 14(7), 607. https://doi.org/10.3390/ph14070607