

Synthesis and Biological Activities of Dehydrodiisoeugenol: A Review
 , and
, and
Abstract
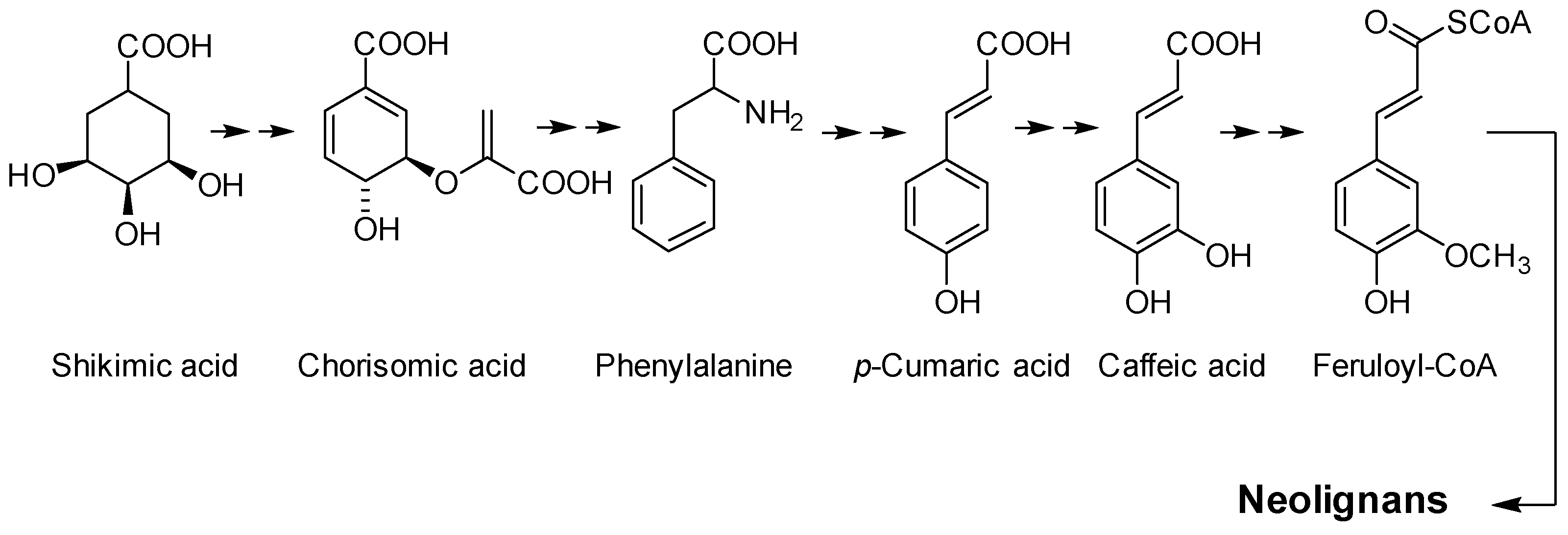
:1. Introduction
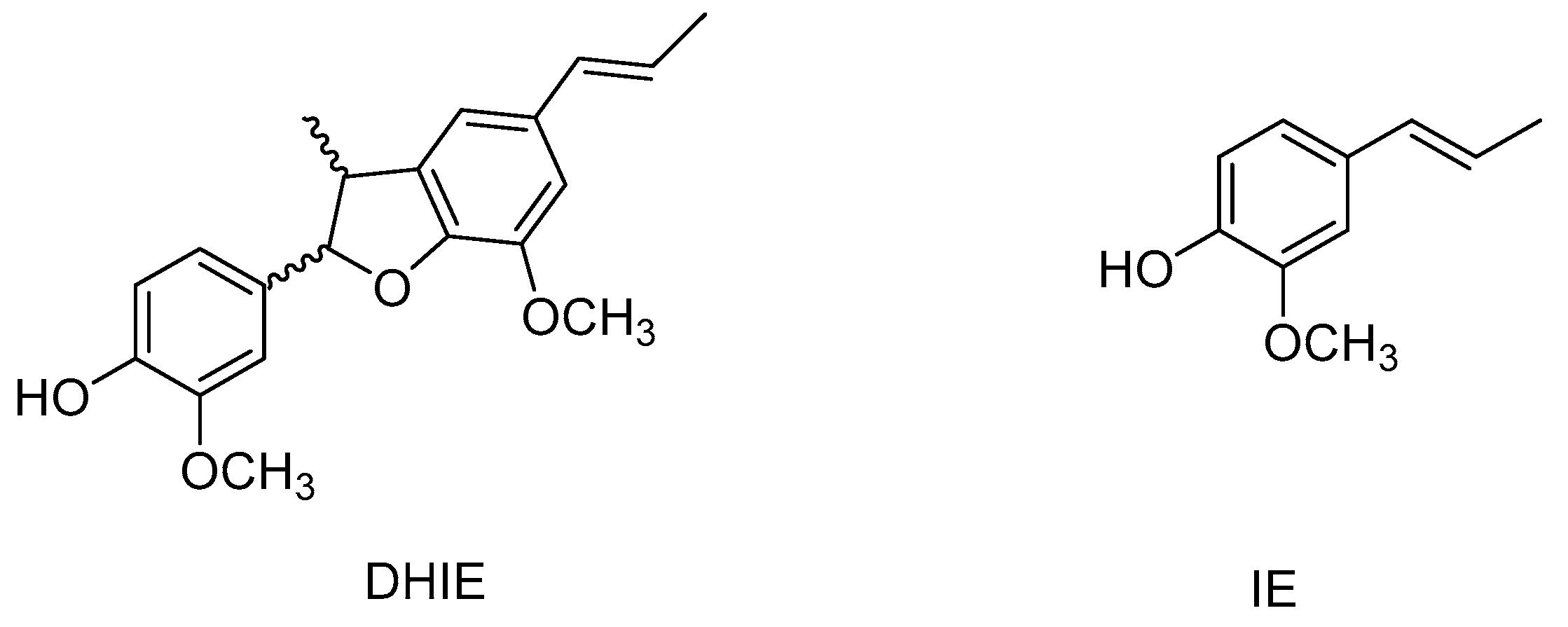
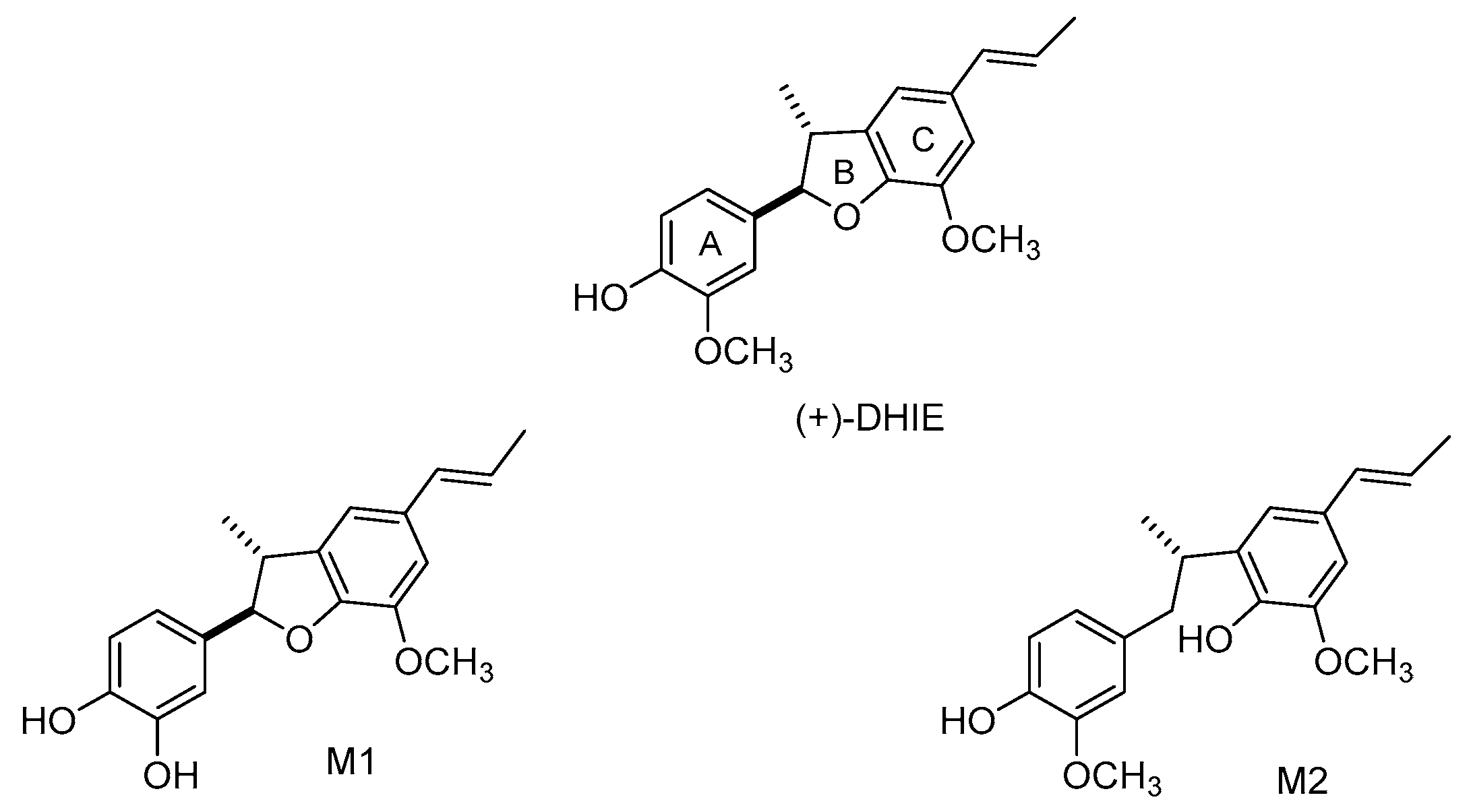
2. Characteristics of DHIE
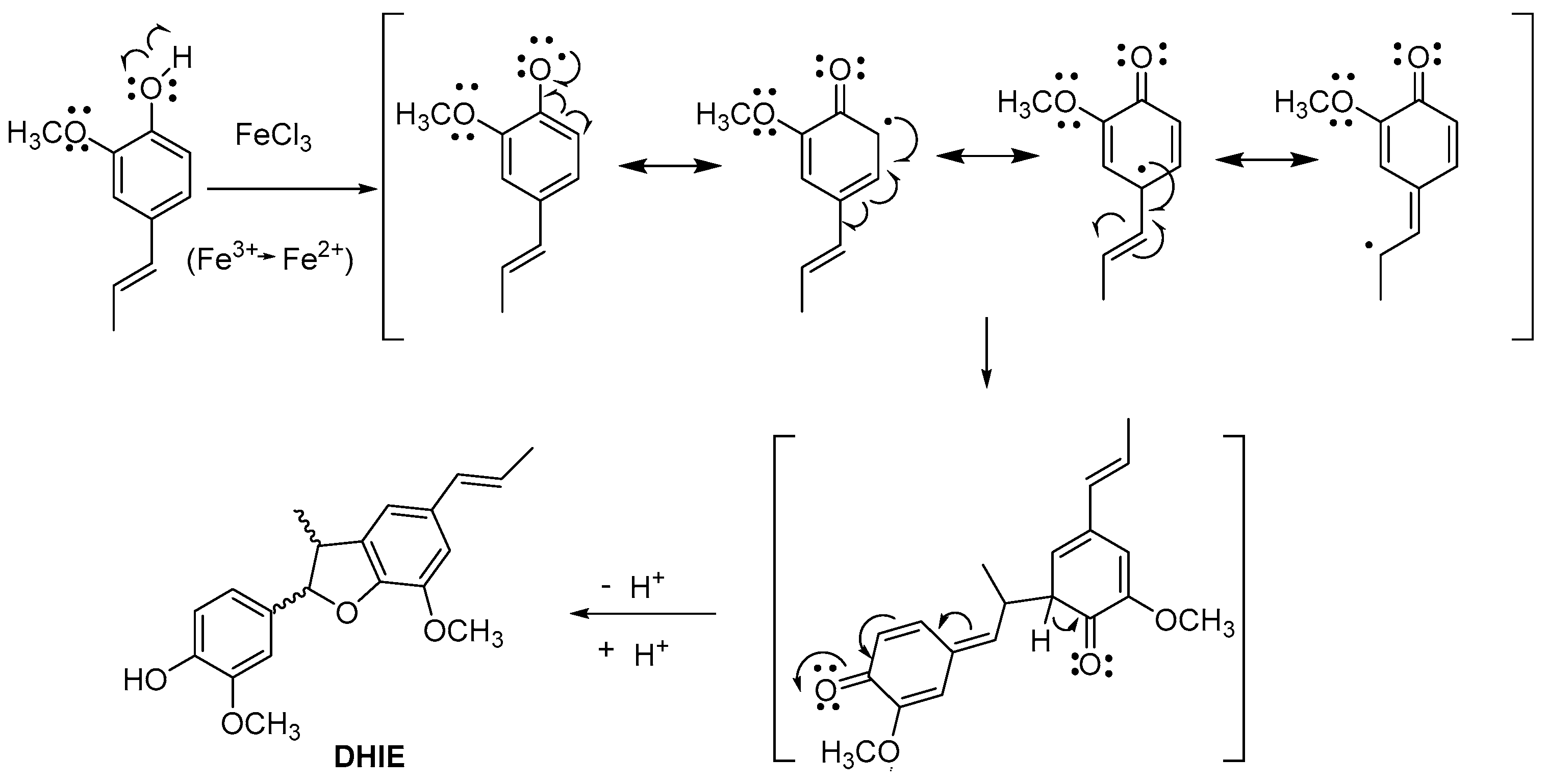
3. Classical Synthesis of DHIE
4. Biocatalytic Synthesis of DHIE
4.1. Horseradish Peroxidase for Synthesis of DHIE
4.2. Peroxidase of Cocos nucifera L. for Synthesis of DHIE
4.3. Laccase for Synthesis of DHIE
4.4. Cell Culture of Plants for Synthesis of DHIE from IE
4.5. Microorganisms for Synthesis of DHIE
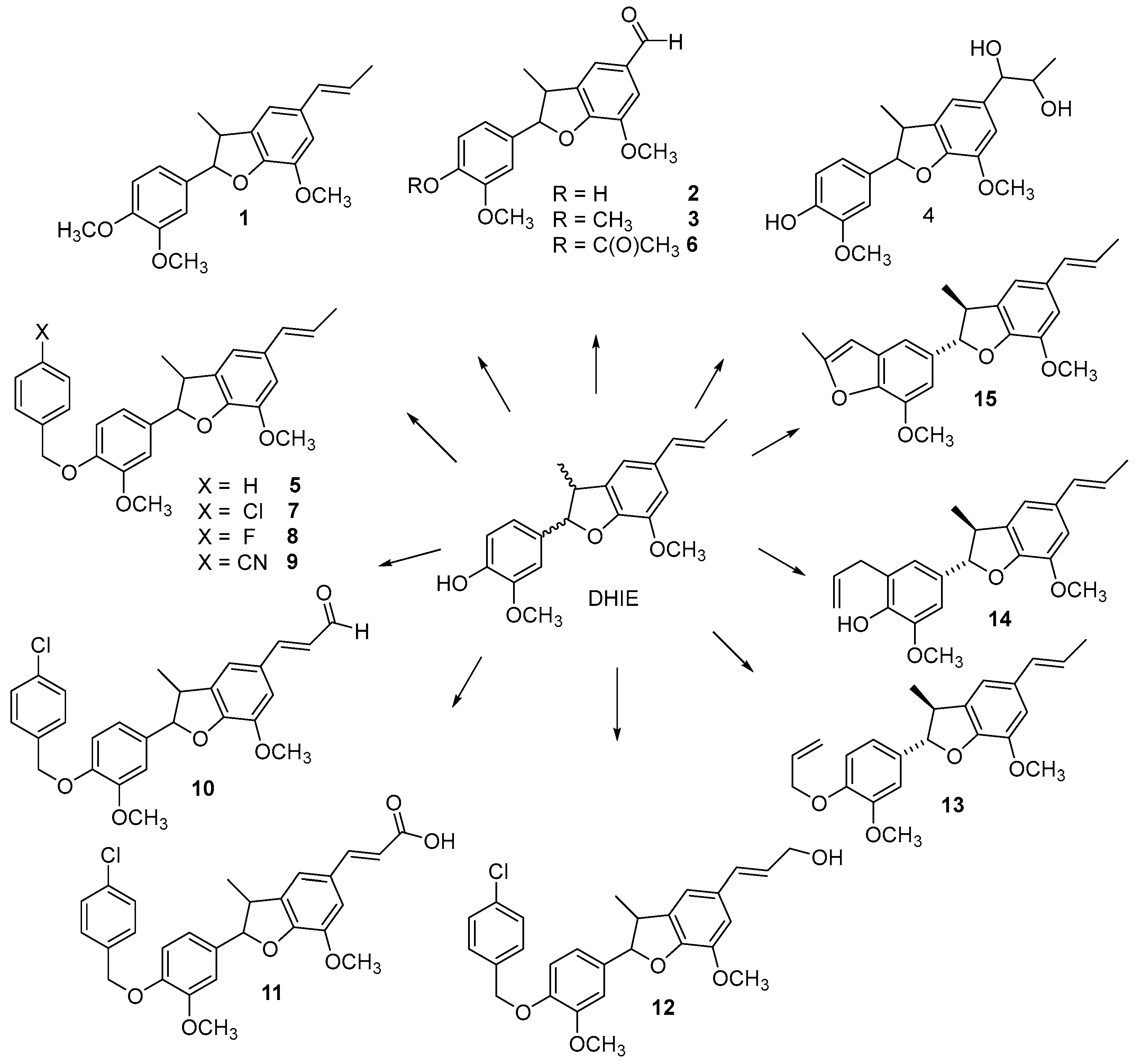
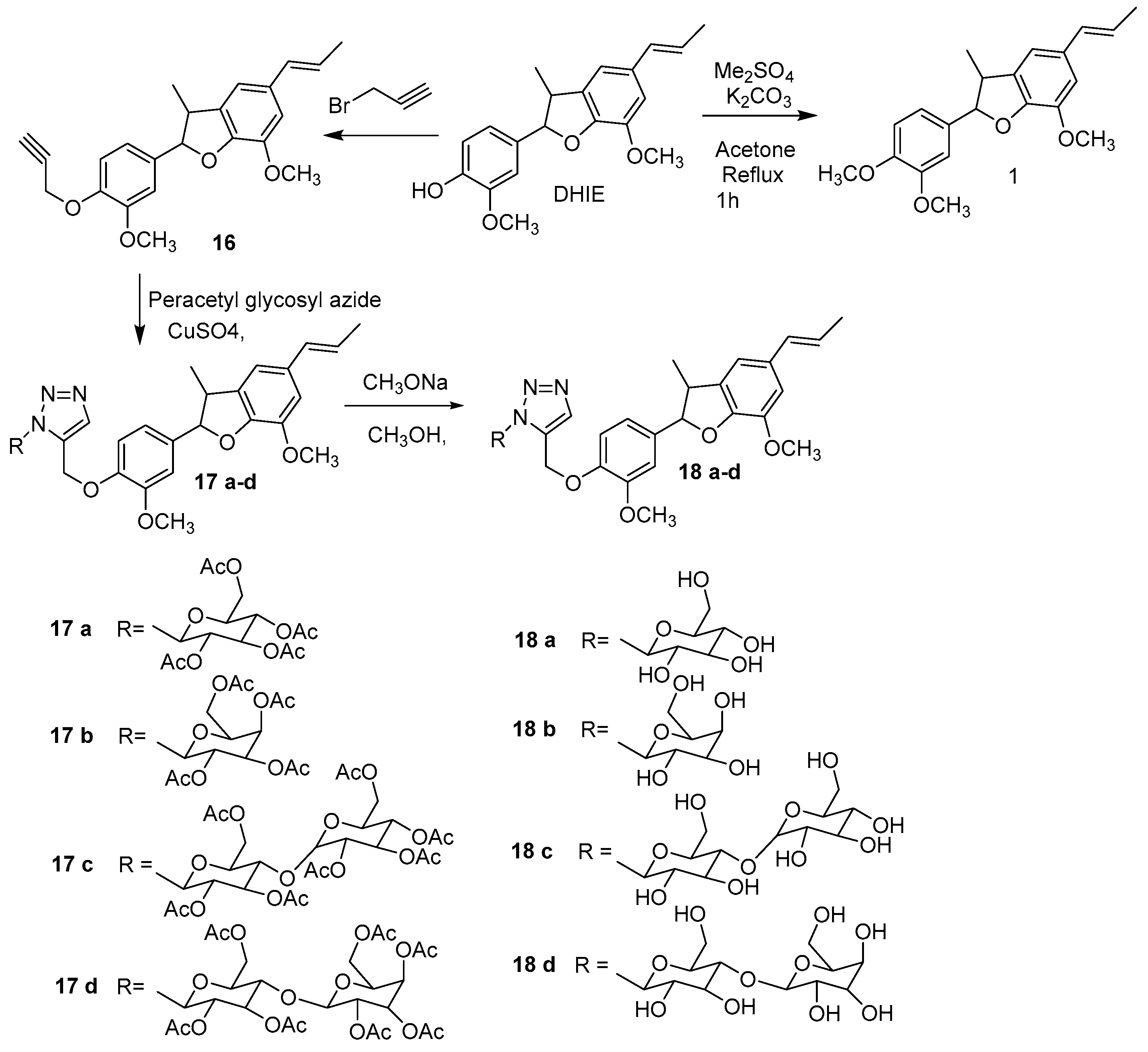
5. Synthesis of DHIE Derivatives
6. Biological Activity of DHIE
6.1. Anti-Oxidant Effect of DHIE
6.2. Anti-Parasitic Activity of DHIE
6.3. Anti-Bacterial Activity of DHIE
6.4. Anticancer Activity of DHIE
6.5. Anti-Inflammatory Effect of DHIE
6.6. Other Biological Properties of DHIE
7. Pharmacokinetic of DHIE
8. Biological Activity of Semi-Synthetic DHIE Derivatives
9. Conclusions
10. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pauletti, P.M.; Araújo, A.R.; Young, M.C.M.; Giesbrecht, A.M.; Da Silva Bolzani, V. nor-lignans from the leaves of Styrax ferrugineus (Styracaceae) with antibacterial and antifungal activity. Phytochemistry 2000, 55, 597–601. [Google Scholar] [CrossRef]
- Saguez, J.; Dubois, F.; Vincent, C.; Laberche, J.C.; Sangwan-Norreel, B.S.; Giordanengo, P. Differential aphicidal effects of chitinase inhibitors on the polyphagous homopteran Myzus persicae (Sulzer). Pest Manag. Sci. 2006, 62, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Talapatra, S.K.; Talapatra, B. Shikimic acid pathway. In Chemistry of Plant Natural Products; Springer: Berlin/Heidelberg, Germany, 2015; pp. 625–678. [Google Scholar]
- Suzuki, S.; Umezawa, T. Biosynthesis of lignans and norlignans. J. Wood Sci. 2007, 53, 273–284. [Google Scholar] [CrossRef]
- Moss, G.P. Nomenclature of lignans and neolignans (IUPAC recommendations 2000). Pure Appl. Chem. 2000, 72, 1493–1523. [Google Scholar] [CrossRef]
- Dar, A.A.; Arumugam, N. Lignans of sesame: Purification methods, biological activities and biosynthesis—A review. Bioorg. Chem. 2013, 50, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.E.; Heacock, R.A.; Forrest, T.P. The isolation of dehydrodiisoeugenol from the aril of Myristica fragrans Houtt. Experientia 1973, 29, 139. [Google Scholar] [CrossRef]
- Aiba, C.J.; Corrêa, R.G.C.; Gottlieb, O.R. Natural occurrence of erdtman’s dehydrodiisoeugenol. Phytochemistry 1973, 12, 1163–1164. [Google Scholar] [CrossRef]
- León-Díaz, R.; Meckes-Fischer, M.; Valdovinos-Martínez, L.; Campos, M.G.; Hernández-Pando, R.; Jiménez-Arellanes, M.A. Antitubercular activity and the subacute toxicity of (−)-Licarin A in BALB/c mice: A neolignan isolated from Aristolochia taliscana. Arch. Med. Res. 2013, 44, 99–104. [Google Scholar] [CrossRef]
- Cabral, M.M.O.; Barbosa-Filho, J.M.; Maia, G.L.A.; Chaves, M.C.O.; Braga, M.V.; De Souza, W.; Soares, R.O.A. Neolignans from plants in northeastern Brazil (Lauraceae) with activity against Trypanosoma cruzi. Exp. Parasitol. 2010, 124, 319–324. [Google Scholar] [CrossRef]
- Li, C.; Zhang, K.; Pan, G.; Ji, H.; Li, C.; Wang, X.; Hu, X.; Liu, R.; Deng, L.; Wang, Y.; et al. Dehydrodiisoeugenol inhibits colorectal cancer growth by endoplasmic reticulum stress-induced autophagic pathways. J. Exp. Clin. Cancer Res. 2021, 40, 125. [Google Scholar] [CrossRef]
- Fujisawa, S.; Ishihara, M.; Murakami, Y.; Atsumi, T.; Kadoma, Y.; Yokoe, I. Predicting the biological activities of 2-methoxyphenol antioxidants: Effects of dimers. In Vivo 2007, 21, 181–188. [Google Scholar] [PubMed]
- León-Díaz, R.; Meckes, M.; Said-Fernández, S.; Molina-Salinas, G.M.; Vargas-Villarreal, J.; Torres, J.; Luna-Herrera, J.; Jiménez-Arellanes, A. Antimycobacterial neolignans isolated from Aristolochia taliscana. Mem. Inst. Oswaldo Cruz 2010, 105, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamilpa, A.; Abarca-Vargas, R.; Ventura-Zapata, E.; Osuna-Torres, L.; Zavala, M.A.; Herrera-Ruiz, M.; Jiménez-Ferrer, E.; González-Cortazar, M. Neolignans from Aristolochia elegans as Antagonists of the neurotropic effect of scorpion venom. J. Ethnopharmacol. 2014, 157, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Filho, J.M.; Yoshida, M.; Gottlieb, O.R. Lignoids from Nectandra amazonum and N. glabrescens. Phytochemistry 1989, 28, 1991. [Google Scholar] [CrossRef]
- Le Quesne, P.W.; Larrahondo, J.E.; Raffauf, R.F. Antitumor Plants. X. Constituents of Nectandra rigida. J. Nat. Prod. 2004, 43, 353–359. [Google Scholar] [CrossRef]
- Mengarda, A.C.; Silva, M.P.; Cirino, M.E.; Morais, T.R.; Conserva, G.A.A.; Lago, J.H.G.; de Moraes, J. Licarin A, a neolignan isolated from Nectandra oppositifolia Nees & Mart. (Lauraceae), exhibited moderate preclinical efficacy against Schistosoma mansoni infection. Phyther. Res. 2021, 35, 5154–5162. [Google Scholar] [CrossRef]
- Morais, T.R.; Conserva, G.A.A.; Varela, M.T.; Costa-Silva, T.A.; Thevenard, F.; Ponci, V.; Fortuna, A.; Falcão, A.C.; Tempone, A.G.; Fernandes, J.P.S.; et al. Improving the drug-likeness of inspiring natural products-evaluation of the antiparasitic activity against Trypanosoma cruzi through semi-synthetic and simplified analogues of Licarin A. Sci. Rep. 2020, 10, 5467. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.J.; Sung, S.H.; Kim, Y.C. Neuroprotective lignans from the bark of Machilus thunbergii. Planta Med. 2004, 70, 79–80. [Google Scholar] [CrossRef]
- Li, F.; Yang, X.W. Three new neolignans from the aril of Myristica fragrans. Helv. Chim. Acta 2007, 90, 1491–1496. [Google Scholar] [CrossRef]
- Francis, K.S.; Suresh, E.; Nair, M.S. Chemical constituents from Myristica fragrans fruit. Nat. Prod. Res. 2014, 28, 1664–1668. [Google Scholar] [CrossRef]
- Read, R.; Taylor, W. Constituents of Eupomatia Species. V. The Isolation of eupomatenoid-13 (a new neolignan), (±)-trans-dehydrodiisoeugenol, and other extractives from the bark of Eupomatia Laurina. Aust. J. Chem. 1979, 32, 2317–2321. [Google Scholar] [CrossRef]
- Ito, K.; Ichino, K.; Iida, T.; Lai, J. Neolignans from Magnolia kachirachirai. Phytochemistry 1984, 23, 2643–2645. [Google Scholar] [CrossRef]
- Ma, Y.; Han, G.Q.; Wang, Y.Y. PAF Antagonistic benzofuran neolignans from Piper kadsura. Yao Xue Xue Bao 1993, 28, 370–373. [Google Scholar] [PubMed]
- Duan, S.; Zhang, P.; Yu, P. Neolignans and lignan from Piper wallichii. Zhongguo Zhongyao Zazhi 2010, 35, 180–182. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Wamba, B.E.N.; Bitchagno, G.T.M.; Tankeo, S.B.; Çelik, İ.; Atontsa, B.C.K.; Nkuété Lonfouo, A.H.; Kuete, V.; Efferth, T. Bioactivity of fractions and constituents of Piper capense fruits towards a broad panel of cancer cells. J. Ethnopharmacol. 2021, 271, 113884. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.G.; Phan, T.S.; Matsunami, K.; Otsuka, H. New neolignans and lignans from vietnamese medicinal plant Machilus odoratissima NEES. Chem. Pharm. Bull. 2006, 54, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y.; Ko, H.H.; Lee, S.J.; Chang, H.S.; Lin, C.H.; Chen, I.S. Biological evaluation of secondary metabolites from the root of Machilus obovatifolia. Chem. Biodivers. 2015, 12, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.F.L.; Ehrenfried, C.A.; Riva, D.; Barison, A.; De Mello-Silva, R.; Stefanello, M.E.A. Essential oil and other constituents from Magnolia ovata fruit. Nat. Prod. Commun. 2012, 7, 1365–1367. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.F.L.; Barison, A.; Salvador, M.J.; De Mello-Silva, R.; Cabral, E.C.; Eberlin, M.N.; Stefanello, M.É.A. Constituents of the leaves of Magnolia ovata. J. Nat. Prod. 2009, 72, 1529–1532. [Google Scholar] [CrossRef]
- Chen, H.; Ji, T.; Chen, J.; Li, X. Matrix solid-phase dispersion combined with HPLC-DAD for simultaneous determination of nine lignans in Saururus chinensis. J. Chromatogr. Sci. 2019, 57, 186–193. [Google Scholar] [CrossRef]
- De, F.; Dias, A.; Giesbrecht, A.M.; Gottlieb, O.R. Neolignans from Urbanodendron verrucosum. Phytochemistry 1982, 21, 1137–1139. [Google Scholar] [CrossRef]
- Chen, Y.P.; Wang, Z.P.; Zheng, H.H.; Xu, Y.T.; Zhu, Y.; Zhang, P.; Wu, H.H. A New caffeate compound from Nardostachys chinensis. Yao Xue Xue Bao 2016, 51, 100–104. [Google Scholar] [PubMed]
- Leopold, B.; Hassel, O.; Lund, E.W. Aromatic keto- and hydroxy-polyethers as lignin models. III. Acta Chem. Scand. 1950, 4, 1523–1537. [Google Scholar] [CrossRef] [Green Version]
- Levita, J.; Mardhatillah, A.; Mutakin, M.; Yani, A.; Terusan Jenderal, J.; Cimahi, S.; Java, W. FeCl3-catalyzed synthesis of dehydrodiisoeugenol. Artic. Int. J. Chem. 2015, 8, 1–7. [Google Scholar] [CrossRef]
- Liu, S.-y.; Wang, G.-q.; Liang, Z.-y.; Wang, Q.-a. Synthesis of dihydrobenzofuran neoligans licarin a and dihydrocarinatin as well as related triazolylglycosides. Chem. Res. Chin. Univ. 2013, 29, 1119–1124. [Google Scholar] [CrossRef]
- Chen, P.Y.; Wu, Y.H.; Hsu, M.H.; Wang, T.P.; Wang, E.C. Cerium ammonium nitrate-mediated the oxidative dimerization of p-alkenylphenols: A new synthesis of substituted (±)-trans-dihydrobenzofurans. Tetrahedron 2013, 69, 653–657. [Google Scholar] [CrossRef]
- Juhász, L.; Kürti, L.; Antus, S. Simple synthesis of benzofuranoid neolignans from Myristica fragrans. J. Nat. Prod. 2000, 63, 866–870. [Google Scholar] [CrossRef]
- Chioccara, F.; Poli, S.; Rindone, B.; Pilati, T.; Brunow, G.; Pietikäinen, P.; Setälä, H.; Trætteberg, M.; Nasiri, A.; Tsuda, T. Regio- and diastereo-selective synthesis of dimeric lignans using oxidative coupling. Acta Chem. Scand. 1993, 47, 610–616. [Google Scholar] [CrossRef]
- Krawczyk, A.R.; Lipkowska, E.; Wróbel, J.T. Horseradish peroxidase-mediated preparation of dimers from eugenol and isoeugenol. Collect. Czechoslov. Chem. Commun. 1991, 56, 1147–1150. [Google Scholar] [CrossRef]
- Nascimento, I.R.; Lopes, L.M.X.; Davin, L.B.; Lewis, N.G. Stereoselective synthesis of 8,9-licarinediols. Tetrahedron 2000, 56, 9181–9193. [Google Scholar] [CrossRef]
- Bortolomeazzi, R.; Verardo, G.; Liessi, A.; Callea, A. Formation of dehydrodiisoeugenol and dehydrodieugenol from the reaction of isoeugenol and eugenol with DPPH radical and their role in the radical scavenging activity. Food Chem. 2010, 118, 256–265. [Google Scholar] [CrossRef]
- Pereira, A.C.; Magalhães, L.G.; Gonalves, U.O.; Luz, P.P.; Moraes, A.C.G.; Rodrigues, V.; Da Matta Guedes, P.M.; Da Silva Filho, A.A.; Cunha, W.R.; Bastos, J.K.; et al. Schistosomicidal and trypanocidal structure–activity relationships for (±)-licarin A and its (−)- and (+)-enantiomers. Phytochemistry 2011, 72, 1424–1430. [Google Scholar] [CrossRef]
- Rodrigues, L.C.; Barbosa-Filho, J.M.; Marques, S.D.G.; Borges, F.V.P.; de Araújo Silva, L.A.; Herrera Bravo de Laguna, I.; Mioso, R. Formation of bioactive benzofuran via oxidative coupling, using coconut water (Cocos nucifera L.) as biocatalyst. Org. Commun. 2017, 10, 72–78. [Google Scholar] [CrossRef]
- Néris, P.L.N.; Caldas, J.P.A.; Rodrigues, Y.K.S.; Amorim, F.M.; Leite, J.A.; Rodrigues-Mascarenhas, S.; Barbosa-Filho, J.M.; Rodrigues, L.C.; Oliveira, M.R. Neolignan licarin a presents effect against Leishmania (Leishmania) major associated with immunomodulation in vitro. Exp. Parasitol. 2013, 135, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba, T.; Xiao, L.; Miyakoshi, T.; Chen, C.L. Oxidation of isoeugenol and coniferyl alcohol catalyzed by laccases isolated from Rhus vernicifera stokes and Pycnoporus coccineus. J. Mol. Catal. B Enzym. 2000, 10, 605–615. [Google Scholar] [CrossRef]
- Hernández-Vázquez, L.; Olivera-Flores, M.T.D.J.; Ruíz-Terán, F.; Ayala, I.; Navarro-Ocaña, A. Screening of plant cell cultures for their capacity to dimerize eugenol and isoeugenol: Preparation of dehydrodieugenol. J. Mol. Catal. B Enzym. 2011, 72, 102–106. [Google Scholar] [CrossRef]
- Hernández-Vázquez, L.; de Jesús Olivera-Flores, M.T.; Luna, H.; Navarro-Ocaña, A. Obtención de dehidrodiisoeugenol por dimerización de isoeugenol con cultivos celulares de Bouvardia ternifolia (trompetilla). TIP Rev. Esp. Cienc. Quím. Bio. 2017, 20, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.; Bhat, S.V. Biotransformation of isoeugenol catalyzed by growing cells of Pseudomonas putida. Biocatal. Biotransformation 2011, 29, 147–150. [Google Scholar] [CrossRef]
- Hua, D.; Ma, C.; Lin, S.; Song, L.; Deng, Z.; Maomy, Z.; Zhang, Z.; Yu, B.; Xu, P. Biotransformation of isoeugenol to vanillin by a newly isolated Bacillus pumilus strain: Identification of major metabolites. J. Biotechnol. 2007, 130, 463–470. [Google Scholar] [CrossRef]
- Oliveira, B.H.M.; Silva, L.A.A.; Leite, F.F.; Duarte, G.D.; Duarte, S.S.; Silva, D.K.F.; Sobral, M.V.; Rodrigues, L.C. Síntese e avaliação antitumoral do (±)-4-o-demethylkadsurenin M um composto natural e seus análogos. Braz. J. Dev. 2019, 5, 19109–19123. [Google Scholar] [CrossRef]
- Meleti, V.R.; Esperandim, V.R.; Flauzino, L.G.B.; Prizantelli, A.H.; de Lima Paula, L.A.; Magalhães, L.G.; Cunha, W.R.; da Silva Laurentiz, R.; da Rocha Pissurno, A.P.; Nanayakkara, N.P.D.; et al. (±)-Licarin A and its semi-synthetic derivatives: In vitro and in silico evaluation of trypanocidal and schistosomicidal activities. Acta Trop. 2020, 202, 105248. [Google Scholar] [CrossRef] [PubMed]
- El-Feraly, F.S.; Cheatham, S.F.; Hufford, C.D.; Li, W.S. Optical resolution of (±)-dehydrodiisoeugenol: Structure revision of acuminatin. Phytochemistry 1982, 21, 1133–1135. [Google Scholar] [CrossRef]
- Alvarenga, D.J.; Matias, L.M.F.; Oliveira, L.M.; de Oliveira Leão, L.P.M.; Hawkes, J.A.; Raimundo, B.V.B.; de Figueiredo Diniz Castro, L.; de Campos, M.M.A.; dos Santos Siqueira, F.; dos Santos, T.; et al. Exploring how structural changes to new licarin a derivatives effects their bioactive properties against rapid growing mycobacteria and biofilm formation. Microb. Pathog. 2020, 144, 104203. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choong, J.M.; So, R.K.; Kim, J.; Young, C.K. Meso-dihydroguaiaretic acid and licarin A of Machilus thunbergii protect against glutamate-induced toxicity in primary cultures of a rat cortical cells. Br. J. Pharmacol. 2005, 146, 752. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.-P.; Wu, H.; Wang, Y.; Weng, X.-C. Isolation of some compounds from nutmeg and their antioxidant activities. Czech J. Food Sci. 2012, 30, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Shoji, M.; Hirata, A.; Tanaka, S.; Yokoe, I.; Fujisawa, S. Dehydrodiisoeugenol, an Isoeugenol Dimer, Inhibits Lipopolysaccharide-Stimulated Nuclear Factor Kappa B Activation and Cyclooxygenase-2 Expression in Macrophages. Arch. Biochem. Biophys. 2005, 434, 326–332. [Google Scholar] [CrossRef]
- Li, C.W.; Chu, Y.C.; Huang, C.Y.; Fu, S.L.; Chen, J.J. Evaluation of antioxidant and anti-α-glucosidase activities of various solvent extracts and major bioactive components from the Seeds of Myristica fragrans. Molecules 2020, 25, 5198. [Google Scholar] [CrossRef]
- Mitra, A.K.; Mawson, A.R. Neglected tropical diseases: Epidemiology and global burden. Trop. Med. Infect. Dis. 2017, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Nagafuji, S.; Yamauchi, T.; Okabe, H.; Maki, J.; Higo, H.; Akahane, H.; Aguilar, A.; Jiménez-Estrada, M.; Reyes-Chilpa, R. Trypanocidal constituents in plants 1. Evaluation of some Mexican plants for their trypanocidal activity and active constituents in Guaco, roots of Aristolochia taliscana. Biol. Pharm. Bull. 2002, 25, 1188–1191. [Google Scholar] [CrossRef]
- Schön, T.; Miotto, P.; Köser, C.U.; Viveiros, M.; Böttger, E.; Cambau, E. Mycobacterium tuberculosis drug-resistance testing: Challenges, recent developments and perspectives. Clin. Microbiol. Infect. 2017, 23, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Arellanes, A.M.; Román-Cortés, N.R.; García, I. Antioxidant and antimicrobial activities of hexane extracts and pure compounds from Aristolochia taliscana rhizome. Rev. Mex. Cienc. Farm. 2011, 42, 35–41. [Google Scholar]
- Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; Xu, R.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life Years for 29 Cancer Groups From 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, J.; Young, U.Y.; Young, C.K. Inhibition of phospholipase Cγ1 and cancer cell proliferation by lignans and flavans from Machilus thunbergii. Arch. Pharmacal Res. 2004, 27, 1043–1047. [Google Scholar] [CrossRef]
- Maheswari, U.; Ghosh, K.; Sadras, S.R. Licarin A induces cell death by activation of autophagy and apoptosis in non-small cell lung cancer cells. Apoptosis 2018, 23, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Ghelli, F.; Panizzolo, M.; Garzaro, G.; Squillacioti, G.; Bellisario, V.; Colombi, N.; Bergamaschi, E.; Canu, I.G.; Bono, R. Inflammatory biomarkers in exhaled breath condensate: A systematic review. Int. J. Mol. Sci. 2022, 23, 9820. [Google Scholar] [CrossRef]
- Walton, R.M.; Siegel, A. Avian inflammatory markers. Vet. Clin. N. Am.-Exot. Anim. Pract. 2022, 25, 679–695. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Ito, C.; Masubuchi, S.; Itoigawa, M. Licarin A Is a Candidate compound for the treatment of immediate hypersensitivity via inhibition of rat mast cell line RBL-2H3 cells. J. Pharm. Pharmacol. 2015, 67, 1723–1732. [Google Scholar] [CrossRef]
- Morikawa, T.; Hachiman, I.; Ninomiya, K.; Hata, H.; Sugawara, K.; Muraoka, O.; Matsuda, H. Degranulation inhibitors from the arils of Myristica fragrans in antigen-stimulated rat basophilic leukemia cells. J. Nat. Med. 2018, 72, 464–473. [Google Scholar] [CrossRef]
- Takeyoshi, M.; Iida, K.; Suzuki, K.; Yamazaki, S. Skin sensitization potency of isoeugenol and its dimers evaluated by a non-radioisotopic modification of the local lymph node assay and guinea pig maximization test. J. Appl. Toxicol. 2008, 28, 530–534. [Google Scholar] [CrossRef]
- Saptarini, N.M.; Saputri, F.A.; Levita, J. Molecular modeling study of PPAR γ agonists: Dehydro-di-isoeugenol, macelignan, pioglitazone, netoglitazone, and rosiglitazone as antidiabetic drugs. Int. J. Chem. 2014, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Muchrtaridi, M.; Lestari, K. In silico Evaluation of potent for PPARγ agonist of lignan derivatives from Myristica fragrans Houtt seeds. Int. J. Pharm. Pharm. Sci. 2014, 6, 795–800. [Google Scholar]
- Yoon, D.; Imran, K.M.; Kim, Y.S. Distinctive effects of licarin A on lipolysis mediated by PKA and on formation of brown adipocytes from C3H10T1/2 mesenchymal stem cells. Toxicol. Appl. Pharmacol. 2018, 340, 9–20. [Google Scholar] [CrossRef]
- El-Alfy, A.T.; Abourashed, E.A.; Patel, C.; Mazhari, N.; An, H.R.; Jeon, A. Phenolic Compounds from Nutmeg (Myristica fragrans Houtt.) Inhibit the endocannabinoid-modulating enzyme fatty acid amide hydrolase. J. Pharm. Pharmacol. 2019, 71, 1879–1889. [Google Scholar] [CrossRef]
- Li, F.; Yang, X.W. Determination of dehydrodiisoeugenol in rat tissues using HPLC method. Biomed. Chromatogr. 2008, 22, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Zhu, L.Q.; Yang, X.W. Cerebral nuclei distribution study of dehydrodiisoeugenol as an anxiogenic agent determined by RP-HPLC. Fitoterapia 2013, 84, 47–53. [Google Scholar] [CrossRef]
- Wu, N.; Xu, W.; Cao, G.Y.; Yang, Y.F.; Yang, X.B.; Yang, X.W. The blood-brain barrier permeability of lignans and malabaricones from the seeds of Myristica fragrans in the MDCK-PHaMDR cell monolayer model. Molecules 2016, 21, 134. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yang, X.W. Simultaneous determination of diastereomers (+)-licarin A and isolicarin A from Myristica fragrans in rat plasma by HPLC and its application to their pharmacokinetics. Planta Med. 2008, 74, 880–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yang, X.W. Metabolism of the lignan dehydrodiisoeugenol in rats. Planta Med. 2011, 77, 1712–1717. [Google Scholar] [CrossRef]
- Li, F.; Yang, X.W. Analysis of anti-inflammatory dehydrodiisoeugenol and metabolites excreted in rat feces and urine using HPLC-UV. Biomed. Chromatogr. 2012, 26, 703–707. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Part | Ref |
|---|---|---|
| Aristolochia taliscana | Roots | [13] |
| Aristolochia elegans | Wild roots | [14] |
| Nectandra glabrescens | Fruits | [15] |
| Nectandra rigida | Bark and stems | [16] |
| Nectandra oppositifolia | Leaves | [17,18] |
| Machilus thunbergii | Bark | [19] |
| Myristica fragrans | Aril | [20,21] |
| Eupomatia laurina | Bark | [22] |
| Magnolia kachirachirai | Leaves | [23] |
| Piper kadsura | Aerial parts | [24] |
| Piper wallichii | Aerial parts | [25] |
| Piper capense | Fruits | [26] |
| Licaria aritu | Wood | [8] |
| Machilius odoratissima | Bark | [27] |
| Machilus obovatifolia | Root | [28] |
| Magnolia ovata | Unripe fruits Leaves | [29] [30] |
| Saururus chinensis | Aerial parts | [31] |
| Urbanodendron verrucosum | Branch wood | [32] |
| Nardostachys chinensis | Roots and rhizomes | [33] |
| Catalyst | Mol-Ratio | Solvent | T (°C) | Reaction Time (h) | Yield (%) | Reference |
|---|---|---|---|---|---|---|
| FeCl3 | 1.4 | water-ethanol | 4 | 24 | 30 | [34] |
| FeCl3 | 0.5 | water-ethanol | 5 | 24 | 22 | [35] |
| Ag2O | 0.5 | toluene-acetone | −70 | 48 | 40 | [36] |
| CAN | 1.5 | tetrahydrofuran | room temp | 0.5 | 81 | [37] |
| IDA | 0.62 | dichloromethane | room temp | 48 | 35 | [38] |
| IE (mmol) | Methanol/Buffer (%) | U | H2O2 (mmol) | T (h) | DHIE | Yield (%) | Reference |
|---|---|---|---|---|---|---|---|
| 6 | 26:74 Phosphoric pH = 6.0 | 4.5–7.5 × 10−6 mmol | 0.5 | 24 | NR | 22 | [40] |
| 4.9–10 | 10:90 Citrate-phosphate pH = 3.0 | 2150–4300 | 2.45–5 | 1 | (±) | 60 | [39] |
| 10 | 10:90 Citrate-phosphate pH = 3.0 | 2500 | 5 | 1 | (±) | 99 | [41] |
| 6.2 | 10:90 Citrate-phosphate pH = 3.0 | 1500 | 3.05 | 0.33 | (±) | 27 | [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godínez-Chaparro, B.; Pérez-Gutiérrez, S.; Pérez-Ramos, J.; Heyerdahl-Viau, I.; Hernández-Vázquez, L. Synthesis and Biological Activities of Dehydrodiisoeugenol: A Review. Pharmaceuticals 2022, 15, 1351. https://doi.org/10.3390/ph15111351
Godínez-Chaparro B, Pérez-Gutiérrez S, Pérez-Ramos J, Heyerdahl-Viau I, Hernández-Vázquez L. Synthesis and Biological Activities of Dehydrodiisoeugenol: A Review. Pharmaceuticals. 2022; 15(11):1351. https://doi.org/10.3390/ph15111351
Chicago/Turabian StyleGodínez-Chaparro, Beatriz, Salud Pérez-Gutiérrez, Julia Pérez-Ramos, Ivo Heyerdahl-Viau, and Liliana Hernández-Vázquez. 2022. "Synthesis and Biological Activities of Dehydrodiisoeugenol: A Review" Pharmaceuticals 15, no. 11: 1351. https://doi.org/10.3390/ph15111351
APA StyleGodínez-Chaparro, B., Pérez-Gutiérrez, S., Pérez-Ramos, J., Heyerdahl-Viau, I., & Hernández-Vázquez, L. (2022). Synthesis and Biological Activities of Dehydrodiisoeugenol: A Review. Pharmaceuticals, 15(11), 1351. https://doi.org/10.3390/ph15111351