
Tibouchina granulosa Leaves Present Anti-Inflammatory Effect

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
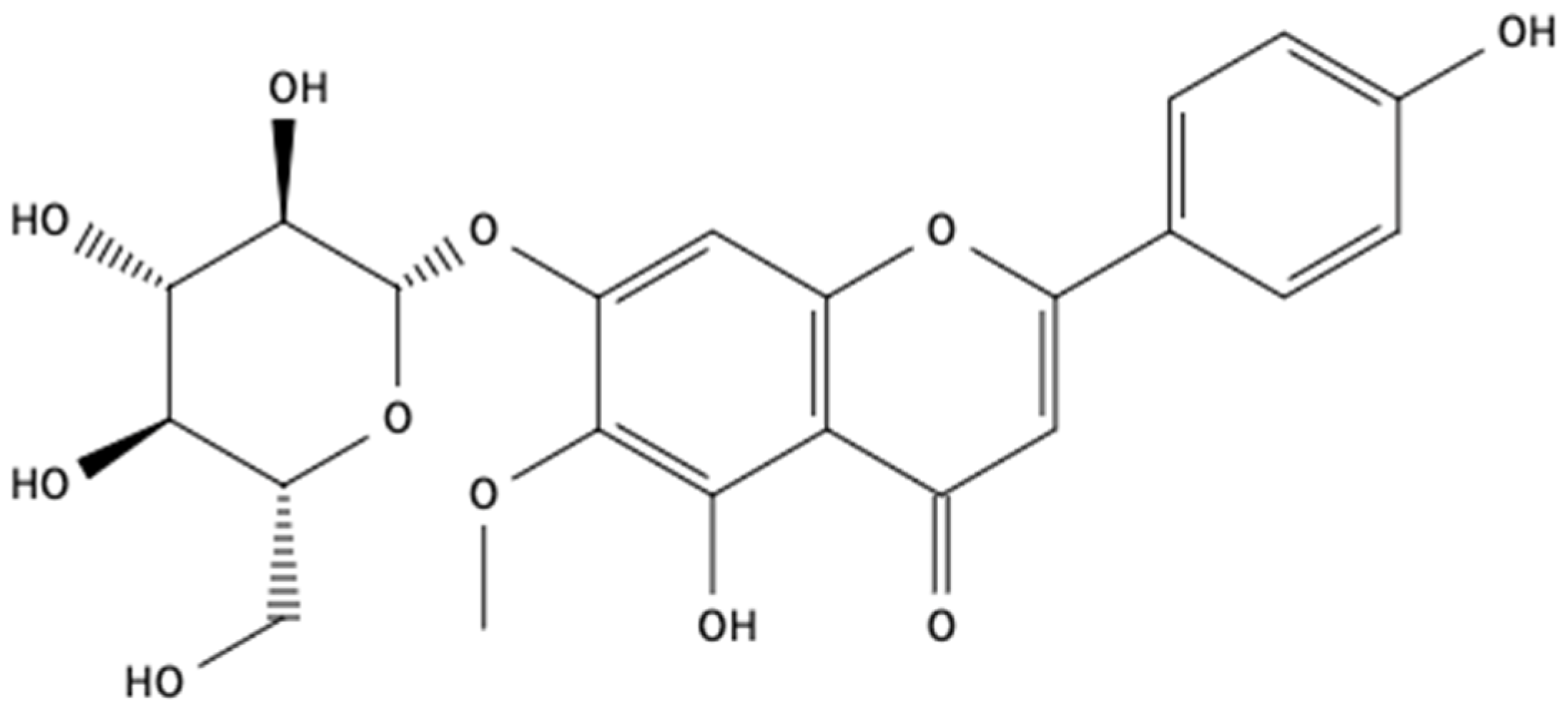
2.1. Chemical Analysis
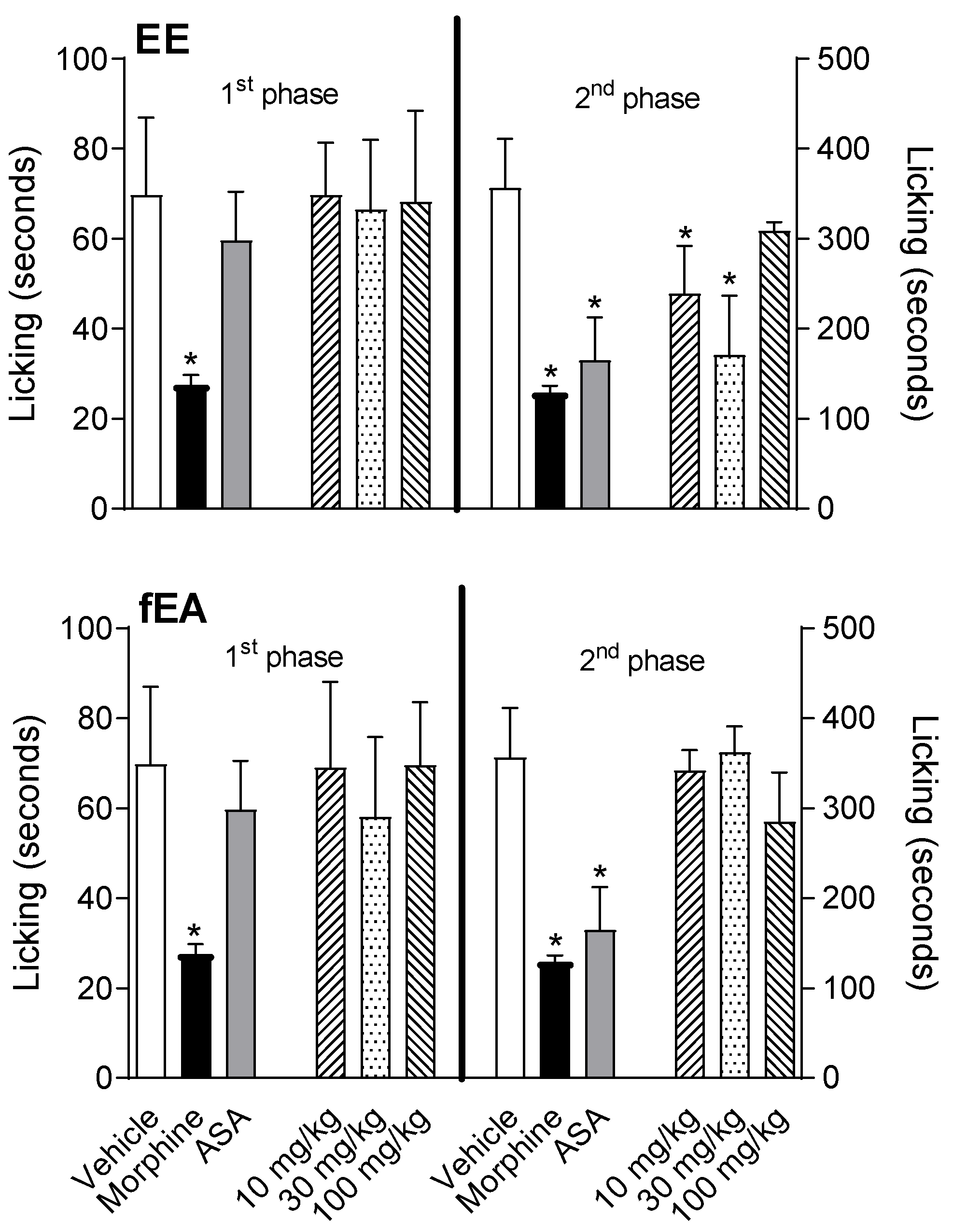
2.2. T. granulosa EE and fEA Reduced Paw Licking Behavior Induced by Formalin Injection
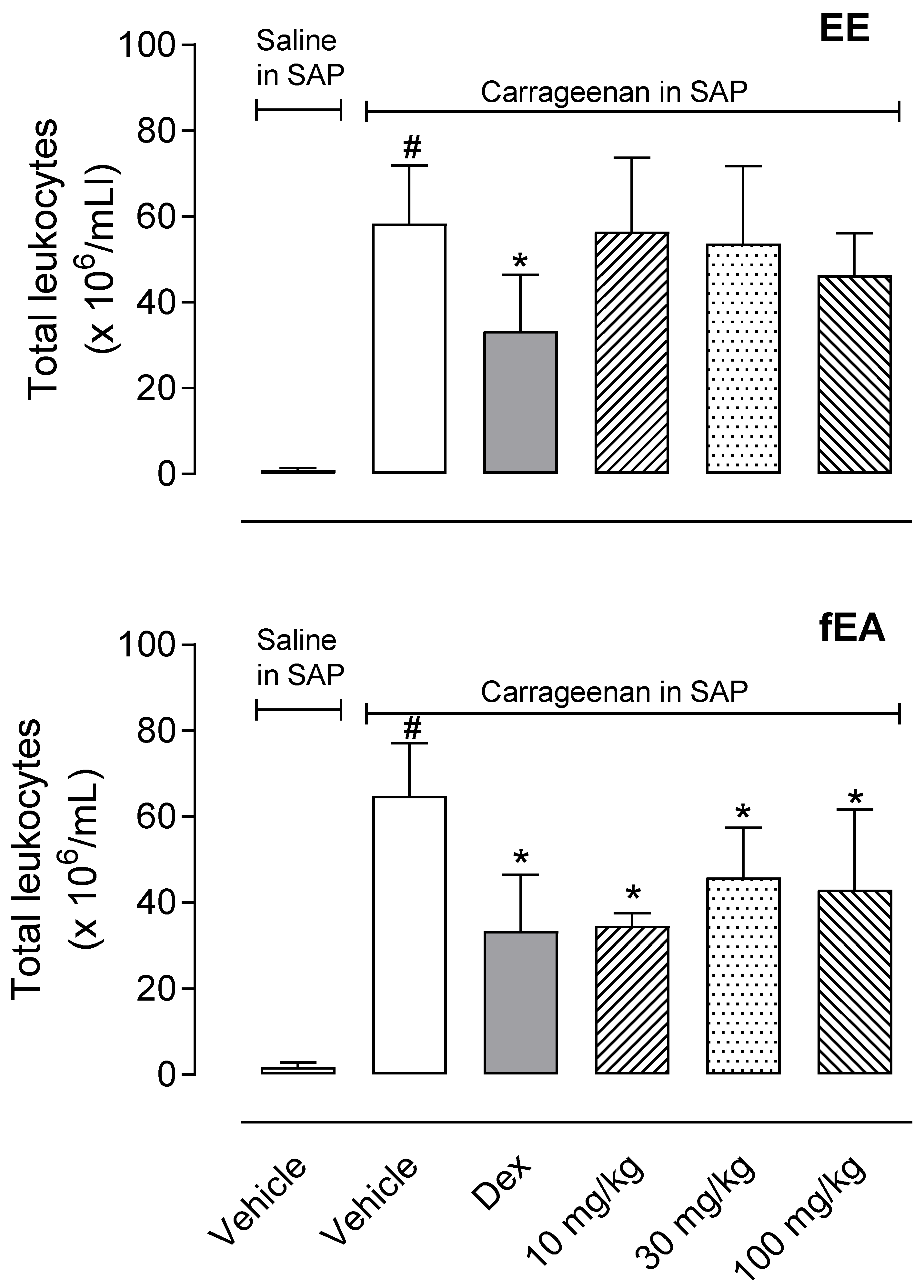
2.3. T. granulosa fEA Reduced Leukocytes Migration into Subcutaneous Air Pouch (SAP)
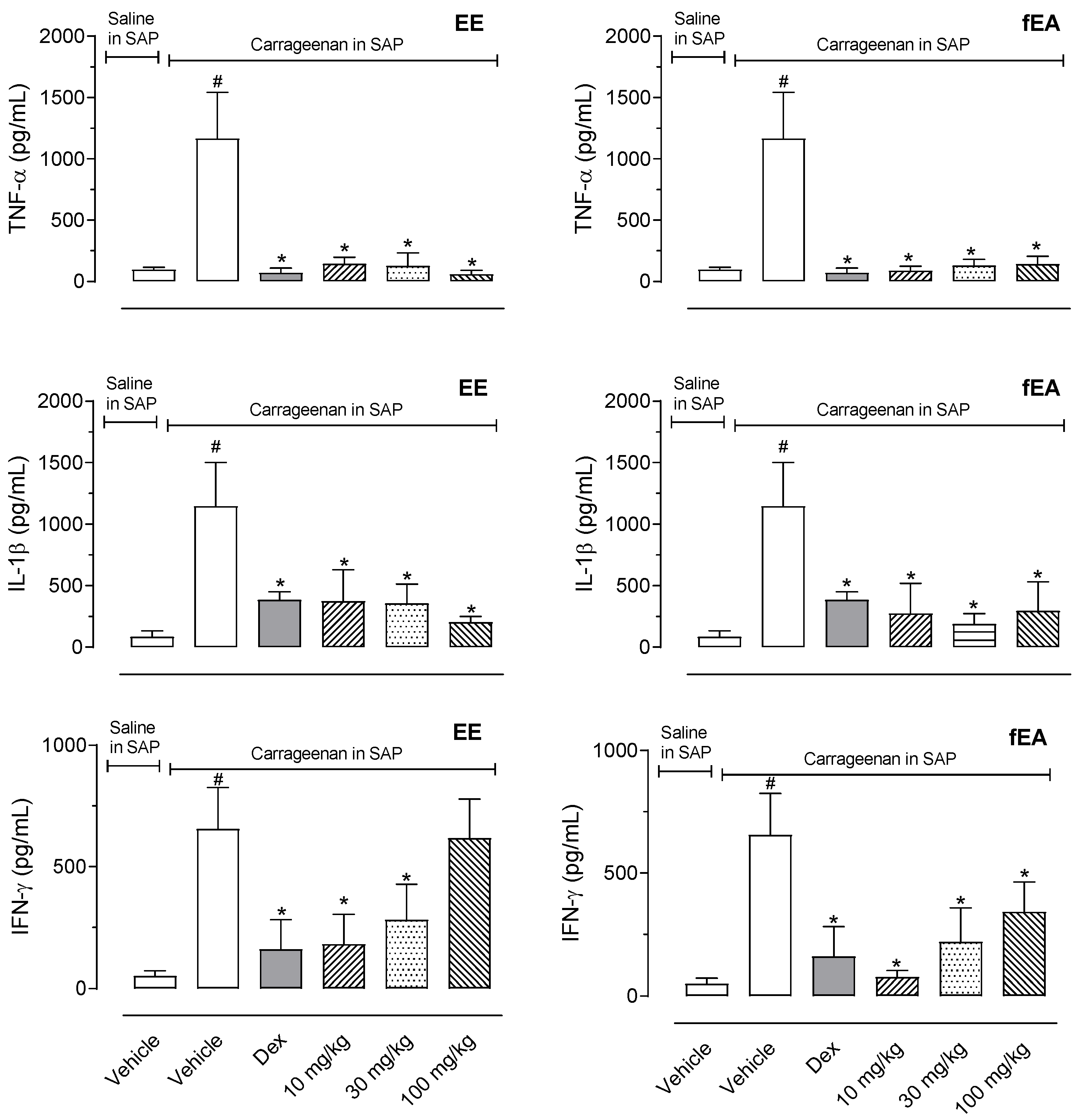
2.4. T. granulosa EE and fEA Reduced Inflammatory Mediators’ Productions in SAP
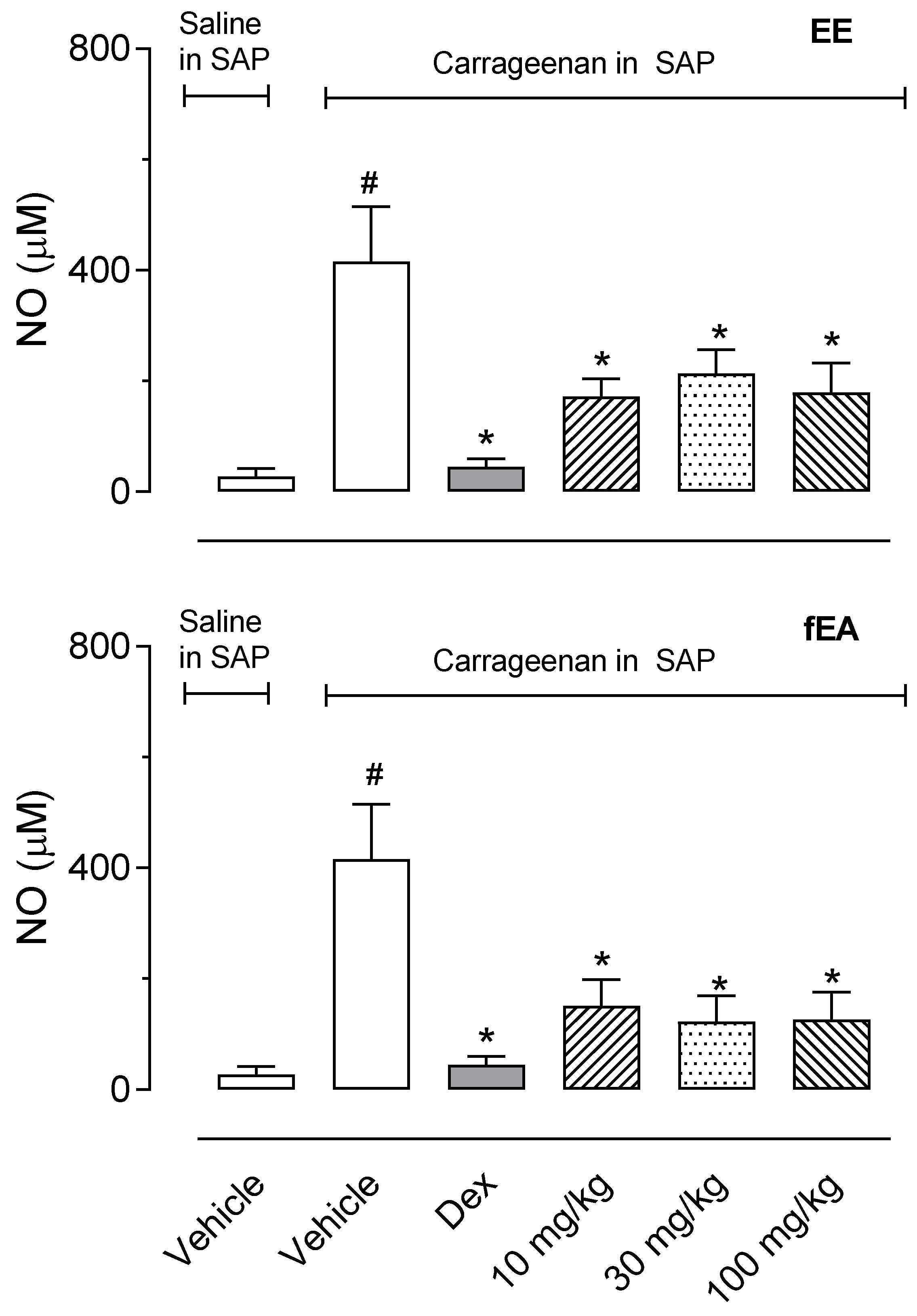
2.5. T. granulosa EE and fEA Reduced the Production of Nitric Oxide in SAP
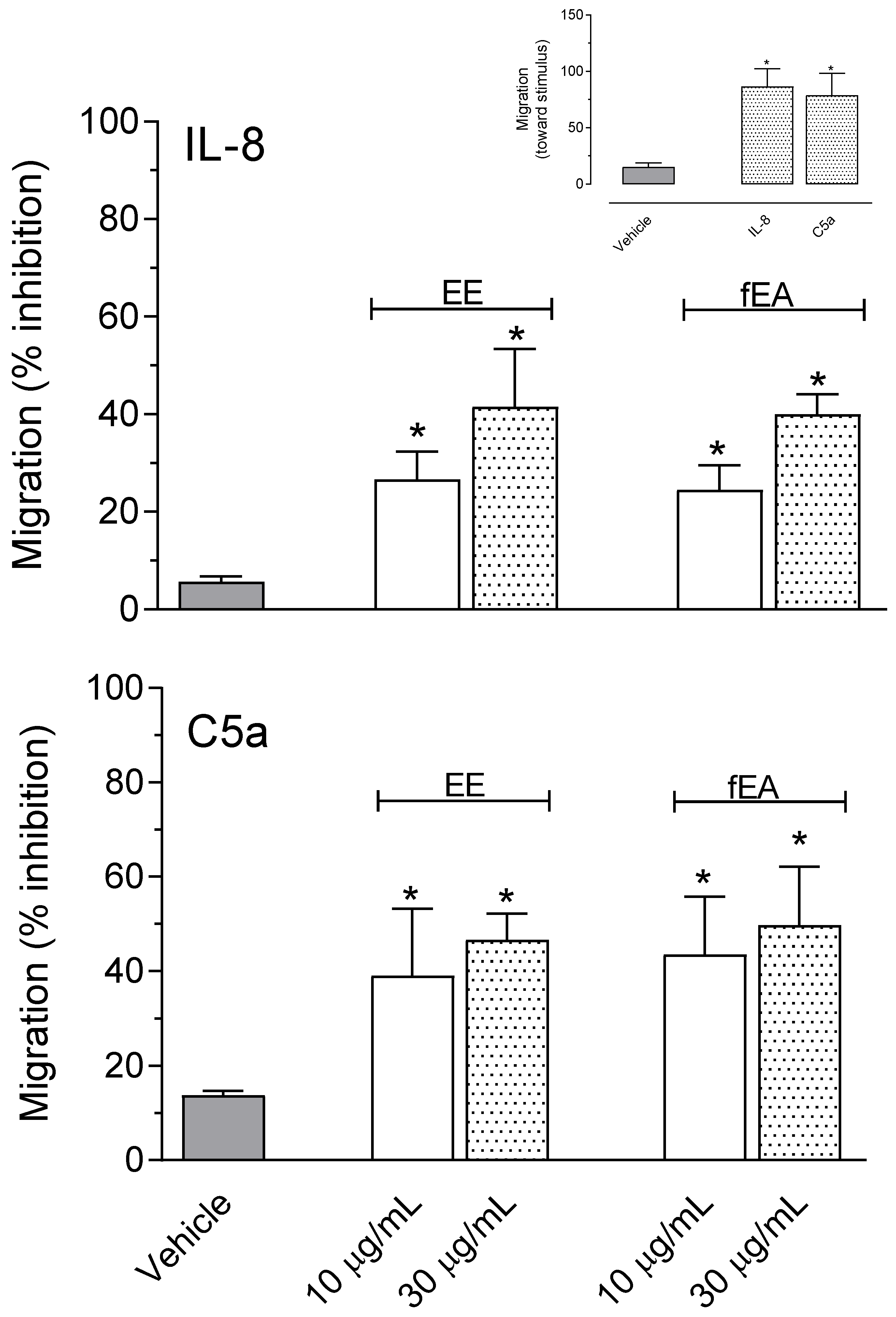
2.6. T. granulosa EE and fEA Decreased Polymorphonuclear Neutrophils (PMNs) Migration Induced by IL-8 and C5a
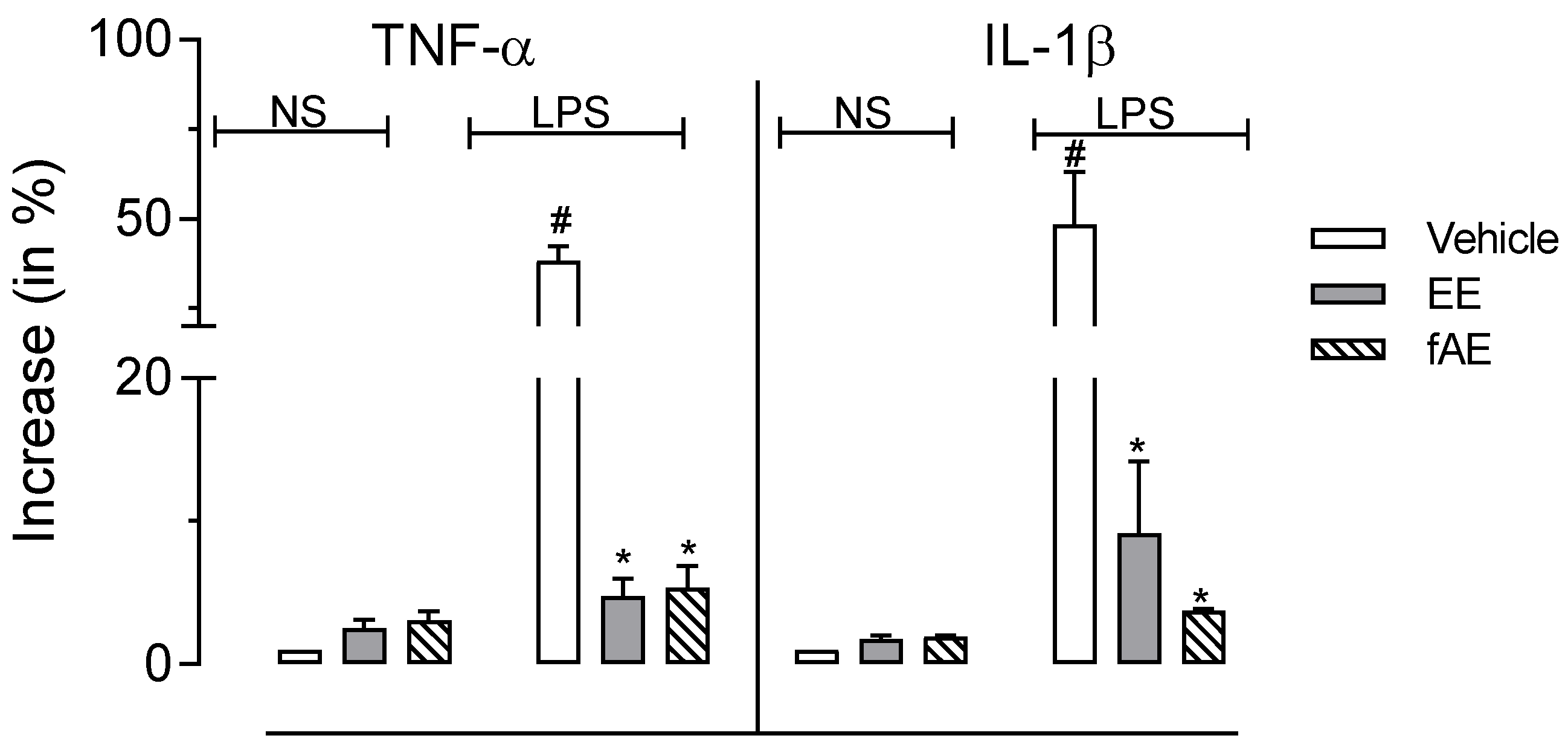
2.7. T. granulosa EE and fEA Inhibited TNF-α and IL-1β Gene Expression
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. Reagents
4.3. Animals
4.4. fEA Chemical Investigation
4.4.1. HPLC-DAD Quantification
4.4.2. Structural Elucidation
4.5. Formalin-Induced Licking Behavior
4.6. Carrageenan-Induced Inflammation into the Subcutaneous Air Pouch (SAP)
4.7. Neutrophils’ Isolation and Stimulation
Cell Viability—Annexin V/Propidium Iodide Staining
4.8. Cytokines (IL-1β, TNF-α, IFN-γ), Nitric Oxide (NO), and Proteins Quantifications
4.9. RNA Isolation
cDNA Reverse Transcription and Real-Time PCR
4.10. Neutrophil Chemotaxis Evaluation
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Valli, M.; Russo, H.M.; Bolzani, V.S. The potential contribution of the natural products from Brazilian biodiversity to bioeconomy. An. Acad. Bras. Ciências 2018, 90, 763–778. [Google Scholar] [CrossRef] [Green Version]
- Koparde, A.A.; Doijad, R.C.; Magdum, C.S. Natural Products in Drug Discovery. In Pharmacognosy-Medicinal Plants; Perveen, S., Ed.; BoD–Books on Demand: London, UK, 2019. [Google Scholar]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Brennan, E.; Kantharidis, P.; Cooper, M.E.; Godson, C. Pro-resolving lipid mediators: Regulators of inflammation, metabolism and kidney function. Nat. Rev. Nephrol. 2021, 17, 725–739. [Google Scholar] [CrossRef]
- Fullerton, J.N.; Gilroy, D.W. Resolution of inflammation: A new therapeutic frontier. Nat. Rev. Drug Discov. 2016, 15, 551–567. [Google Scholar] [CrossRef]
- Varela, M.L.; Mogildea, M.; Moreno, I.; Lopes, A. Acute Inflammation and Metabolism. Inflammation 2018, 41, 1115–1127. [Google Scholar] [CrossRef]
- Moore, N.; Duong, M.; Gulmez, S.E.; Blin, P.; Droz, C. Pharmacoepidemiology of non-steroidal anti-inflammatory drugs. Therapie 2019, 74, 271–277. [Google Scholar] [CrossRef]
- Alice, C.B.; Vargas, V.M.F.; Silva, G.A.A.B.; De Siqueira, N.C.S.; Schapoval, E.E.S.; Gleye, J.; Henriques, J.A.P.; Henriques, A.T. Screening of plants used in south Brazilian folk medicine. J. Ethnopharmacol. 1991, 35, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.; Schadow, B.; Quijano, C.E.; Marx, F. Chemical characterization and antioxidant capacity of berries from Clidemia rubra (Aubl.) Mart. (Melastomataceae). Food Res. Int. 2011, 44, 2120–2127. [Google Scholar] [CrossRef]
- Murugan, R.; Parimelazhagan, T. Study of anti-nociceptive, anti-inflammatory properties and phytochemical profiles of Osbeckia parvifolia Arn. (Melastomataceae). Ind. Crops Prod. 2013, 51, 360–369. [Google Scholar] [CrossRef]
- Jiménez, N.; Carrillo-Hormaza, L.; Pujol, A.; Álzate, F.; Osorio, E.; Lara-Guzman, O. Antioxidant capacity and phenolic content of commonly used anti-inflammatory medicinal plants in Colombia. Ind. Crops Prod. 2015, 70, 272–279. [Google Scholar] [CrossRef]
- Jones, E.; Ekundayo, O.; Kingston, D.G. Plant anticancer agents. XI. 2, 6-Dimethoxybenzoquinone as a cytotoxic con-stituent of Tibouchina pulchra. J. Nat. Prod. 1981, 44, 493–494. [Google Scholar] [CrossRef]
- Santos, F.M.D.; Souza, M.G.D.; Crotti, A.E.M.; Martins, C.H.; Ambrósio, S.R.; Veneziani, R.; Silva, M.L.A.; Cunha, W.R. Evaluation of antimicrobial activity of extracts of Tibouchina can-dolleana (melastomataceae), isolated compounds and semi-synthetic derivatives against en-dodontic bacteria. Braz. J. Microbiol. 2012, 43, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuster, R.M.; Arnold, N.; Wessjohann, L. Anti-fungal flavonoids from Tibouchina grandifolia. Biochem. Syst. Ecol. 2009, 37, 63–65. [Google Scholar] [CrossRef]
- Niño, J.; Espinal, C.M.; Mosquera, O.M.; Correa, Y.M. Antimycotic activity of 20 plants from Colombian flora. Pharm. Biol. 2003, 41, 491–496. [Google Scholar] [CrossRef]
- Singha, U.K.; Guru, P.Y.; Sen, A.B.; Tandon, J.S. Antileishmanial activity of traditional plants against Leishmania donovani in Golden hamsters. Int. J. Pharmacogn. 1992, 30, 289–295. [Google Scholar] [CrossRef]
- Tracanna, M.I.; Fortuna, A.M.; Contreras Cardenas, A.V.; Marr, A.K.; McMaster, W.R.; Gómez-Velasco, A.; Sánchez-Arreola, E.; Hernández, L.R.; Bach, H. Anti-Leishmanial, anti-inflammatory and antimicrobial activities of phenolic derivatives from Tibouchina paratropica. Phytother. Res. 2015, 29, 393–397. [Google Scholar] [CrossRef]
- Dias, Ê.R.; Dias, T.D.L.M.F.; Alexandre-Moreira, M.S.; Branco, A. Antinociceptive activity of Tibouchina pereirae, an endemic plant from the Brazilian semiarid region. Z. Naturforsch. C 2016, 71, 261–265. [Google Scholar] [CrossRef]
- Sobrinho, A.P.; Minho, A.S.; Ferreira, L.L.; Martins, G.R.; Boylan, F.; Fernandes, P.D. Characterization of anti-inflammatory effect and possible mechanism of action of Tibouchina granulosa. J. Pharm. Pharmacol. 2017, 69, 706–713. [Google Scholar] [CrossRef]
- Iwashina, T.; Kamenosono, K.; Ueno, T. Hispidulin and nepetin 4’-glucosides from Cirsium olipophyllum. Phytochemistry 1999, 51, 1109–1111. [Google Scholar] [CrossRef]
- Parada, C.A.; Tambeli, C.H.; Cunha, F.Q.; Ferreira, S.H. The major role of peripheral release of histamine and 5-hydroxytryptamine in formalin-induced nociception. Neuroscience 2001, 102, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.R.; Calixto, J.B. Further evidence for the involvement of tachykinin receptor subtypes in formalin and capsaicin models of pain in mice. Neuropeptides 1997, 31, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Omote, K.; Kawamata, T.; Kawamata, M.; Namiki, A. Formalin-induced release of excitatory amino acids in the skin of the rat hindpaw. Brain Res. 1984, 787, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Milano, J.; Oliveira, S.M.; Rossato, M.F.; Sauzem, P.D.; Machado, P.; Beck, P.; Zanatta, N.; Martins, M.A.; Mello, C.F.; Rubin, M.A.; et al. Antinociceptive effect of novel trihalomethyl-substituted pyrazoline methyl esters in formalin and hot-plate tests in mice. Eur. J. Pharmacol. 2008, 581, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Malmberg, A.B.; Yaksh, T.L. Cyclooxygenase inhibition and the spinal release of prostaglandin E2 and amino acids evoked by paw formalin injection: A microdialysis study in unanesthetized rats. J. Neurosci. 1995, 15, 2768–2776. [Google Scholar] [CrossRef]
- Sedgwick, A.D.; Lees, P. A comparison of air pouch, sponge and pleurisy models of acute carrageenan inflammation in the rat. Agents Actions 1986, 18, 439–446. [Google Scholar] [CrossRef]
- Ferrándiz, M.L.; Gil, B.; Sanz, M.J.; Ubeda, A.; Erazo, S.; González, E.; Negrete, R.; Pacheco, S.; Payáa, M.; Alcaraz, M.J. Effect of bakuchiol on leukocyte functions and some inflammatory responses in mice. J. Pharm. Pharmacol. 1996, 48, 975–980. [Google Scholar] [CrossRef]
- Fröde, T.S.; Ferreira, S.I.A.C.P.; Medeiros, Y.S. Analysis of local and systemic inflammatory responses induced by polymicrobial peritonitis in mice. Mediat. Inflamm. 2001, 10, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Sun, R.; Wei, H.; Tian, Z. Toll-like receptor 3 ligand attenuates LPS-induced liver injury by down-regulation of toll-like receptor 4 expression on macrophages. Proc. Natl. Acad. Sci. USA 2005, 102, 17077–17082. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Liu, J.; Cao, X. Regulation of type I interferon signaling in immunity and inflammation: A comprehensive review. J. Autoimmun. 2017, 83, 1–11. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as cytokine mediators: A possible therapy for Inflammation related diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Min, K.J.; Kim, M.J.; Kim, S.H.; Kwon, T.K. Hispidulin inhibits mast cell-mediated allergic inflammation through down-regulation of histamine release and inflammatory cytokines. Molecules 2019, 24, 2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunskaar, S.; Fasmer, O.B.; Hole, K. Formalin test in mice, a useful technique for evaluating mild analgesia. J. Neurosci. Methods 1985, 14, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.M.; Rezende, C.M.; Fontes, S.P.; Matheus, M.E.; Fernandes, P.D. Antinociceptive activity of Amazonian Copaiba oils. J. Ethnopharmacol. 2007, 109, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, L.J.; Guilhon, C.C.; Alviano, D.S.; Matheus, M.E.; Antoniolli, A.R.; Cavalcanti, S.C.; Alves, P.B.; Alviano, C.S.; Fernandes, P.D. Characterization of the anti-inflammatory and antinociceptive ac-tivities of the Hyptis pectinata (L.) Poit essential oil. J. Ethnopharmacol. 2011, 134, 725–732. [Google Scholar] [CrossRef]
- Pouliot, M.; Fiset, M.É.; Massé, M.; Naccache, P.H.; Borgeat, P. Adenosine up-regulates cyclooxygenase-2 in human granulocytes: Impact on the balance of eicosanoid generation. J. Immunol. 2002, 169, 5279–5286. [Google Scholar] [CrossRef] [Green Version]
- Pietkiewicz, S.; Schmidt, J.H.; Lavrik, I.N. Quantification of apoptosis and necroptosis at the single cell level by a combination of imaging flow cytometry with classical Annexin V/propidium iodide staining. J. Immunol. Methods 2015, 423, 99–103. [Google Scholar] [CrossRef]
- Bartholomew, B. A rapid method for the assay of nitrate in urine using the nitrate reductase enzyme of Escherichia coli. Food Chem. Toxicol. 1984, 22, 541–543. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [5N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Dussault, A.A.; Pouliot, M. Rapid and simple comparison of messenger RNA levels using real-time PCR. Biol. Proced. Online 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Mills, S.C.; Goh, P.H.; Kudatsih, J.; Ncube, S.; Gurung, R.; Maxwell, W.; Mueller, A. Cell migration towards CXCL12 in leukemic cells compared to breast cancer cells. Cell. Signal. 2016, 28, 316–324. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
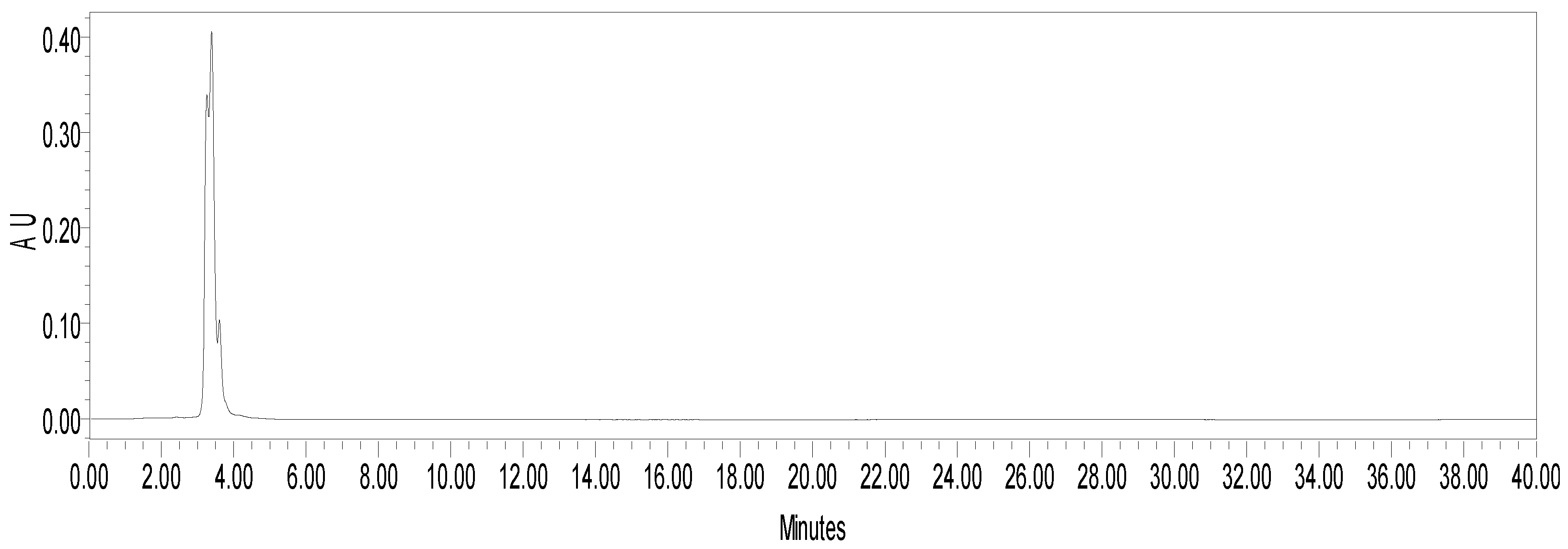{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence | |
|---|---|---|
| ACTB | F | 5′-CAAATGCTTCTAGGTGGACT-3′ |
| R | 5′-GCTGTCACCTTCACCGTTC-3′ | |
| TNF | F | 5′-AGCCATGTTGTAGCAAACC-3′ |
| R | 5′-TGAGGTACAGGCCCTCTGAT-3′ | |
| IL-1β | F | 5′-GGACAAGCTGAGGAAGATGC-3′ |
| R | 5′-TCGTTATCCCATGTGTCGAA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guilhon, C.C.; Minho, A.S.; Pouliot, M.; Boylan, F.; Fernandes, P.D. Tibouchina granulosa Leaves Present Anti-Inflammatory Effect. Pharmaceuticals 2022, 15, 1458. https://doi.org/10.3390/ph15121458
Guilhon CC, Minho AS, Pouliot M, Boylan F, Fernandes PD. Tibouchina granulosa Leaves Present Anti-Inflammatory Effect. Pharmaceuticals. 2022; 15(12):1458. https://doi.org/10.3390/ph15121458
Chicago/Turabian StyleGuilhon, Carolina Carvalho, Alan Silva Minho, Marc Pouliot, Fabio Boylan, and Patricia Dias Fernandes. 2022. "Tibouchina granulosa Leaves Present Anti-Inflammatory Effect" Pharmaceuticals 15, no. 12: 1458. https://doi.org/10.3390/ph15121458
APA StyleGuilhon, C. C., Minho, A. S., Pouliot, M., Boylan, F., & Fernandes, P. D. (2022). Tibouchina granulosa Leaves Present Anti-Inflammatory Effect. Pharmaceuticals, 15(12), 1458. https://doi.org/10.3390/ph15121458