
Component Characterization, In Vitro Activities and Molecular Mechanism of Cydonia oblonga Mill. against Diabetic

 and
and
Abstract
:1. Introduction
2. Results
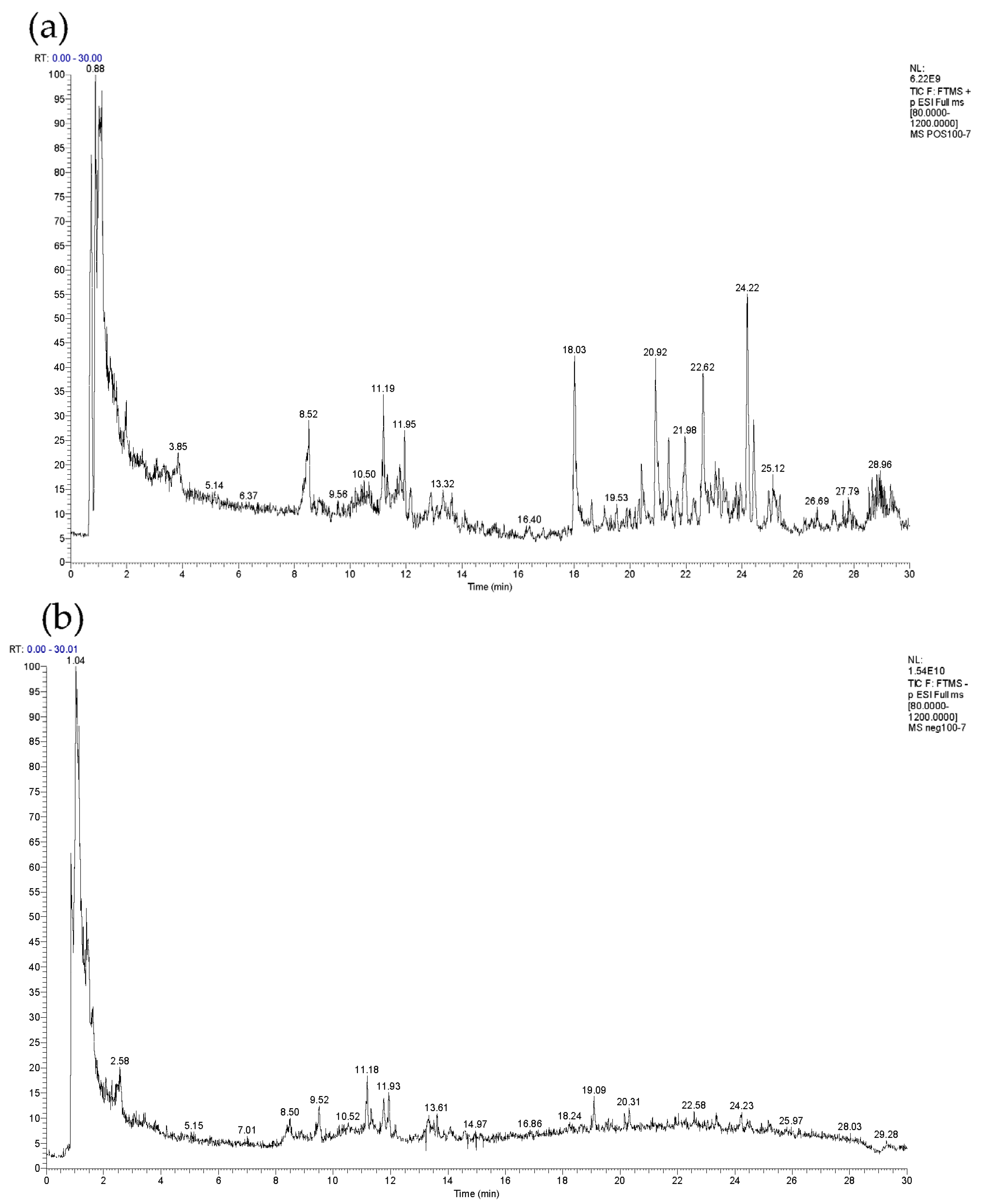
2.1. Identification of COMF Components
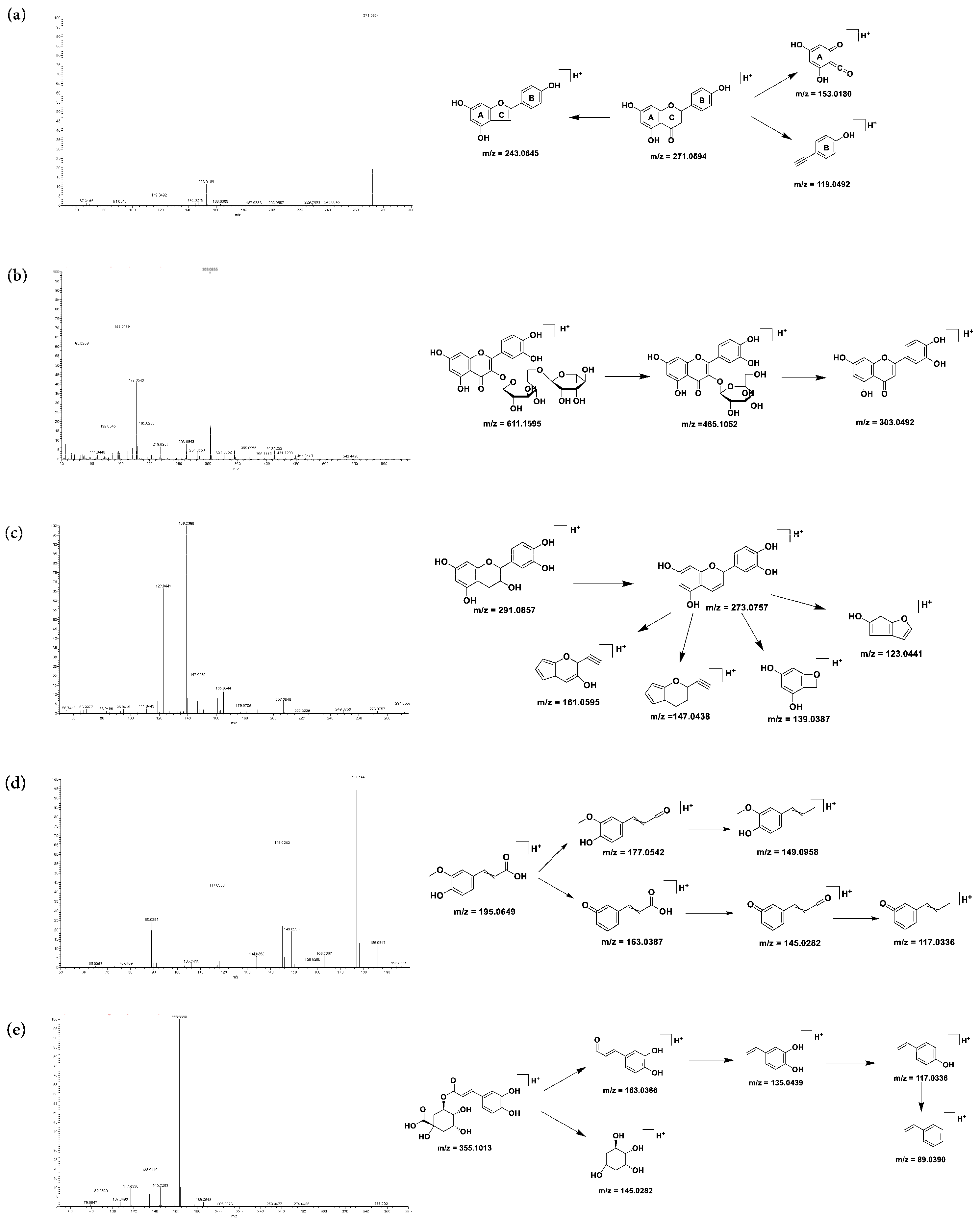
2.1.1. Fragmentation Pattern of Flavonoid Compounds
2.1.2. Organic Acid and Polyphenol Compounds
2.1.3. Other Compounds
2.2. In Vitro Activity Assay
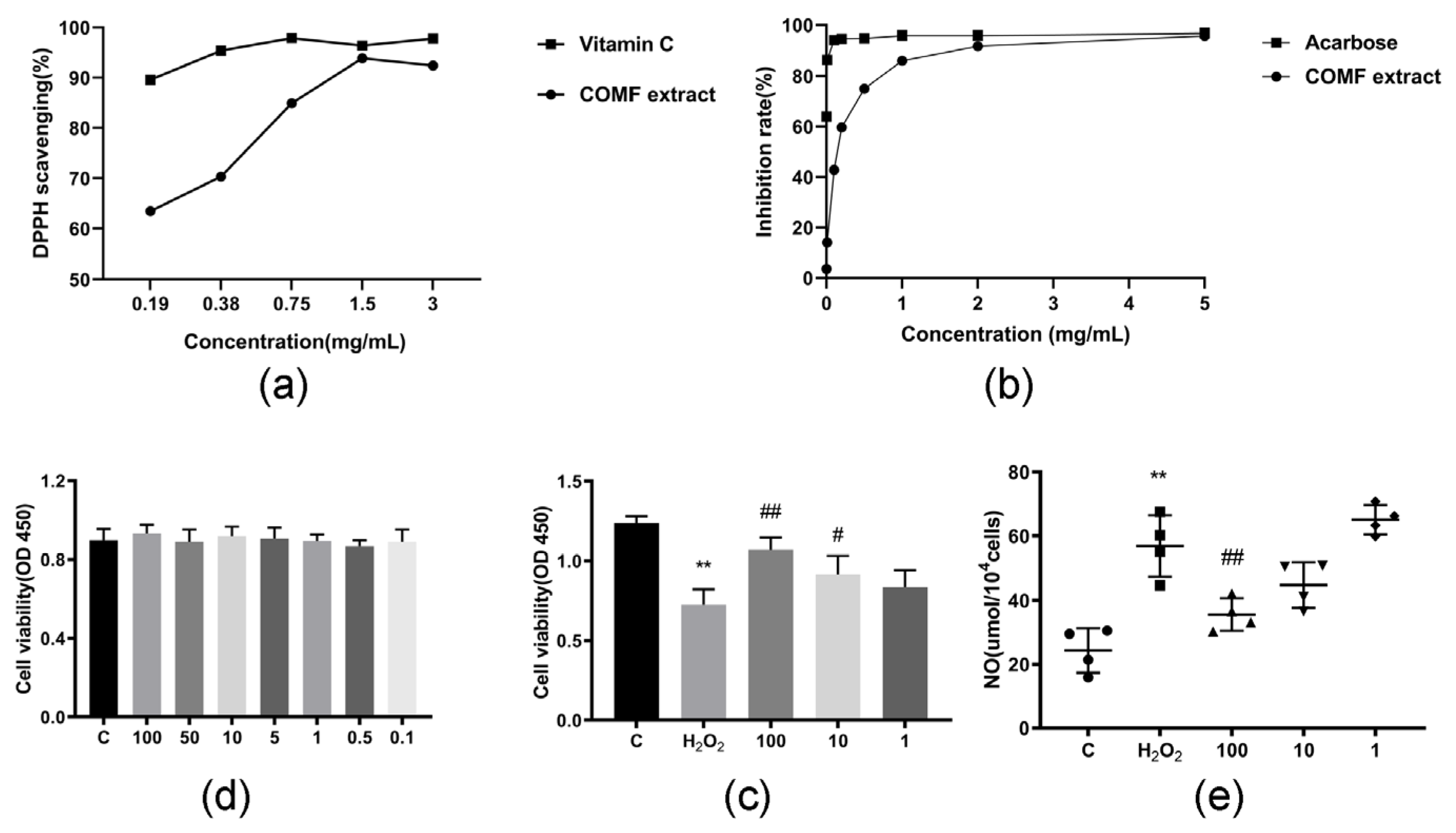
2.2.1. DPPH Free Radical Scavenging Ability of COMF Extract
2.2.2. α-Glucosidase Inhibitory Potency
2.2.3. Effect of COMF Extracts on HUVECs Proliferation
2.2.4. Cytoprotective Effect of Extracts in H2O2-Induced Oxidative Stress
2.2.5. The Extracts Effects on NO Production
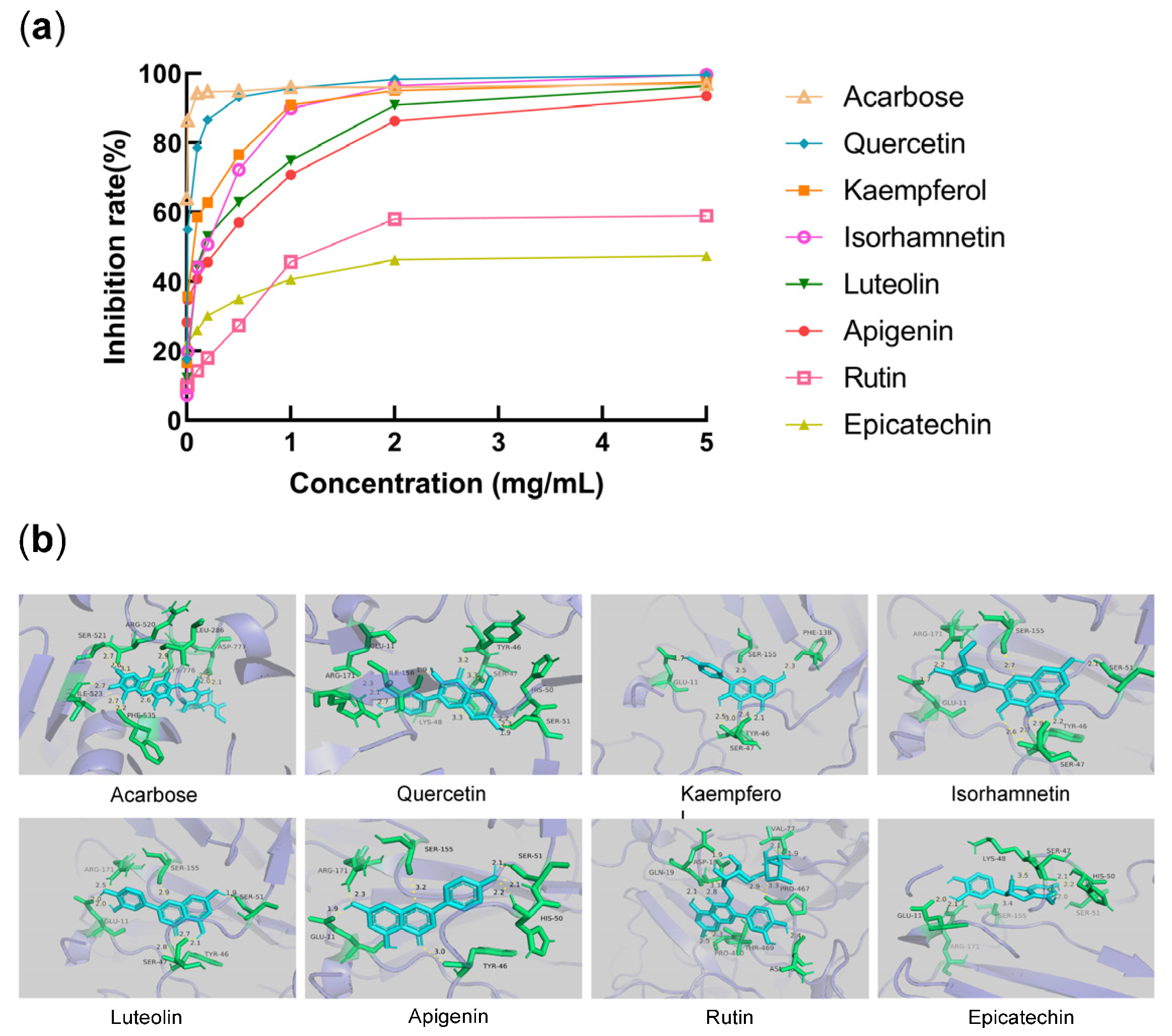
2.3. Selection of Active Monomers
2.4. In Vitro Activity Assay Result and Validation of Molecular Mechanisms
2.4.1. α-Glucosidase Inhibitory Potency
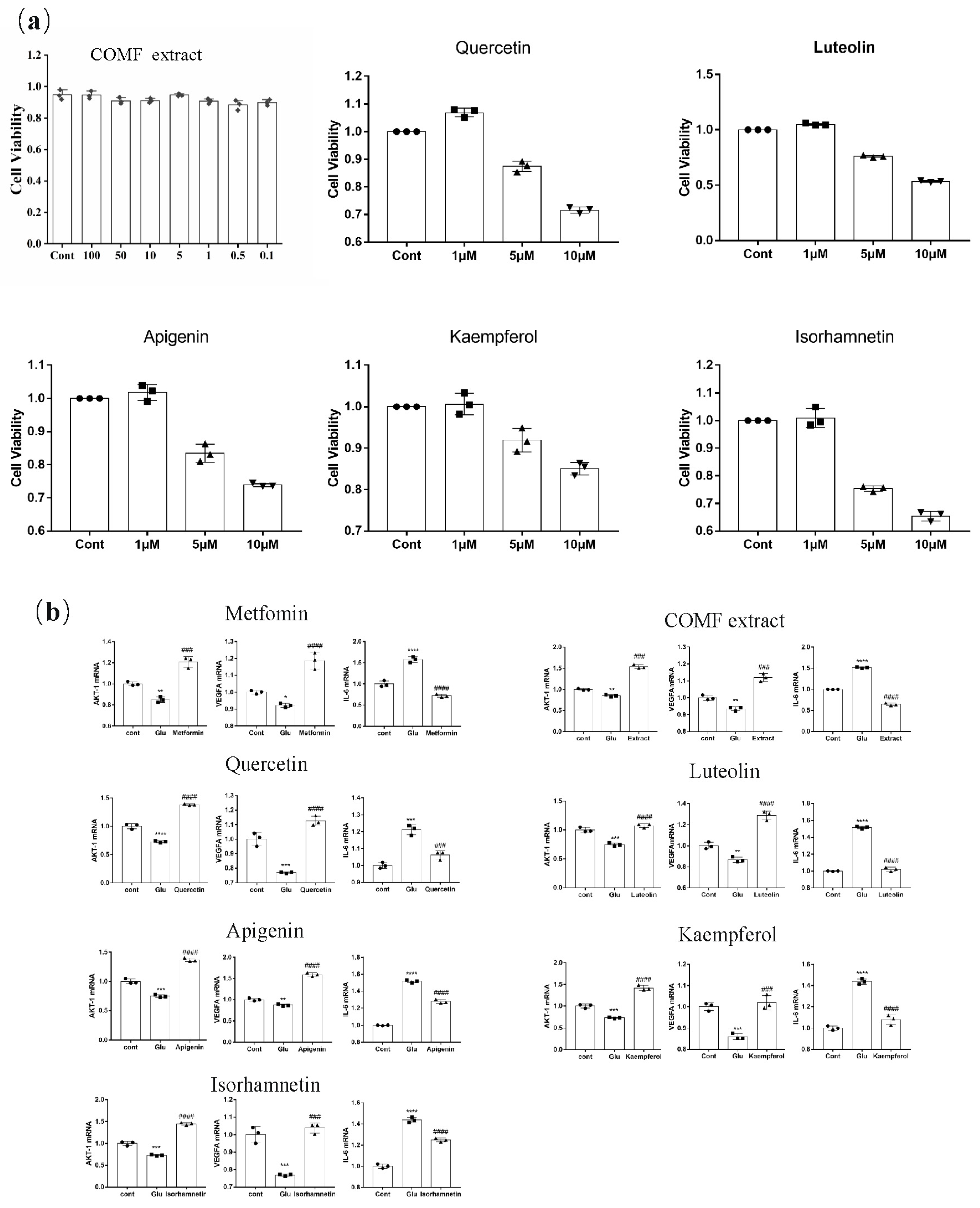
2.4.2. Effect of COMF Extracts and Active Compounds on HUVECs Proliferation
2.4.3. Expression of AKT1, IL-6 and VEGFA mRNA
3. Materials and Methods
3.1. Sample Preparation
3.2. UPLC-MS/MS Analysis
3.3. In Vitro Activity Assay
3.3.1. Determination of DPPH Clearance Rate
3.3.2. Determination of α-Glucosidase Inhibition Rate
3.3.3. Cell Proliferation Assay
3.3.4. Detection of Antioxidant Stress
3.3.5. Detection of Cellular NO
3.4. Prediction of Active Ingredients
3.5. α-Glucosidase Inhibition Activity and Molecular Docking Analysis of Representative Flavonoids
3.6. Validation of the Molecular Mechanism
3.6.1. Cell Proliferation Assay and Establishment of Cellular Glucose Damage Model
3.6.2. Expression of AKT1, IL-6 and VEGFA mRNA by RT-qPCR
3.7. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ogurtsova, K.; Guariguata, L.; Barengo, N.C.; Ruiz, P.L.; Sacre, J.W.; Karuranga, S.; Sun, H.; Boyko, E.J.; Magliano, D.J. IDF Diabetes Atlas: Global estimates of undiagnosed diabetes in adults for 2021. Diabetes Res. Clin. Pract. 2022, 183, 109118. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Fang, S. Therapeutic Strategies for Diabetes: Immune Modulation in Pancreatic β Cells. Front. Endocrinol. 2021, 12, 716692. [Google Scholar] [CrossRef]
- Katome, T.; Obata, T.; Matsushima, R. Use of RNA interference-mediated gene silencing and adenoviral overexpression to elucidate the roles of AKT/protein kinase B isoforms in insulin actions. J. Biol. Chem. 2003, 278, 28312–28323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, T.; Kubota, N.; Kadowaki, T. Imbalanced insulin actions in obesity and type 2 diabetes: Key mouse models of insulin signaling pathway. Cell Metab. 2017, 25, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Huilin, L.; Hengxia, Z.; Miao, S.; Deliang, L. Effects of hydroxysafflor yellow A on the PI3K/AKT pathway and apoptosis of pancreatic β-Cells in type 2 diabetes mellitus rats. Diabet Metab. Synd. OB 2020, 13, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Dan, T. Cydonia oblonga Mill. seed extracts enhance glucose consumption by activating PI3K/AKT insulin signaling pathway in L6 myotubes. In Proceedings of the 2015 Workshop 6, Suzhou, China, 12 June 2015. [Google Scholar]
- El-Nashar, H.A.S.; Mostafa, N.M.; El-Shazly, M.; Eldahshan, O.A. The Role of Plant-Derived Compounds in Managing Diabetes Mellitus: A Review of Literature from 2014 to 2019. Curr. Med. Chem. 2021, 28, 4694–4730. [Google Scholar] [CrossRef]
- Abdelghffar, E.; Mostafa, N.M.; El-Nashar, H.A.S.; Eldahshan, O.A.; Singab, A.N. Chilean pepper (Schinus polygamus) ameliorates the adverse effects of hyperglycaemia/dyslipidaemia in high fat diet/streptozotocin-induced type 2 diabetic rat model. Ind. Crops Prod. 2022, 183, 114953. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Valentão, P.; Ferreres, F.; Seabra, R.M.; Ferreira, M.A. Quince (Cydonia oblonga Miller) fruit (pulp, peel, and seed) and jam: Antioxidant activity. J. Agric. Food Chem. 2004, 52, 4705–4712. [Google Scholar] [CrossRef]
- Isabel, G.; Galindo, A.; Hernández, P.R.; Morales, D.; Hernandez, F. Volatile composition and sensory and quality attributes of quince (Cydonia oblonga Mill.) fruits as affected by water stress. Sci. Hortic. 2019, 244, 68–74. [Google Scholar] [CrossRef]
- Carbonell-Barrachina, A.A.; Szychowski, P.J.; Vásquez, M.V.; Hernández, F.; Wojdyło, A. Technological aspects as the main impact on quality of quince liquors. Food Chem. 2015, 167, 387–395. [Google Scholar] [CrossRef]
- Blanda, G.; Rodriguez-Roque, M.J.; Comandini, P.; Flores-Cordova, M.A.; Soto-Caballero, M.C. Phenolic profile and physicochemical characterization of quince (Cydonia oblonga Mill) fruits at different maturity index. Not. Bot Horti. Agrobot. Cluj-Napoca 2020, 48, 2306–2315. [Google Scholar] [CrossRef]
- Zhang, L.; Rocchetti, G.; Zengin, G.; Ak, G.; Lucini, L. The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential. Foods 2021, 10, 1230. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, A.B.; Strugała, P.; Dudra, A.; Kucharska, A.Z.; Sokół-Łętowska, A.; Wojnicz, D.; Cisowska, A.; Sroka, Z.; Gabrielska, J. Microbiological, antioxidant and lipoxygenase-1 inhibitory activities of fruit extracts of chosen Rosaceae family species. Adv. Clin. Exp. Med. 2020, 29, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaida, B.; Singh, N.B.; Singh, K. In-Vitro Evaluation of Anti-Inflammatory and Anti-Microbial Properties of Ethanolic Extract of Cydonia Oblonga Seeds. J. Sci. Ind. Res. 2020, 79, 4. [Google Scholar]
- Abliz, A.; Aji, Q.; Abdusalam, E.; Sun, X.; Abdurahman, A.; Zhou, W.; Moore, N.; Umar, A. Effect of Cydonia oblonga Mill. leaf extract on serum lipids and liver function in a rat model of hyperlipidaemia. J. Ethnopharmacol. 2014, 151, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.Y.; Zhou, H.L.; Zhou, Q.; Sun, L.; Jiang, H.Q.; Zhang, S.M.; Guo, F.X.; Hou, L.J.; Zhang, C.S.; Dong, S.H. Analysis on chemical constituents of fruit of Chaenomels speciosa UPLC-Q-Exactive Orbitrap-MS. Chin. Herb. Med. 2018, 49, 4773–4779. [Google Scholar]
- Simona, P.; Giuseppina, C.; Paola, N.; Francesca, P.; Lorenzo, M.; Severina, P. Ultrasound-assisted aqueous extraction, LC-MS/MS analysis and radiomodulating capability of autochthonous Italian sweet cherry fruits. Food Funct. 2018, 9, 1840–1849. [Google Scholar] [CrossRef]
- Yang, L.M.; Yang, L.J.; Jia, P.; Wei, L.; Zhang, Y.J.; Wang, S.X.; Zhang, P.; Zheng, X.H. HPLC-Q-TOF-MS/MS-based analysis of chemical constituents in Choerospondiatis fructus. Acad. J. Second Mil Med. Univ. 2016, 37, 159–166. [Google Scholar] [CrossRef]
- Momoka, S.; Hiroto, K.; Yasunori, N. Determination of Organic Acids in Honey by Liquid Chromatography with Tandem Mass Spectrometry. Food Anal Methods 2020, 13, 1–9. [Google Scholar] [CrossRef]
- Yuan, H.; Xiaoyan, L.; Zilin, C. Determination of bioactive components in the fruits of Cercis chinensis Bunge by HPLC-MS/MS and quality evaluation by principal components and hierarchical cluster analyses. J. Pharm. Anal. 2021, 11, 465–471. [Google Scholar]
- Xu, W.; Fu, Z.; Lin, J.; Huang, X.; Chen, D.; Yu, H.; Huang, Z.; Fan, S. Qualitative and quantitative analysis of major components in Tetrastigma hemsleyanum by HPLC-Q-TOF-MS and UPLC-Qq Q-MS. China J. Chin. Mater. Med. 2014, 39, 4365–4372. [Google Scholar]
- Baohong, L.; Jindong, W.; Xianglu, L. LC-MS/MS determination and pharmacokinetic study of bergenin, the main bioactive component of Bergenia purpurascens after oral administration in rats. Planta Med. 2013, 3, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yu, L.; Ma, F.; Li, P. Quantification of phenolic compounds in vegetable oils by mixed-mode solid-phase extraction isotope chemical labeling coupled with UHPLC-MS/MS. Food Chem. 2021, 334, 127572. [Google Scholar] [CrossRef] [PubMed]
- Qinghan, G.; Chunsen, W.; Min, W. The jujube (Ziziphus jujuba Mill.) fruit: A review of current knowledge of fruit composition and health benefits. J. Agric. Food Chem. 2013, 61, 3351–3363. [Google Scholar] [CrossRef]
- Yahui, Y.; Tao, W.; Ju, C.; Abudula, R.; Lei, H.; Isa, A. Identification and Attribution of Chemical Constituents in Naizilai Granules by UHPLC-Q-Orbitrap-MS. Chin. J. Exper. Tradit. Med. Form. 2021, 27, 156–166. [Google Scholar] [CrossRef]
- Antal, D.S.; Schwaiger, S.; Ellmerer-Müller, E.P.; Stuppner, H. Cotinus coggygria wood: Novel flavanone dimer and development of an HPLC/UV/MS method for the simultaneous determination of fourteen phenolic constituents. Planta Med. 2010, 76, 1765–1772. [Google Scholar] [CrossRef] [Green Version]
- Reddy, Y.M.; Kumar, S.P.J.; Saritha, K.V.; Gopal, P.; Reddy, T.M.; Simalgandara, J. Phytochemical Profiling of Methanolic Fruit Extract of Gardenia latifolia Ait. by LC-MS/MS Analysis and Evaluation of Its Antioxidant and Antimicrobial Activity. Plants 2021, 10, 545. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Jia, Z.; Han, T.; Liu, M.; Jia, T.; Qu, W.; Xu, X.; Li, X. UPLC-MS/MS Determination of Chlorogenic Acid, Hyperoside and Astragalin in Plasma and its Pharmacokinetic Application in Liver Injury Rats. Curr. Pharm. Anal. 2021, 17, 1016–1025. [Google Scholar] [CrossRef]
- Wei, L.; Yulin, F.; Tianer, T.; Huan, W.; Yan, L.; Guoyue, Z.; Bei, W.; Mingzhen, H. Rapid Analysis on Flavonoids in Glechoma longituba (Nakai) Kupr by UPLC-Q-TOF/MS Couple with Diagnostic Ions. J. Chinese Mass Spectrom. Soc. 2016, 37, 504–516. [Google Scholar] [CrossRef]
- Ghallab, D.S.; Mohyeldin, M.M.; Shawky, E.; Metwally, A.M.; Ibrahim, R.S. Chemical profiling of Egyptian propolis and determination of its xanthine oxidase inhibitory properties using UPLC-MS/MS and chemometrics. LWT Food Sci. Technol. 2021, 136, 110298. [Google Scholar] [CrossRef]
- Huang, H.; Kang, J.; Yu, Y.; Zhao, P.; Feng, S.; Du, Y.; Li, J. Identification of Chemical Constituents of Bufei Yishen Formula by UPLC-Q-Orbitrap MS. J. Instrum. Anal. 2019, 38, 1–13. [Google Scholar]
- Yan, W.; Yan, L.; Zhang, X.; Yan, X.; Huang, S. UHPLC-ESI-MS/MS analysis and identification of the Artemisia sphaeroce Krasch. Chin. Sci. 2015, 10, 2127–2129+2168. [Google Scholar]
- Jiang, J.; Xun, A.; Li, Y.; Duan, F.; Zeng, Z.; Sun, D. Analysis of constituents from Saururi Herba by UHPLC-Q-TOF-MS. J. Guangdong Pharm. Univ. 2019, 35, 746–751. [Google Scholar] [CrossRef]
- Zou, Y.; Yan, H.Y. Research progress on chemical constituents and pharmacological activities of Chinese traditional medicine papaya. Int. J. Pharm. 2019, 46, 507–515. [Google Scholar] [CrossRef]
- Zhang, L.; Tai, Y.L.; Wang, Y.J.; Meng, Q.; Wan, X. The proposed biosynthesis of procyanidins by the comparative chemical analysis of five Camellia species using LC-MS. Sci. Rep. 2017, 7, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, D.; Zhang, C.; Yang, W.; Li, C.; Gao, Z.; Pei, K.; Li, Y. Piezo1 mediates endothelial atherogenic inflammatory responses via regulation of YAP/TAZ activation. Hum. Cell 2022, 35, 51–62. [Google Scholar] [CrossRef]
- Yan, X.C.; Cao, J.; Liang, L.; Wang, L.; Gao, F.; Yang, Z.Y.; Duan, J.L.; Chang, T.F.; Deng, S.M.; Liu, Y.; et al. miR-342-5p Is a Notch Downstream Molecule and Regulates Multiple Angiogenic Pathways Including Notch, Vascular Endothelial Growth Factor and Transforming Growth Factor β Signaling. J. Am. Heart Assoc. 2016, 5, e003042. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Rocha, K.M.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Larrosa-Pérez, M.; Moreno-Jiménez, M.R. Phenolic Acids and Flavonoids in Acetonic Extract from Quince (Cydonia oblonga Mill.): Nutraceuticals with Antioxidant and Anti-Inflammatory Potential. Molecules 2022, 27, 2462. [Google Scholar] [CrossRef]
- Patar, P.; Abdureheman, A.D.; Tursun, N.; Salamu, Q.; Hao, Q.; Shunjie, Y.; Yimin, O.; Wumaier, A. The effect of total flavonoid of Cydonia oblonga Mill. leaf on blood sugar, liver function and lipid metabolism disorders. J. Xinjiang Med. Univ. 2016, 39, 835–840. [Google Scholar]
- He, Y.; Deng, X.; He, Y.; Wang, Z.; Wang, C.; Wu, R. Study on fragmentation patterns of five prenlated flavonols by applying electrospray ionization mass spectrometry based on UHPLC-Orbitrap MS. J. Tradit. Chin. Med. 2021, 36, 5595–5598. [Google Scholar]
- Lee, M.S.; Chyau, C.C.; Wang, C.P.; Wang, T.H.; Chen, J.H.; Lin, H.H. Flavonoids Identification and Pancreatic Beta-Cell Protective Effect of Lotus Seedpod. Antioxidants 2020, 9, 658. [Google Scholar] [CrossRef]
- Qin, C.; Xia, T.; Li, G.; Zou, Y.; Cheng, Z.; Wang, Q. Hawthorne leaf flavonoids prevent oxidative stress injury of renal tissues in rats with diabetic kidney disease by regulating the p38 MAPK signaling pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 3440–3446. [Google Scholar]
- Feng, X.; Duan, H.; Li, S. Protective role of Pollen Typhae total flavone against the palmitic acid-induced impairment of glucose-stimulated insulin secretion involving GPR40 signaling in INS-1 cells. Int. J. Mol. Med. 2017, 40, 922–930. [Google Scholar] [CrossRef] [Green Version]
- Dhanya, R.; Kartha, C.C. Quercetin improves oxidative stress-induced pancreatic beta cell alterations via mTOR-signaling. Mol. Cell. Biochem. 2021, 476, 3879–3887. [Google Scholar] [CrossRef]
- Arya, D.V.; Alik, S.M.; Suchal, K.; Bhatia, J. A18239 Apigenin ameliorates streptozotocin induced diabetic nephropathy in rats by modulation of oxidative stress, apoptosis and inflammation through MAPK pathway. J. Hypertens. 2018, 36, e63. [Google Scholar] [CrossRef]
- Krueger, E.; Austin, M.; Ekpo, I.; Beales, J.; Tessem, J.S. Screening Phytochemicals for Alternative Diabetes Treatments. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Alwhaibi, A.; Verma, A.; Adil, M.S.; Somanath, P.R. The unconventional role of Akt1 in the advanced cancers and in diabetes-promoted carcinogenesis. Pharmacol. Res. 2019, 145, 104270. [Google Scholar] [CrossRef]
- Cai, Y.; Wu, L.; Lin, X.; Hu, X.; Wang, L. Phenolic profiles and screening of potential α-glucosidase inhibitors from Polygonum aviculare L. leaves using ultra-filtration combined with HPLC-ESI-qTOF-MS/MS and molecular docking analysis. Ind. Crops Prod. 2019, 129, 156–168. [Google Scholar] [CrossRef]
- Okuyama, T.; Shirakawa, J.; Yanagisawa, H.; Kyohara, M.; Yamazaki, S.; Tajima, K.; Togashi, Y.; Terauchi, Y. Identification of the matricellular protein Fibulin-5 as a target molecule of glucokinase-mediated calcineurin/NFAT signaling in pancreatic islets. Sci. Rep. 2017, 7, 2364. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, H.; Schmidt, A.M. AGE/RAGE produces endothelial dysfunction in coronary arterioles in Type 2 diabetic mice. Am. J. Physiol. Heart C 2008, 295, H491–H498. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Y.; Zhou, Q.L.; Coleman, K.A.; Chouinard, M.; Boese, Q.; Czech, M.P. Insulin signaling through Akt/protein kinase B analyzed by small interfering RNA-mediated gene silencing. Proc. Natl. Acad. Sci. USA 2003, 100, 7569–7574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, A.K.; Heimberg, H.; Heremans, Y. A comprehensive analysis of cytokine-induced and nuclear factor-kappa B-dependent genes in primary rat pancreatic beta-cells. J. Biol. Chem. 2001, 276, 48879–48886. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR/ Min | Precursor Ions | Detected/ m/z | Calculated/ m/z | Error/ppm | Molecular Formula | Fragment Ions/ m/z | Name | Class | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.88 | [M + H]+ | 175.1187 | 175.1190 | −1.612 | C6H14N4O2 | 175.1187, 116.0707, 71.0691, 70.0658, 60.0564 | Arginine | amino acid | [17] |
| 2 | 1.12 | [M + H]+ | 183.0860 | 183.0863 | −1.719 | C6H14O6 | 183.0860, 165.0546, 147.0648, 129.0546, 111.0442, 69.0341 | Hexitol | other compound | [18] |
| 3 | 1.15 | [M + H]+ | 193.0704 | 193.0707 | −1.215 | C7H12O6 | 157.0494, 147.0649, 129.0546 | Quinic acid | organic acid | [17] |
| 4 | 1.43 | [M + H]+ | 135.0286 | 135.0288 | −1.184 | C4H6O5 | 135.0286, 107.0493, 89.0390 | Malic acid | organic acid | [19] |
| 5 | 1.56 | [M + H]+ | 118.0863 | 118.0863 | 0.803 | C5H11NO2 | 118.0863, 72.0814, 59.0737, 58.0659 | Valine | amino acid | [17] |
| 6 | 1.75 | [M − H]− | 117.0179 | 117.0182 | −2.950 | C4H6O4 | 117.0179, 99.0072, 73.0279, 55.0173 | Succinic Acid | organic acid | [17] |
| 7 | 2.57 | [M + H]+ | 175.0234 | 175.0237 | −1.911 | C6H6O6 | 175.0234, 139.0023, 67.0549 | Shikimic acid | organic acid | [17] |
| 8 | 2.58 | [M − H]− | 191.0190 | 191.0186 | −3.904 | C6H8O7 | 191.0190, 173.0085, 129.0180, 111.0073 | Citric acid | organic acid | [20] |
| 9 | 3.07 | [M + H]+ | 132.1017 | 132.1019 | −1.326 | C6H13NO2 | 132.1017, 86.0969, 69.0705, 57.0343 | Leucine | amino acid | [17] |
| 10 | 3.85 | [M + H]+ | 182.0808 | 182.0812 | −1.866 | C9H11NO3 | 182.0808, 165.0543, 147.0438, 136.0754, 123.0440 | Tyrosine | amino acid | [19] |
| 11 | 6.26 | [M − H]− | 169.0133 | 169.0131 | 0.948 | C7H6O5 | 169.0133, 125.0230 | Gallic acid | polyphenol | None 1 |
| 12 | 8.46 | [M + H]+ | 331.0675 | 331.0660 | 1.420 | C13H16O10 | 331.0675, 169.0132, 125.0230 | Glucogallin | polyphenol | [21] |
| 13 | 8.52 | [M + H]+ | 166.0860 | 166.0863 | −1.837 | C9H11NO2 | 166.0860, 120.0808, 103.0545, 91.0546 | Phenylalanine | amino acid | [17] |
| 14 | 9.48 | [M − H]− | 153.0182 | 153.0182 | −0.099 | C7H6O4 | 153.0182, 109.0280 | Protocatechuic acid | polyphenol | None 1 |
| 15 | 9.91 | [M + H]+ | 579.1486 | 579.1497 | −1.973 | C30H26O12 | 427.1006, 409.0909, 289.0698, 127.0388 | Procyanidin B1 | flavonoid | [22] |
| 16 | 10.50 | [M + H]+ | 355.1014 | 355.1024 | −2.587 | C16H18O9 | 163.0386, 145.0281, 135.0438, 117.0335, 89.0389 | neochlorogenic acid | polyphenol | None 1 |
| 17 | 10.67 | [M − H]− | 271.0829 | 271.0812 | 2.080 | C12H16O7 | 271.0829, 109.0280, 101.0226, 71.0122 | Arbutin | other compound | [23] |
| 18 | 10.74 | [M + H]+ | 579.1485 | 579.1497 | −2.077 | C30H26O12 | 427.1020, 409.0908, 289.0701, 127.0388 | Procyanidin B2 | flavonoid | [22] |
| 19 | 10.79 | [M − H]− | 137.0232 | 137.0233 | −0.953 | C7H6O3 | 137.0232, 108.0201 | 3,4-Dihydroxybenzaldehyde | polyphenol | None 1 |
| 20 | 10.81 | [M + H]+ | 139.0387 | 139.0390 | −1.731 | C7H6O3 | 139.0387, 111.0442, 95.0493, 65.0393 | 3-Hydroxybenzoic acid | organic acid | [24] |
| 21 | 11.19 | [M + H]+ | 355.1013 | 355.1024 | −2.840 | C16H18O9 | 163.0386, 145.0282, 135.0439, 117.0336, 89.0390 | Chlorogenic acid | polyphenol | None 1 |
| 22 | 11.34 | [M + H]+ | 291.0855 | 291.0863 | −2.867 | C15H14O6 | 291.0855, 273.0735, 161.0594, 147.0437, 139.0387, 123.0441 | epicatechin | flavonoid | [22] |
| 23 | 11.52 | [M + H]+ | 169.0492 | 169.0495 | −1.924 | C8H8O4 | 169.0492, 151.0387, 123.0440, 93.0576 | Vanillic acid | organic acid | [25] |
| 24 | 11.65 | [M + H]+ | 135.0544 | 153.0546 | −1.768 | C8H8O3 | 153.0544, 135.0803, 107.0848 | 4-Hydroxyphenylacetic acid | organic acid | [24] |
| 25 | 11.69 | [M + H]+ | 153.0544 | 153.0546 | −1.572 | C8H8O3 | 153.0544, 123.0440, 109.1014, 81.0704 | Vanillin | other compound | [24] |
| 26 | 11.88 | [M + H]+ | 181.0492 | 181.0495 | −1.907 | C9H8O4 | 181.0492, 163.1115, 139.0751, 111.0443 | Caffeic acid | organic acid | None 1 |
| 27 | 11.95 | [M + H]+ | 355.1015 | 355.1024 | −2.333 | C16H18O9 | 355.1015, 193.0492, 163.0386 | Cryptochlorogenic acid | polyphenol | [22] |
| 28 | 12.17 | [M + H]+ | 291.0857 | 291.0863 | −2.249 | C15H14O6 | 291.0857, 273.0747, 161.0595, 147.0438, 139.0387, 123.0441 | catechin | flavonoid | None 1 |
| 29 | 12.87 | [M + H]+ | 147.0438 | 147.0441 | −1.673 | C9H6O2 | 147.0438, 119.0492, 91.0546 | Coumarin | organic acid | [26] |
| 30 | 13.11 | [M + H]+ | 369.1173 | 369.1180 | −1.811 | C17H20O9 | 351.1061, 177.0540, 145.0282 | Methyl chlorogenate | polyphenol | [17] |
| 31 | 13.25 | [M + H]+ | 165.0544 | 165.0546 | −1.458 | C9H8O3 | 165.0544, 147.0438, 123.0441, 103.0545 | 2-Hydroxycinnamic acid | organic acid | [24] |
| 32 | 13.29 | [M + H]+ | 165.0544 | 165.0546 | −1.276 | C9H8O3 | 165.0544, 147.0438, 119.0492, 91.0546 | 4-Hydroxycinnamic acid | organic acid | [26] |
| 33 | 13.37 | [M + H]+ | 611.1595 | 611.1607 | −1.818 | C27H30O16 | 465.1052, 303.0492 | rutin | flavonoid | None 1 |
| 34 | 13.52 | [M + H]+ | 273.0751 | 273.0757 | −2.490 | C15H12O5 | 273.0751, 254.0443, 135.0801 | Butein | other compound | [27] |
| 35 | 13.64 | [M + H]+ | 465.1020 | 465.1028 | −1.704 | C21H20O12 | 303.0492, 153.0180, 137.0232, 85.0289 | Hyperoside | flavonoid | None 1 |
| 36 | 13.73 | [M + H]+ | 465.1019 | 465.1028 | −1.897 | C21H20O12 | 303.0491, 153.0179, 137.0231, 85.0289 | Isoquercitrin | flavonoid | [28] |
| 37 | 13.76 | [M + H]+ | 449.1070 | 449.1078 | −1.799 | C21H20O11 | 449.1070, 287.0542, 153.0179 | Luteoloside | flavonoid | None 1 |
| 38 | 13.81 | [M + H]+ | 195.0649 | 195.0652 | −1.565 | C10H10O4 | 177.0544, 163.0387, 149.0959, 145.0283, 117.0336 | Ferulic acid | organic acid | None 1 |
| 39 | 14.04 | [M + H]+ | 595.1646 | 595.1657 | −2.010 | C27H30O15 | 595.1657, 287.0543 | Nicotiflorin | flavonoid | [22] |
| 40 | 14.09 | [M + H]+ | 149.0595 | 149.0597 | −1.517 | C9H8O2 | 149.0595, 131.0491, 103.0545 | Cinnamic acid | organic acid | [25] |
| 41 | 14.12 | [M + H]+ | 517.1334 | 517.1341 | −1.223 | C25H24O12 | 517.1334, 499.1225, 163.0386. 145.0283, 135.0440, 117.0337, 89.0390 | Isochlorogenic acid B | polyphenol | None 1 |
| 42 | 14.41 | [M + H]+ | 449.1070 | 449.1078 | −1.888 | C21H20O11 | 449.1070, 287.0542 | Astragalin | flavonoid | [29] |
| 43 | 14.42 | [M + H]+ | 517.1333 | 517.1341 | −1.455 | C25H24O12 | 517.1333, 499.1224, 163.0387. 145.0283, 135.0440, 117.0336, 89.0390 | Isochlorogenic acid A | polyphenol | None 1 |
| 44 | 14.59 | [M + H]+ | 611.1957 | 611.1970 | −2.236 | C28H34O15 | 611.1957, 303.0492 | hesperidin | flavonoid | None 1 |
| 45 | 14.61 | [M + H]+ | 449.1068 | 449.1078 | −2.222 | C21H20O11 | 449.1078, 431.1379, 303.0829 | Quercitrin | flavonoid | [22] |
| 46 | 14.69 | [M + H]+ | 273.0750 | 273.0757 | −2.820 | C15H12O5 | 273.0750, 171.0286, 153.0179, 147.0437, 119.0492 | naringenin | flavonoid | None 1 |
| 47 | 14.77 | [M + H]+ | 517.1331 | 517.1341 | −1.919 | C25H24O12 | 517.1331, 499.1224, 163.0385. 145.0281, 135.0438, 117.0335, 89.0390 | Isochlorogenic acid C | polyphenol | None 1 |
| 48 | 14.96 | [M + H]+ | 189.1118 | 189.1121 | −1.774 | C9H16O4 | 189.1118, 171.1165, 161.1321, 147.0802 | Azelaic acid | organic acid | [26] |
| 49 | 15.46 | [M + H]+ | 211.0961 | 211.0965 | −1.968 | C11H14O4 | 211.0961, 179.0699, 167.0700, 151.0387 | Sinapyl alcohol | other compound | [24] |
| 50 | 16.79 | [M + H]+ | 287.0543 | 287.0550 | −2.524 | C15H10O6 | 287.0543, 269.0439, 153.0179, 135.0439 | Luteolin | flavonoid | None 1 |
| 51 | 16.89 | [M + H]+ | 303.0492 | 303.0499 | −2.538 | C15H10O7 | 303.0492, 285.0380, 257.0440, 229.0490 | Quercetin | flavonoid | [30] |
| 52 | 17.91 | [M + H]+ | 271.0593 | 271.0601 | −2.508 | C15H10O5 | 271.0594, 243.0643, 153.0180, 119.0492 | apigenin | flavonoid | None 1 |
| 53 | 18.09 | [M + H]+ | 287.0542 | 287.0550 | −2.942 | C15H10O6 | 287.0542, 258.0515, 165.0543, 153.0181 | Kaempferol | flavonoid | [30] |
| 54 | 18.22 | [M − H]− | 271.0615 | 271.0601 | 1.082 | C15H12O5 | 271.0615, 253.0506, 225.0557, 151.0024, 119.0484 | pinobanksin | flavonoid | [31] |
| 55 | 18.24 | [M + H]+ | 181.0856 | 181.0859 | −1.606 | C10H12O3 | 181.0859, 163.1115, 145.1009, 123.0804 | Coniferyl alcohol | other compound | [24] |
| 56 | 18.27 | [M + H]+ | 317.0649 | 317.1656 | −2.205 | C16H12O7 | 317.1016, 302.0779, 153.0543 | Isorhamnetin | flavonoid | None 1 |
| 57 | 20.08 | [M + H]+ | 375.1068 | 375.1074 | −1.690 | C19H18O8 | 375.1068, 360.1110 | Casticin | flavonoid | [26] |
| 58 | 20.42 | [M + H]+ | 318.2994 | 318.3003 | −2.798 | C18H39NO3 | 318.2994, 300.2887, 282.2783 | 2-Aminooctadecane-1,3,4-triol | other compound | [32] |
| 59 | 21.03 | [M + H]+ | 287.0905 | 287.0914 | −3.065 | C16H14O5 | 287.0905, 272.0672, 244.0681 | sakuranetin | flavonoid | [33] |
| 60 | 21.46 | [M + H]+ | 287.0905 | 287.0914 | −2.961 | C16H14O5 | 287.0905, 272.0669, 244.0703 | Isosakuranetin | flavonoid | [33] |
| 61 | 23.52 | [M + H]+ | 473.3613 | 473.3625 | −2.612 | C30H48O4 | 473.3613, 455.3511, 409.3450 | Maslinic acid | pentacyclic triterpenoids | [17] |
| 62 | 23.75 | [M + H]+ | 281.2467 | 281.2475 | −2.975 | C18H32O2 | 281.2467, 263.2369, 221.2256, 179.1425, 165.1273 | linoleic acid | other compound | [17] |
| 63 | 24.44 | [M + H]+ | 277.2154 | 277.2162 | −2.946 | C18H28O2 | 277.2155, 93.0702, 79.0547 | 12-Phenyldodecanoic acid | organic acid | [34] |
| 64 | 28.34 | [M + H]+ | 457.3666 | 457.3676 | −2.212 | C30H48O3 | 457.3666, 439.3564, 411.3608, 249.1844 | oleanolic acid | pentacyclic triterpenoids | [17] |
| 65 | 28.88 | [M + H]+ | 457.3664 | 457.3676 | −2.759 | C30H48O3 | 457.3664, 439.3557, 411.3609, 393.3507 | Ursolic Acid | pentacyclic triterpenoids | None 1 |
| Compound | ASPL | BC | CC | Degree |
|---|---|---|---|---|
| Quercetin | 1.69090909 | 0.20754625 | 0.59139785 | 98 |
| Apigenin | 2.39393939 | 0.03164871 | 0.41772152 | 39 |
| Ursolic Acid | 2.43030303 | 0.03084614 | 0.41147132 | 36 |
| Kaempferol | 2.45454545 | 0.02240119 | 0.40740741 | 35 |
| Luteolin | 2.45454545 | 0.02116046 | 0.40740741 | 34 |
| Epicatechin | 2.61212121 | 0.01209597 | 0.38283063 | 21 |
| Isorhamnetin | 2.64848485 | 0.00989390 | 0.37757437 | 18 |
| Rutin | 2.67272727 | 0.00675207 | 0.37414966 | 16 |
| Casticin | 2.78181818 | 0.00134943 | 0.35947712 | 7 |
| Quercitrin | 2.78181818 | 0.00109912 | 0.35947712 | 7 |
| Astragalin | 2.78181818 | 0.00155056 | 0.35947712 | 7 |
| Oleanolic acid | 2.78181818 | 0.00128503 | 0.35947712 | 7 |
| Hesperidin | 2.79393939 | 0.00069670 | 0.35791757 | 6 |
| Catechin | 2.79393939 | 0.00108638 | 0.35791757 | 6 |
| Procyanidin B2 | 2.80606061 | 0.00118610 | 0.35637149 | 5 |
| Hyperoside | 2.80606061 | 0.00031936 | 0.35637149 | 5 |
| Luteoloside | 2.81818182 | 0.00021088 | 0.35483871 | 4 |
| Butein | 2.83030303 | 0.00006069 | 0.35331906 | 3 |
| Procyanidin B1 | 2.83030303 | 0.00006069 | 0.35331906 | 3 |
| Cryptochlorogenic acid | 2.84242424 | 0.00001026 | 0.35181237 | 2 |
| Nicotiflorin | 2.84242424 | 0.00013911 | 0.35181237 | 2 |
| Chlorogenic acid | 2.84242424 | 0.00001026 | 0.35181237 | 2 |
| Name | IC50 (mg/mL) | Binding Energy (KJ/mol) |
|---|---|---|
| Acarbose | <0.0010 | −44.81 |
| Quercetin | 0.0090 | −30.88 |
| Kaempferol | 0.0646 | −29.12 |
| Isorhamnetin | 0.1588 | −29.33 |
| Luteolin | 0.1904 | −32.97 |
| Apigenin | 0.2508 | −32.17 |
| Rutin | 1.4790 | −13.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chi, B.; Liang, X.; Wang, L.; Bian, Y.; Zhang, M.; Tang, Z.; Wang, D.; Tian, Z. Component Characterization, In Vitro Activities and Molecular Mechanism of Cydonia oblonga Mill. against Diabetic. Pharmaceuticals 2022, 15, 1566. https://doi.org/10.3390/ph15121566
Chi B, Liang X, Wang L, Bian Y, Zhang M, Tang Z, Wang D, Tian Z. Component Characterization, In Vitro Activities and Molecular Mechanism of Cydonia oblonga Mill. against Diabetic. Pharmaceuticals. 2022; 15(12):1566. https://doi.org/10.3390/ph15121566
Chicago/Turabian StyleChi, Bingqing, Xilong Liang, Lihua Wang, Yifei Bian, Meng Zhang, Zhixin Tang, Danyang Wang, and Zhenhua Tian. 2022. "Component Characterization, In Vitro Activities and Molecular Mechanism of Cydonia oblonga Mill. against Diabetic" Pharmaceuticals 15, no. 12: 1566. https://doi.org/10.3390/ph15121566
APA StyleChi, B., Liang, X., Wang, L., Bian, Y., Zhang, M., Tang, Z., Wang, D., & Tian, Z. (2022). Component Characterization, In Vitro Activities and Molecular Mechanism of Cydonia oblonga Mill. against Diabetic. Pharmaceuticals, 15(12), 1566. https://doi.org/10.3390/ph15121566