
Serotonin 1A Receptor Pharmacotherapy and Neuroplasticity in Spinal Cord Injury

 and
and 
Abstract
:1. Introduction
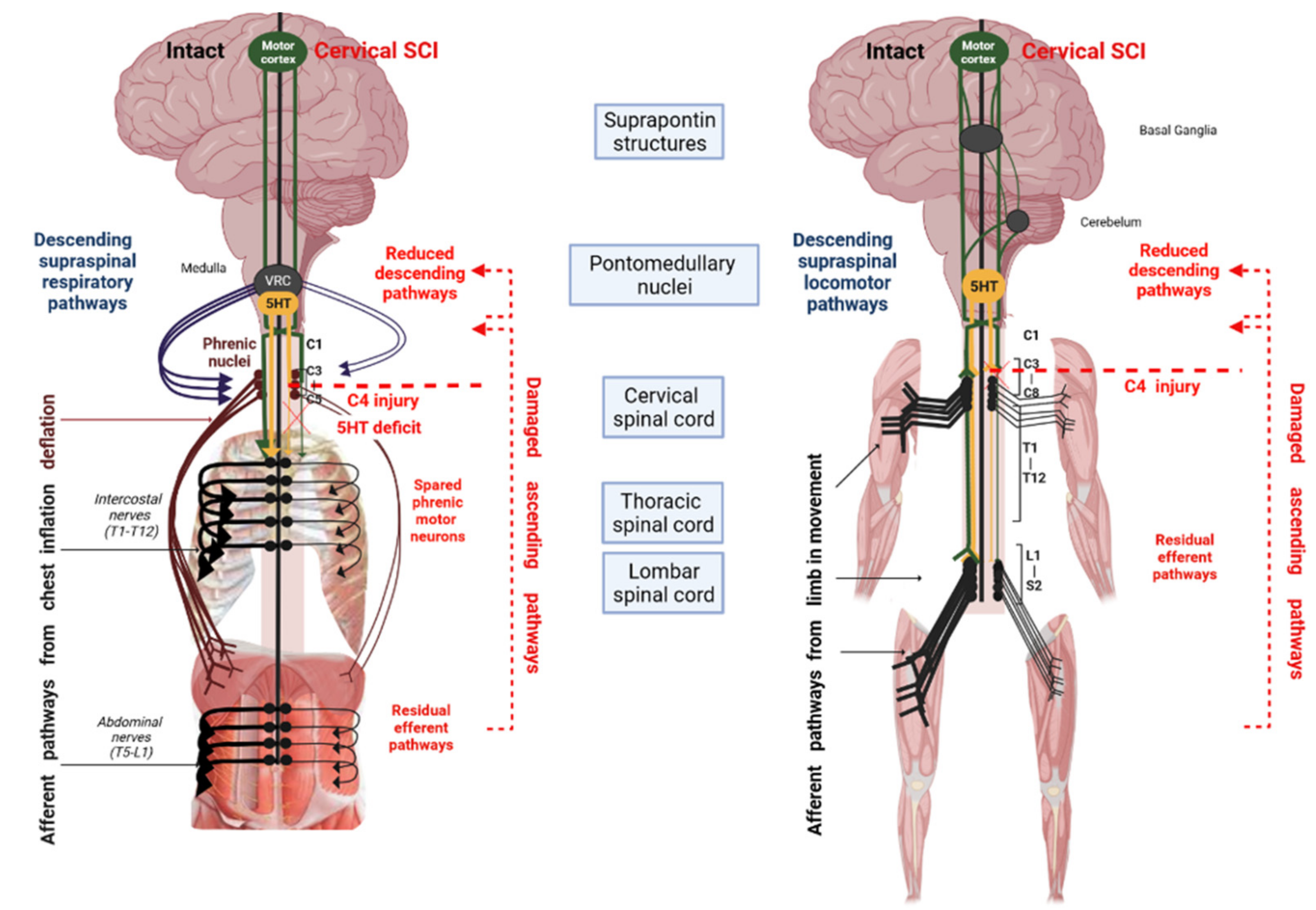
1.1. Locomotor Impact
1.2. Respiratory Impact
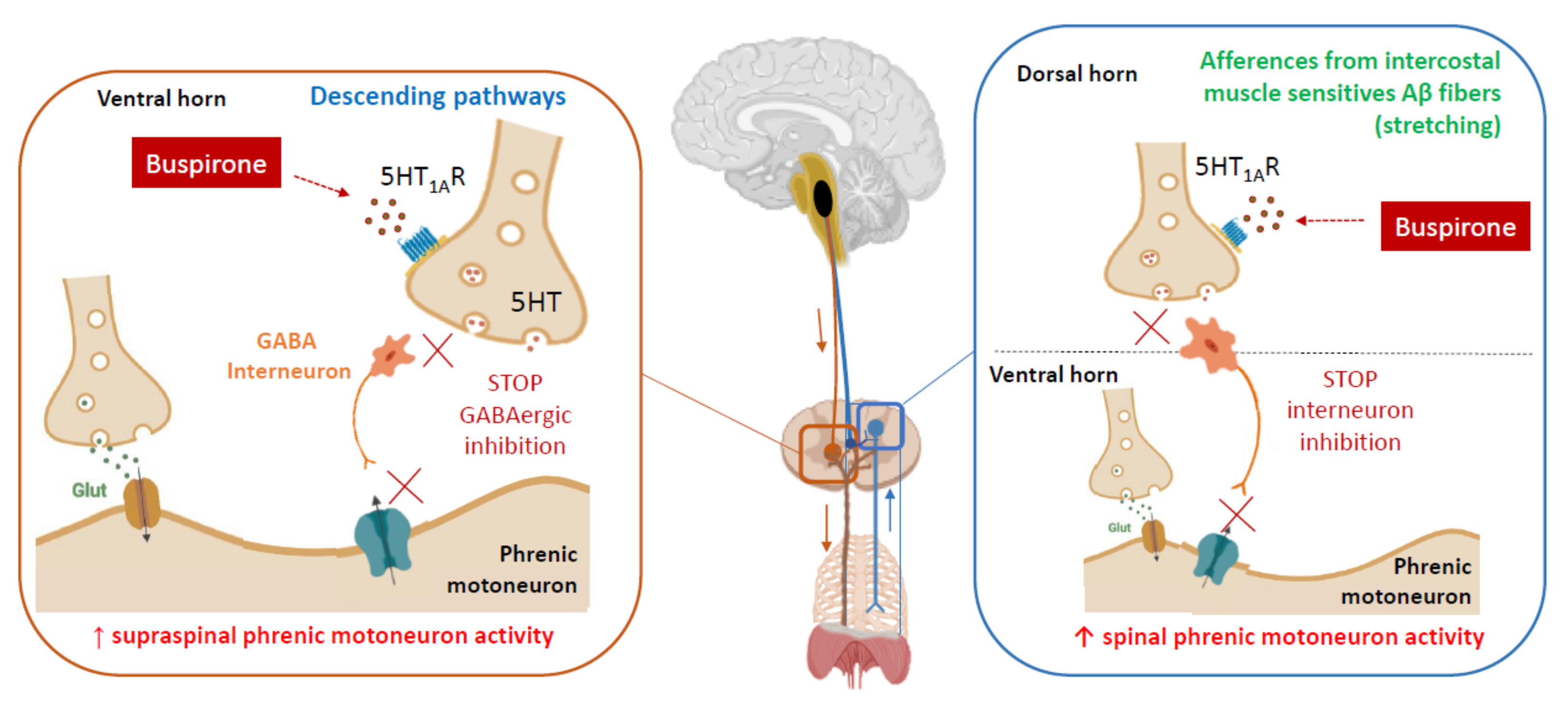
1.3. Serotonin 1A-R agonists and Neuromodulation
2. Locomotor Effects of 5-HT1A Agonists Pharmacology in SCI
2.1. Isolated Effects of 5-HT1A Agonist Pharmacology
2.2. Combinatorial Effects of 5-HT1A Agonist with Other Pharmacotherapies
2.3. Combinatorial Effects of 5-HT1A Agonist with Rehabilitation
2.4. Combinatorial Effects of 5-HT1A Agonist with Spinal Cord Stimulation
3. Respiratory Effects of 5-HT1A Agonist Pharmacology in SCI
4. Mechanistic Hypotheses
5. Clinical Impact
6. Perspectives and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wien, M.F.; Garshick, E.; Tun, C.G.; Lieberman, S.L.; Kelley, A.; Brown, R. Breathlessness and exercise in spinal cord injury. J. Spinal Cord Med. 1999, 22, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Bezdudnaya, T.; Marchenko, V.; Zholudeva, L.V.; Spruance, V.M.; Lane, M.A. Supraspinal respiratory plasticity following acute cervical spinal cord injury. Exp. Neurol. 2017, 293, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Buttry, J.L.; Goshgarian, H.G. Injection of WGA-Alexa 488 into the ipsilateral hemidiaphragm of acutely and chronically C2 hemisected rats reveals activity-dependent synaptic plasticity in the respiratory motor pathways. Exp. Neurol. 2014, 261, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K. Muscular, skeletal, and neural adaptations following spinal cord injury. J. Orthop. Sports Phys. Ther. 2002, 32, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzolato, C.; Gunduz, M.A.; Palipana, D.; Wu, J.; Grant, G.; Hall, S.; Dennison, R.; Zafonte, R.D.; Lloyd, D.G.; Teng, Y.D. Non-invasive approaches to functional recovery after spinal cord injury: Therapeutic targets and multimodal device interventions. Exp. Neurol. 2021, 339, 113612. [Google Scholar] [CrossRef]
- Vivodtzev, I.; Taylor, J.A. Cardiac, Autonomic, and Cardiometabolic Impact of Exercise Training in Spinal Cord Injury: A Qualitative Review. J. Cardiopulm. Rehabil. Prev. 2021, 41, 6–12. [Google Scholar] [CrossRef]
- Cragg, J.J.; Noonan, V.K.; Krassioukov, A.; Borisoff, J. Cardiovascular disease and spinal cord injury: Results from a national population health survey. Neurology 2013, 81, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Tanasescu, M.; Leitzmann, M.F.; Rimm, E.B.; Willett, W.C.; Stampfer, M.J.; Hu, F.B. Exercise type and intensity in relation to coronary heart disease in men. JAMA 2002, 288, 1994–2000. [Google Scholar] [CrossRef]
- Winslow, C.; Rozovsky, J. Effect of spinal cord injury on the respiratory system. Am. J. Phys. Med. Rehabil. 2003, 82, 803–814. [Google Scholar] [CrossRef]
- Savic, G.; DeVivo, M.J.; Frankel, H.L.; Jamous, M.A.; Soni, B.M.; Charlifue, S. Causes of death after traumatic spinal cord injury-a 70-year British study. Spinal Cord 2017, 55, 891–897. [Google Scholar] [CrossRef]
- Masmoudi, H.; Coirault, C.; Demoule, A.; Mayaux, J.; Beuvin, M.; Romero, N.; Assouad, J.; Similowski, T. Can phrenic stimulation protect the diaphragm from mechanical ventilation-induced damage? Eur. Respir. J. 2013, 42, 280–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.A.; Picard, G.; Porter, A.; Morse, L.R.; Pronovost, M.F.; Deley, G. Hybrid functional electrical stimulation exercise training alters the relationship between spinal cord injury level and aerobic capacity. Arch. Phys. Med. Rehabil. 2014, 95, 2172–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battikha, M.; Sa, L.; Porter, A.; Taylor, J.A. Relationship between pulmonary function and exercise capacity in individuals with spinal cord injury. Am. J. Phys. Med. Rehabil. 2014, 93, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Vivodtzev, I.; Napolitano, A.; Picard, G.; Taylor, J.A. Ventilatory support during whole-body row training improves oxygen uptake efficiency in patients with high-level spinal cord injury: A pilot study. Respir. Med. 2020, 171, 106104. [Google Scholar] [CrossRef]
- Sankari, A.; Vaughan, S.; Bascom, A.; Martin, J.L.; Badr, M.S. Sleep-Disordered Breathing and Spinal Cord Injury: A State-of-the-Art Review. Chest 2019, 155, 438–445. [Google Scholar] [CrossRef]
- Perrier, J.F.; Cotel, F. Serotonergic modulation of spinal motor control. Curr. Opin. Neurobiol. 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Chopek, J.W.; Sheppard, P.C.; Gardiner, K.; Gardiner, P.F. Serotonin receptor and KCC2 gene expression in lumbar flexor and extensor motoneurons posttransection with and without passive cycling. J. Neurophysiol. 2015, 113, 1369–1376. [Google Scholar] [CrossRef]
- Hadjiconstantinou, M.; Panula, P.; Lackovic, Z.; Neff, N.H. Spinal cord serotonin: A biochemical and immunohistochemical study following transection. Brain Res. 1984, 322, 245–254. [Google Scholar] [CrossRef]
- Otoshi, C.K.; Walwyn, W.M.; Tillakaratne, N.J.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Distribution and localization of 5-HT(1A) receptors in the rat lumbar spinal cord after transection and deafferentation. J. Neurotrauma 2009, 26, 575–584. [Google Scholar] [CrossRef]
- Nothias, J.M.; Mitsui, T.; Shumsky, J.S.; Fischer, I.; Antonacci, M.D.; Murray, M. Combined effects of neurotrophin secreting transplants, exercise, and serotonergic drug challenge improve function in spinal rats. Neurorehabil. Neural Repair 2005, 19, 296–312. [Google Scholar] [CrossRef]
- Garner, S.J.; Eldridge, F.L.; Wagner, P.G.; Dowell, R.T. Buspirone, an anxiolytic drug that stimulates respiration. Am. Rev. Respir. Dis. 1989, 139, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.B.; Goshgarian, H.G. Spinal cord injury in neonates alters respiratory motor output via supraspinal mechanisms. Exp. Neurol. 2007, 206, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.B.; Goshgarian, H.G. Spinal activation of serotonin 1A receptors enhances latent respiratory activity after spinal cord injury. J. Spinal Cord. Med. 2006, 29, 147–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saruhashi, Y.; Matsusue, Y.; Hukuda, S. Effects of serotonin 1A agonist on acute spinal cord injury. Spinal Cord 2002, 40, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Landry, E.S.; Lapointe, N.P.; Rouillard, C.; Levesque, D.; Hedlund, P.B.; Guertin, P.A. Contribution of spinal 5-HT1A and 5-HT7 receptors to locomotor-like movement induced by 8-OH-DPAT in spinal cord-transected mice. Eur. J. Neurosci. 2006, 24, 535–546. [Google Scholar] [CrossRef]
- Lapointe, N.P.; Guertin, P.A. Synergistic effects of D1/5 and 5-HT1A/7 receptor agonists on locomotor movement induction in complete spinal cord-transected mice. J. Neurophysiol. 2008, 100, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey-Gauthier, R.; Josset, N.; Bretzner, F.; Leblond, H. Facilitation of Locomotor Spinal Networks Activity by Buspirone after a Complete Spinal Cord Lesion in Mice. J. Neurotrauma 2018, 35, 2208–2221. [Google Scholar] [CrossRef]
- Develle, Y.; Leblond, H. Biphasic Effect of Buspirone on the H-Reflex in Acute Spinal Decerebrated Mice. Front. Cell Neurosci. 2019, 13, 573. [Google Scholar] [CrossRef]
- Jin, B.; Alam, M.; Tierno, A.; Zhong, H.; Roy, R.R.; Gerasimenko, Y.; Lu, D.C.; Edgerton, V.R. Serotonergic Facilitation of Forelimb Functional Recovery in Rats with Cervical Spinal Cord Injury. Neurotherapeutics 2021, 18, 1226–1243. [Google Scholar] [CrossRef]
- Sławińska, U.; Majczyński, H.; Dai, Y.; Jordan, L.M. The upright posture improves plantar stepping and alters responses to serotonergic drugs in spinal rats. J. Physiol. 2012, 590, 1721–1736. [Google Scholar] [CrossRef]
- Guertin, P.A.; Ung, R.V.; Rouleau, P.; Steuer, I. Effects on locomotion, muscle, bone, and blood induced by a combination therapy eliciting weight-bearing stepping in nonassisted spinal cord-transected mice. Neurorehabil. Neural Repair 2011, 25, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guertin, P.A.; Ung, R.V.; Rouleau, P. Oral administration of a tri-therapy for central pattern generator activation in paraplegic mice: Proof-of-concept of efficacy. Biotechnol. J. 2010, 5, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishna, M.; Steuer, I.; Prince, F.; Roberts, M.; Mongeon, D.; Kia, M.; Dyck, S.; Matte, G.; Vaillancourt, M.; Guertin, P.A. Double-Blind, Placebo-Controlled, Randomized Phase I/IIa Study (Safety and Efficacy) with Buspirone/Levodopa/Carbidopa (SpinalonTM) in Subjects with Complete AIS A or Motor-Complete AIS B Spinal Cord Injury. Curr. Pharm. Des. 2017, 23, 1789–1804. [Google Scholar] [CrossRef]
- Ung, R.V.; Rouleau, P.; Guertin, P.A. Functional and physiological effects of treadmill training induced by buspirone, carbidopa, and L-DOPA in clenbuterol-treated paraplegic mice. Neurorehabil. Neural Repair 2012, 26, 385–394. [Google Scholar] [CrossRef]
- Ganzer, P.D.; Beringer, C.R.; Shumsky, J.S.; Nwaobasi, C.; Moxon, K.A. Serotonin receptor and dendritic plasticity in the spinal cord mediated by chronic serotonergic pharmacotherapy combined with exercise following complete SCI in the adult rat. Exp. Neurol. 2018, 304, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Vivodtzev, I.; Picard, G.; O’Connor, K.; Taylor, J.A. Serotonin 1A agonist and cardiopulmonary improvements with whole-body exercise in acute, high-level spinal cord injury: A retrospective analysis. Eur. J. Appl. Physiol. 2021, 121, 453–463. [Google Scholar] [CrossRef]
- Morgan, J.W.; Solinsky, R. Buspirone for functional improvement after acute traumatic spinal cord injury: A propensity score-matched cohort study. Spinal Cord 2021, 59, 563–570. [Google Scholar] [CrossRef]
- Moshonkina, T.R.; Shapkova, E.Y.; Sukhotina, I.A.; Emeljannikov, D.V.; Gerasimenko, Y.P. Effect of Combination of Non-Invasive Spinal Cord Electrical Stimulation and Serotonin Receptor Activation in Patients with Chronic Spinal Cord Lesion. Bull. Exp. Biol. Med. 2016, 161, 749–754. [Google Scholar] [CrossRef]
- Gad, P.; Gerasimenko, Y.; Zdunowski, S.; Turner, A.; Sayenko, D.; Lu, D.C.; Edgerton, V.R. Weight Bearing Over-ground Stepping in an Exoskeleton with Non-invasive Spinal Cord Neuromodulation after Motor Complete Paraplegia. Front. Neurosci. 2017, 11, 333. [Google Scholar] [CrossRef]
- Freyvert, Y.; Yong, N.A.; Morikawa, E.; Zdunowski, S.; Sarino, M.E.; Gerasimenko, Y.; Edgerton, V.R.; Lu, D.C. Engaging cervical spinal circuitry with non-invasive spinal stimulation and buspirone to restore hand function in chronic motor complete patients. Sci. Rep. 2018, 8, 15546. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Roy, R.R.; Edgerton, V.R. Epidural stimulation: Comparison of the spinal circuits that generate and control locomotion in rats, cats and humans. Exp. Neurol. 2008, 209, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.D.; Bingaman, M.; Taveira-DaSilva, A.M.; Pace, P.P.; Gillis, R.A.; Wrathall, J.R. Serotonin 1A receptor agonists reverse respiratory abnormalities in spinal cord-injured rats. J. Neurosci. 2003, 23, 4182–4189. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Liao, W.L.; Newton, K.M.; Onario, R.C.; King, A.M.; Desilets, F.C.; Woodard, E.J.; Eichler, M.E.; Frontera, W.R.; Sabharwal, S.; et al. Respiratory abnormalities resulting from midcervical spinal cord injury and their reversal by serotonin 1A agonists in conscious rats. J. Neurosci. 2005, 25, 4550–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheriyan, T.; Ryan, D.J.; Weinreb, J.H.; Cheriyan, J.; Paul, J.C.; Lafage, V.; Kirsch, T.; Errico, T.J. Spinal cord injury models: A review. Spinal Cord 2014, 52, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Maresh, S.; Prowting, J.; Vaughan, S.; Kruppe, E.; Alsabri, B.; Yarandi, H.; Badr, M.S.; Sankari, A. Buspirone decreases susceptibility to hypocapnic central sleep apnea in chronic SCI patients. J. Appl. Physiol. 2020, 129, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Perrier, J.F.; Rasmussen, H.B.; Christensen, R.K.; Petersen, A.V. Modulation of the intrinsic properties of motoneurons by serotonin. Curr. Pharm. Des. 2013, 19, 4371–4384. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Fornal, C.A. 5-HT and motor control: A hypothesis. Trends Neurosci. 1993, 16, 346–352. [Google Scholar] [CrossRef]
- Aggelopoulos, N.C.; Burton, M.J.; Clarke, R.W.; Edgley, S.A. Characterization of a descending system that enables crossed group II inhibitory reflex pathways in the cat spinal cord. J. Neurosci. 1996, 16, 723–729. [Google Scholar] [CrossRef]
- Thor, K.B.; Nickolaus, S.; Helke, C.J. Autoradiographic localization of 5-hydroxytryptamine1A, 5-hydroxytryptamine1B and 5-hydroxytryptamine1C/2 binding sites in the rat spinal cord. Neuroscience 1993, 55, 235–252. [Google Scholar] [CrossRef]
- Gautier, H. Respiratory responses of the anesthetized rabbit to vagotomy and thoracic dorsal rhizotomy. Respir. Physiol. 1973, 17, 238–247. [Google Scholar] [CrossRef]
- Jensen, V.N.; Alilain, W.J.; Crone, S.A. Role of Propriospinal Neurons in Control of Respiratory Muscles and Recovery of Breathing Following Injury. Front. Syst. Neurosci. 2019, 13, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, J.; Bezdudnaya, T.; Zholudeva, L.V.; Detloff, M.R.; Reier, P.J.; Lane, M.A.; Fuller, D.D. Histological identification of phrenic afferent projections to the spinal cord. Respir. Physiol. Neurobiol. 2017, 236, 57–68. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Brown, I.G.; Pender, M.P. Apneusis responding to buspirone in multiple sclerosis. Mult. Scler. 2008, 14, 705–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andaku, D.K.; Mercadante, M.T.; Schwartzman, J.S. Buspirone in Rett syndrome respiratory dysfunction. Brain Dev. 2005, 27, 437–438. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Y.; Wienecke, J.; Chen, M.; Hultborn, H.; Zhang, M. The time course of serotonin 2A receptor expression after spinal transection of rats: An immunohistochemical study. Neuroscience 2011, 177, 114–126. [Google Scholar] [CrossRef]
- Barrett, K.T.; Kinney, H.C.; Li, A.; Daubenspeck, J.A.; Leiter, J.C.; Nattie, E.E. Subtle alterations in breathing and heart rate control in the 5-HT1A receptor knockout mouse in early postnatal development. J. Appl. Physiol. 2012, 113, 1585–1593. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Authors | Species/Type of SCI | Drug and Dose | Combined with | Method | Main Outcome Measures | Main Results |
|---|---|---|---|---|---|---|
| Drug Alone | ||||||
| Saruhashi et al., 2002 | Rats, in vitro compressive injury, n = 24 | Tandospirone, 10 and 100 µM, single dose, Mode: incubated | Monotherapy | Electrical stimulation recording inducing action potential. | Amplitude change and latency change of the evoked action potential | ↑ recovery of mean action potential |
| Landry et al., 2006 | Mice, Th9/Th10 transection, 7 day post-Tx, n = 36 | 8-OH-DPAT, 1mg/kg, single dose, Mode: ip | Monotherapy | Quantitative kinematic analysis of hindlimb movements. In situ hybridization | Locomotor-like movements (LM) and non-locomotor-like (NLM) movements. Footstep amplitude. Angular excursion for the ankle. | Induction of locomotor like movement. |
| Lapointe et al., 2008 | Mice, Th9/Th10 transection, 6 days post-Tx, n = 40 | 8-OH-DPAT, 0.5 mg/kg, single dose, Mode: ip 1 | Monotherapy | Assessment of hindlimb movements | LM, NLM 2 | Acute LM induction |
| Jeffrey-Gauthier et al., 2018 | Mice, Th7-Th8 hemisection, 2 days post-Tx, n = 19 | Buspirone, 8 mg/kg/day, single dose, Mode: ip | Monotherapy | Assessment of locomotor recovery. Histology | Number of steps. Number of consecutive steps. | ↑ Number of steps taken, Improved paw positioning, ↑ Locomotor spinal networks activity |
| Develle et al., 2020 | Mice, decerebrated, n = 20 | Buspirone, 8mg/kg, single dose, Mode: ip | Monotherapy | H-reflex recording | Frequency-dependent depression of the H-reflex | ↑ Reflex excitability |
| Ahmed et al., 2021 | Rats, C4 bilateral crush injury, 1 week post-injury, n = 17 | Buspirone, 1 dose/day, 1.5–2.5–3.5 mg/kg, single dose, Mode: ip | Monotherapy | EMG, grip strength, reaching and grasping tasks | Reaching (R) and 3 grasping (G) functions | Low doses facilitate R and G and improve forelimb grip strength |
| Jin et al., 2021 | Rats, C4 bilateral crush injury, 1 to 8 weeks post-injury, n = 29 | Buspirone, 1–2 mg/kg, Single dose Mode: ip | Monotherapy | R and G success rates | Forelimb motor function recovery, performance within 2 weeks of buspirone withdrawal | |
| Combined with other pharmacology | ||||||
| Guertin et al., 2010 | Mice, Th9/Th10 transection, 7 days post-Tx n = 22 | Buspirone, differential 0.5, 1.5, 7.5, or 10 mg/kg, Mode: oral | Tritherapy 4 | Behavioral assays | Locomotor-like behavior Movement frequency | Locomotor activity induction |
| Guertin et al., 2012 | Mice, Th9/TH10 transection, 1week post-Tx, n = 21 | Buspirone, 1.5 mg/kg, repetitive administration, Mode: subcutaneous | Tritherapy | Behavioral assays, Histology | Hindlimb movement, induced stepping | Central pattern generator activation, induced episodes of weight-bearing stepping |
| Slawinska et al., 2012 | Rats, Th9/Th10 transection, 10 weeks post-Tx, n = 15 | 8-OH-DPAT, 0.2–0.4 mg/kg, single dose Mode: ip | Quipazine | EMG, Evaluation of hindlimb, locomotor performance | Hindlimb locomotor movements, Weight supported stepping, EMG 5 | LM activity improvement mitigated the need to activate the LM with exteroceptive stimulation |
| Radhakrishna et al., 2017 | Patients with complete and incomplete SCI, > 3 months post-SCI, n = 45 | BuspironeGrp1: 10mg, Grp2: 15 mg, Grp3: 25 mg, Grp4: 35 mg, Grp5: 50 mg, Grp6: 75 mg, repetitive administration, Mode: oral | Tritherapy 6 | Dose-escalation study of Buspirone | EMG, leg movement | ↑ EMG activity ↑ locomotor-like characteristics |
| Combined with exercise training | ||||||
| Ung et al., 2012 | Mice, Th9/Th10 transection, 1-week post-Tx, n = 43 | Buspirone (3 mg/kg)3 times/w × 8 weeks, repetitive administration, Mode: ip | Tritherapy4 + training + clenbuterol | LM 7 assessment, Muscle fiber immunofluorescence | Locomotor movement, Movement frequency, Cross-sectional area of muscle fibers | ↑ locomotor movement and muscle properties, ↑ type II fiber cross sectional area values, ↓ fiber type conversion |
| Ganzer et al., 2018 | Rats, Th8/Th9 transection, 1 week and 8 weeks post-Tx, n = 41 | 8-OH-DPAT, single dose: 0.075–0.125 mg/kg, repetitive administration: 0.125 mg/kg, 5 days/week × 2–8 weeks, Mode: ip | Bike therapy | Acute dose response test, Behavioral testing, Locomotor Scoring Immunohistochemisty | Open field scores, Spinal 5-HT1AR densities caudal to the SCI, MAP2 dendrite density | Significant open-field weigh-supported stepping, mediated in part by restoring spinal dendritic density |
| Jeffrey-Gauthier et al., 2018 | Mice, Th7/Th8 hemisection, 2 days post-Tx, n = 19 | Buspirone, 8 mg/kg/day, single dose, Mode: ip | Treadmill | Assessment oflocomotor recovery, Histology | Number of steps, Step occurrence, Number of consecutive steps | ↑ Number of steps taken Improved paw positioning, ↑ Locomotor spinal networks activity |
| Morgan et al., 2021 | Patients with acute traumatic SCI, cohort study from 2011 to 2017, n = 84 | Inpatient rehabilitation +/− treated by Buspirone | Inpatient rehabilitation +/− treated by Buspirone | Functional scores comparison | Upper extremity motor score, lower extremity motor score, American Spinal Injury Association Impairment Scale, neurological level of injury, and functional impairment measure | ↑ 1-year conversion rate to incomplete injury (42.9% with Buspirone vs 21.2% without Buspirone, though this was not significantly different from non-buspirone local controls |
| Vivodtzev et al., 2021 | Patients with complete and incomplete SCI at Th3 or above, <2 years post injury, n = 21 | Buspirone, 29 ± 17 mg/day × 3 months, repetitive administration, Mode: oral | FES 8-rowing | FES-rowing testCardiopulmonaryfunction testing Spirometry | Peak Power output, VO2, VCO2, Vt, Bf 9 | ↑ Aerobic capacity ↑ Ventilatory capacity |
| Combined with Spinal stimulation | ||||||
| Monshonkina et al., 2017 | Patients with complete thoracic SCI, > 1-year post-SCI, n = 10 | Buspirone, Differential-dose 7.5 mg × 2/day × 18 days, repetitive administration, Mode: oral | Spinal stimulation | Percutaneous electrical stimulation of the spinal cord | Activity of the knee and Achille assessment, Rehabilitation status assessment | Muscular force improvement, potentiated the effect of spinal cord stimulation, ↑ pain sensitivity |
| Gad et al., 2017 | Patient with complete SCI at Th9 and L1, 4 years post-SCI, n = 1 | Buspirone repetitive administration/1 week Mode: oral | Spinal stimulation | EKSO bionics exoskeleton, Painless cutaneous (pcEmc) and Pharmacological (fEmc) enabling motor control EMG | Evoked potential, Robotic assistance, Percent effort, EMG amplitude | ↓ robotic assistance, ↑ EMG activity Change in knee angle |
| Freyvert, Sci Rep. 2018 | Patient with complete SCI at C5 or above, >1-year post-SCI, n = 6 | Buspirone, 15 mg/day × 15 days, repetitive administration, Mode: oral | Spinal stimulation | Handgrip force measurement, EMG | Functional metrics, EMG amplitude, Changes in mean grip strength | ↑ Hand function↑ EMG amplitudes |
| Authors | Species/Type of SCI | Drug & Dose | Combined with | Method | Main Outcome Measures | Main Results |
| Teng et al., 2003 | Rats T8 hemicompression 24 h and 7 days post-injury n = 16 | Buspirone, 1.5 mg/kg, Single dose Mode: ip | Monotherapy | Plethysmography at ambient air and 7% CO2 exposure | Bf, VT 1, Ventilatory response to 7% CO2 | Normalized Bf, Vt and respiratory response to 7% CO2 |
| Choi et al., 2005 | Rats C5 Hemicontusion 2, 4, and 6 weeks post-injuryn = 44 | 8-OH-DPAT 2 250 µg/kg, or Buspirone 1.5 mg/kg, Single dose Mode: ip | Monotherapy | Plethysmography at ambient air and 7% CO2 exposure Histology | Bf, VT, Ve while breathing ambient air or 7% CO2 | ↑ hypercapnic ventilatory response to CO2, 2 and 4 weeks post injury for up to 4 h |
| Zimmer et al., 2006 | Rats C2 Hemi-transected 24 h/48 h and 1 week post-injury n = 52 | 8-OH-DPAT, 17 µg/kg, Mode: intravenous | Monotherapy | Phrenic activity recording | PO2, PCO2, Apneic thresholds, Phrenic nerve activity | ↑ phrenic activity and amplitude, ↑ phrenic amplitude, ↑ respiratory response and f after systemic injection |
| Maresh et al., 2020 | Chronic SCI patients C5-T3 n = 8 | Buspirone, 30 mg/day × 13 days, repetitive administration, Mode: oral | Monotherapy | Pneumotachometer connected to a tight-fitting nasal mask, ECG | CO2 reserve, Apneic threshold Vt, PETCO2, PETO2 3 | Widened CO2 reserve ↓ hypocapnic central sleep apnea |
| Vivodtzev et al., 2021 | Subacute SCI patients < 2 years post injury C5-T3 n = 21 | Buspirone, 29 ± 17 mg/day × 3 months, repetitive administration, Mode: oral | FES-rowing 4 | FES-rowing test, Cardiopulmonary function testing, Spirometry | Peak VO2, VCO2, Vt, Bf Respiratoy function (FEV1 and FVC) 5 | ↑ Aerobic capacity ↑ Ventilatory capacity Changes in ventilatory capacity proportional to changes in respiratory function |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajjig, A.; Cayetanot, F.; Taylor, J.A.; Bodineau, L.; Vivodtzev, I. Serotonin 1A Receptor Pharmacotherapy and Neuroplasticity in Spinal Cord Injury. Pharmaceuticals 2022, 15, 460. https://doi.org/10.3390/ph15040460
Bajjig A, Cayetanot F, Taylor JA, Bodineau L, Vivodtzev I. Serotonin 1A Receptor Pharmacotherapy and Neuroplasticity in Spinal Cord Injury. Pharmaceuticals. 2022; 15(4):460. https://doi.org/10.3390/ph15040460
Chicago/Turabian StyleBajjig, Afaf, Florence Cayetanot, J. Andrew Taylor, Laurence Bodineau, and Isabelle Vivodtzev. 2022. "Serotonin 1A Receptor Pharmacotherapy and Neuroplasticity in Spinal Cord Injury" Pharmaceuticals 15, no. 4: 460. https://doi.org/10.3390/ph15040460
APA StyleBajjig, A., Cayetanot, F., Taylor, J. A., Bodineau, L., & Vivodtzev, I. (2022). Serotonin 1A Receptor Pharmacotherapy and Neuroplasticity in Spinal Cord Injury. Pharmaceuticals, 15(4), 460. https://doi.org/10.3390/ph15040460