
Effect of Escitalopram on the Number of DCX-Positive Cells and NMUR2 Receptor Expression in the Rat Hippocampus under the Condition of NPSR Receptor Blockade

 , ,
, ,  and
and 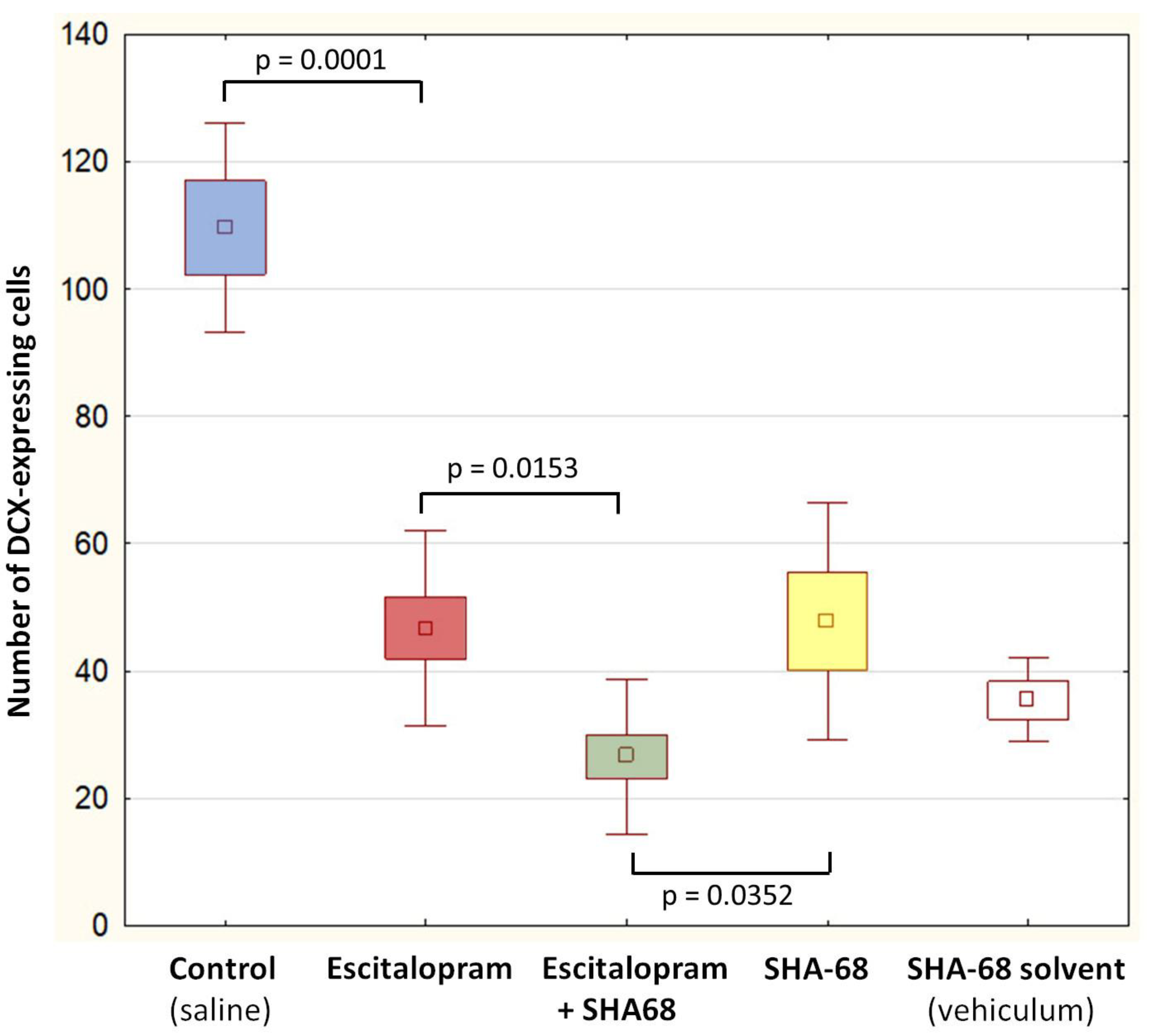{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
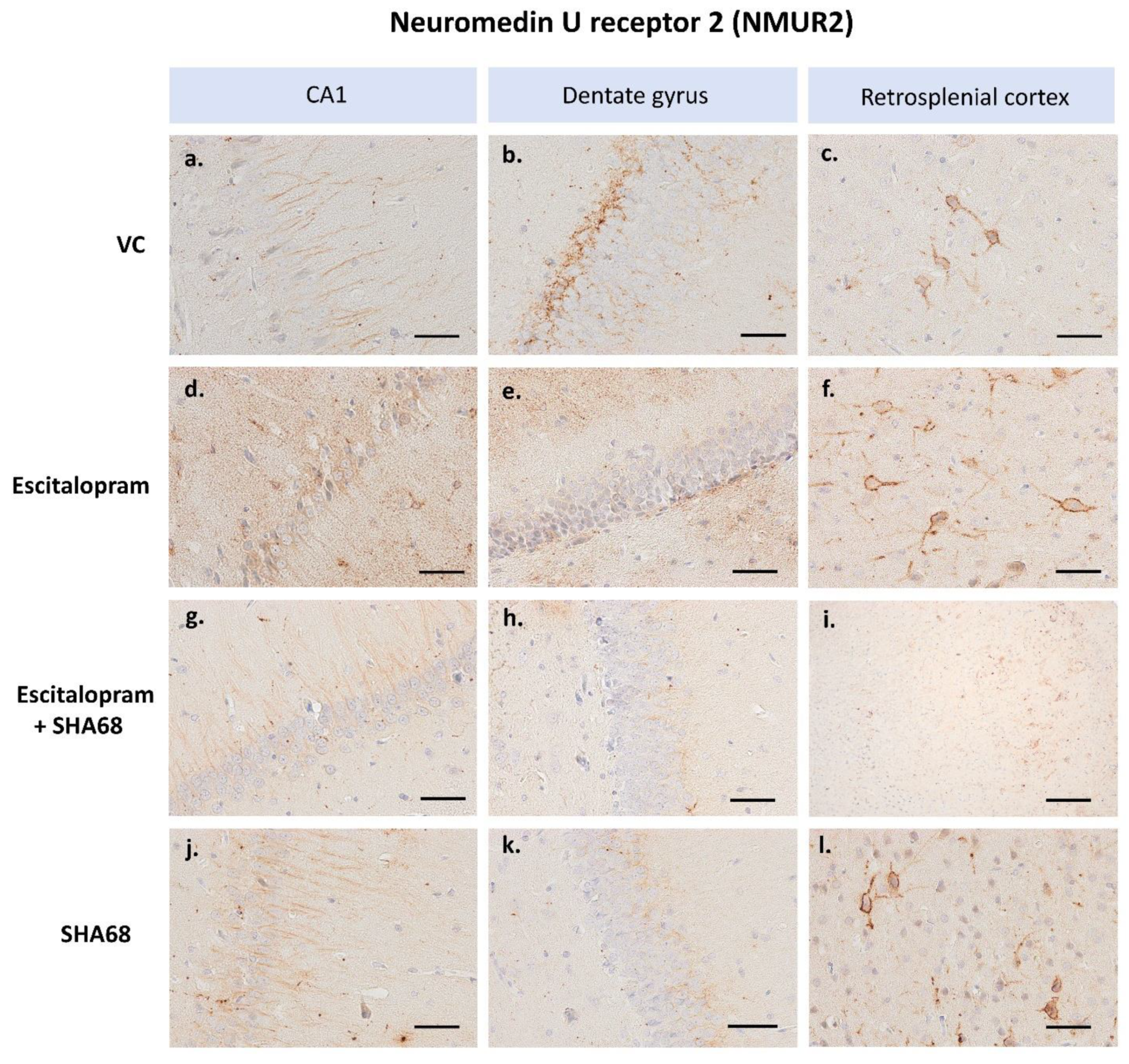
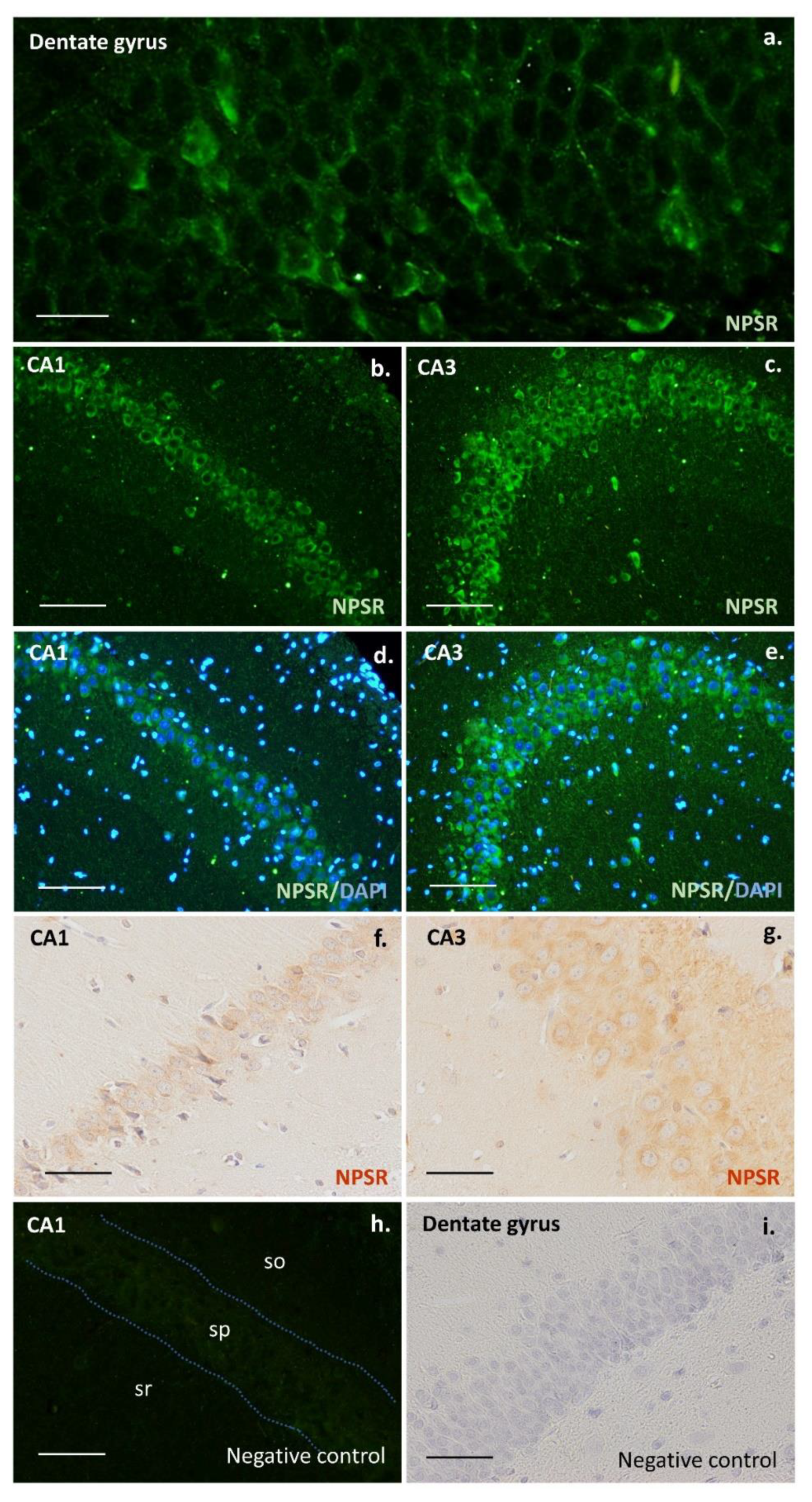
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Administration
4.3. Brain Tissue Collection
4.4. Immunohistochemistry and Immunofluorescence
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rana, T.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Abdeen, A.; Ibrahim, S.F.; Mani, V.; Iqbal, M.S.; Bhatia, S.; et al. Exploring the role of neuropeptides in depression and anxiety. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 114, 110478. [Google Scholar] [CrossRef] [PubMed]
- Pałasz, A.; Janas-Kozik, M.; Borrow, A.; Arias-Carrión, O.; Worthington, J.J. The potential role of the novel hypothalamic neuropeptides nesfatin-1, phoenixin, spexin and kisspeptin in the pathogenesis of anxiety and anorexia nervosa. Neurochem. Int. 2018, 113, 120–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormos, V.; Gaszner, B. Role of neuropeptides in anxiety, stress, and depression: From animals to humans. Neuropeptides 2013, 47, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jiménez, E.P.; Terreros-Roncal, J.; Flor-García, M.; Rábano, A.; Llorens-Martín, M.J. Evidences for Adult Hippocampal Neurogenesis in Humans. Neuroscience 2021, 41, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.; Spector, R. A balanced evaluation of the evidence for adult neurogenesis in humans: Implication for neuropsychiatric disorders. Brain Struct. Funct. 2019, 224, 2281–2295. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Zaben, M.J.; Gray, W.P. Neuropeptides and hippocampal neurogenesis. Neuropeptides 2013, 47, 431–438. [Google Scholar] [CrossRef]
- David, D.J.; Samuels, B.A.; Rainer, Q.; Wang, J.W.; Marsteller, D.; Mendez, I.; Drew, M.; Craig, D.A.; Guiard, B.P.; Guilloux, J.P.; et al. Neurogenesis-dependent and -independent effects of fluoxetine in an animal model of anxiety/depression. Neuron 2009, 62, 479–493. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Brazo, J.; Baekelandt, V.; Encinas, J.M. Neurogenesis as a new target for the development of antidepressant drugs. Curr. Pharm. Des. 2014, 20, 3763–3775. [Google Scholar] [CrossRef]
- Belovicova, K.; Bogi, E.; Koprdova, R.; Ujhazy, E.; Mach, M.; Dubovicky, M. Effects of venlafaxine and chronic unpredictable stress on behavior and hippocampal neurogenesis of rat dams. Neuroendocrinol. Lett. 2017, 38, 19–26. [Google Scholar]
- Chen, A.T.; Nasrallah, H.A. Neuroprotective effects of the second generation antipsychotics. Schizophr. Res. 2019, 208, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Krzystanek, M.; Krzystanek, E.; Skałacka, K.; Pałasz, A. Enhancement in Phospholipase D Activity as a New Proposed Molecular Mechanism of Haloperidol-Induced Neurotoxicity. Int. J. Mol. Sci. 2020, 21, 9265. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, S.; Pai, N.B.; Deng, C. The Association of Antidepressant Medication and Body Weight Gain. Online J. Health Allied Sci. 2013, 12, 1–9. [Google Scholar]
- Owens, M.J.; Knight, D.L.; Nemeroff, C.B. Second-generation SSRIs: Human monoamine transporter binding profile of escitalopram and R-fluoxetine. Biol. Psychiatry 2001, 50, 345–350. [Google Scholar] [CrossRef]
- Reinscheid, R.K.; Ruzza, C. Pharmacology, Physiology and Genetics of the Neuropeptide S System. Pharmaceuticals 2021, 14, 401. [Google Scholar] [CrossRef]
- Pape, H.C.; Jüngling, K.; Seidenbecher, T.; Lesting, J.; Reinscheid, R.K. Neuropeptide S: A transmitter system in the brain regulating fear and anxiety. Neuropharmacology 2010, 58, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Reinscheid, R.K.; Xu, Y.L. Neuropeptide S and its receptor: A newly deorphanized G protein-coupled receptor system. Neuroscientist 2005, 11, 532–538. [Google Scholar] [CrossRef]
- Clark, S.D.; Duangdao, D.M.; Schulz, S.; Zhang, L.; Liu, X.; Xu, Y.L.; Reinscheid, R.K. Anatomical characterization of the neuropeptide S system in the mouse brain by in situ hybridization and immunohistochemistry. J. Comp. Neurol. 2011, 519, 1867–1893. [Google Scholar] [CrossRef]
- Xu, Y.L.; Reinscheid, R.K.; Huitron-Resendiz, S.; Clark, S.D.; Wang, Z.; Lin, S.J.; Brucher, F.A.; Zeng, J.; Ly, N.K.; Henriksen, S.J.; et al. Neuropeptide S: A neuropeptide promoting arousal and anxiolytic-like effects. Neuron 2004, 43, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.L.; Gall, C.M.; Jackson, V.R.; Civelli, O.; Reinscheid, R.K. Distribution of neuropeptide S receptor mRNA and neurochemical characteristics of neuropeptide S-expressing neurons in the rat brain. J. Comp. Neurol. 2007, 500, 84–102. [Google Scholar] [CrossRef]
- Tobinski, A.M.; Rappeneau, V. Role of the Neuropeptide S System in Emotionality, Stress Responsiveness and Addiction-Like Behaviours in Rodents: Relevance to Stress-Related Disorders. Pharmaceuticals 2021, 14, 780. [Google Scholar] [CrossRef] [PubMed]
- Cannella, N.; Economidou, D.; Kallupi, M.; Stopponi, S.; Heiling, M.; Massi, M.; Ciccocioppo, R. Persistent Increase of Alcohol-Seeking Evoked by Neuropeptide S: An Effect Mediated by the Hypothalamic Hypocretin System. Neuropsychopharmacology 2009, 34, 2125–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoetxea, X.; Goedecke, L.; Remmes, J.; Blaesse, P.; Grosch, T.; Lesting, J.; Pape, H.P.; Jüngling, K. Human-Specific Neuropeptide S Receptor Variants Regulate Fear Extinction in the Basal Amygdala of Male and Female Mice Depending on Threat Salience. Biol. Psychiatry 2021, 90, 145–155. [Google Scholar] [CrossRef]
- Grund, T.; Neumann, I.D. Neuropeptide S Induces Acute Anxiolysis by Phospholipase C-Dependent Signaling within the Medial Amygdala. Neuropsychopharmacology 2018, 43, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Domschke, K.; Reif, A.; Weber, H.; Richter, J.; Hohoff, C.; Ohrmann, P.; Pedersen, A.; Bauer, J.; Suslow, T.; Kugel, H.; et al. Neuropeptide S receptor gene - converging evidence for a role in panic disorder. Mol. Psychiatry 2011, 16, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Ebner, K.; Rjabokon, A.; Pape, H.C.; Singewald, N. Increased in vivo release of neuropeptide S in the amygdala of freely moving rats after local depolarisation and emotional stress. Amino Acids 2011, 41, 991–996. [Google Scholar] [CrossRef] [Green Version]
- Adori, C.; Barde, S.; Bogdanovic, N.; Uhlén, M.; Reinscheid, R.R.; Kovacs, G.G.; Hökfelt, T. Neuropeptide S- and Neuropeptide S receptor-expressing neuron populations in the human pons. Front. Neuroanat. 2015, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Teranishi, H.; Hanada, R. Neuromedin U, a Key Molecule in Metabolic Disorders. Int. J. Mol. Sci. 2021, 22, 4238. [Google Scholar] [CrossRef]
- Malendowicz, L.K.; Rucinski, M. Neuromedins NMU and NMS: An Updated Overview of Their Functions. Front. Endocrinol. 2021, 12, 713961. [Google Scholar] [CrossRef]
- Brighton, P.J.; Wise, A.; Dass, N.B.; Willars, G.B. Paradoxical behavior of neuromedin U in isolated smooth muscle cells and intact tissue. J. Pharmacol. Exp. Ther. 2008, 325, 154–164. [Google Scholar] [CrossRef]
- Szekeres, P.G.; Muir, A.I.; Spinage, L.D.; Miller, J.E.; Butler, S.I.; Smith, A.; Rennie, G.I.; Murdock, P.R.; Fitzgerald, L.R.; Wu, H.L.; et al. Neuromedin U is a potent agonist at the orphan G protein-coupled receptor FM3. J. Biol. Chem. 2000, 275, 20247–20250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, A.D.; Wang, R.; Pong, S.S.; Mellin, T.N.; Strack, A.; Guan, X.M.; Zeng, Z.; Williams, D.L., Jr.; Feighner, S.D.; Nunes, C.N.; et al. Identification of receptors for neuromedin U and its role in feeding. Nature 2000, 406, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Qiao, X.; Crona, J.H.; Behan, J.; Wang, S.; Laz, T.; Bayne, M.; Gustafson, E.L.; Monsma, F.J., Jr.; Hedrick, J.A. Identification of a Novel Neuromedin U Receptor Subtype Expressed in the Central Nervous System. J. Biol. Chem. 2000, 275, 39482–39486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolasci, C.C.; Spolding, B.; Kidnapillai, S.; Connor, T.; Truong, T.T.T.; Liu, Z.S.J.; Panizzutti, B.; Richardson, M.F.; Gray, L.; Berk, M.; et al. Transcriptional Effects of Psychoactive Drugs on Genes Involved in Neurogenesis. Int. J. Mol. Sci. 2020, 21, 8333. [Google Scholar] [CrossRef]
- Taupin, P. Adult neurogenesis and neural stem cells as a model for the discovery and development of novel drugs. Expert Opin. Drug Discov. 2010, 5, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Couillard-Despres, S.; Wuertinger, C.; Kandasamy, M.; Caioni, M.; Stadler, K.; Aigner, R.; Bogdahn, U.; Aigner, L. Ageing abolishes the effects of fluoxetine on neurogenesis. Mol. Psychiatry 2009, 14, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodes, G.E.; Hill-Smith, T.E.; Suckow, R.F.; Cooper, T.B.; Lucki, I. Sex-specific effects of chronic fluoxetine treatment on neuroplasticity and pharmacokinetics in mice. J. Pharmacol. Exp. Ther. 2010, 332, 266–273. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Stumpel, M.W.; Wang, Q.; Aronica, E. Decreased numbers of progenitor cells but no response to antidepressant drugs in the hippocampus of elderly depressed patients. Neuropharmacology 2010, 58, 940–949. [Google Scholar] [CrossRef]
- Micheli, L.; Ceccarelli, M.; D’Andrea, G.; Tirone, F. Depression and adult neurogenesis: Positive effects of the antidepressant fluoxetine and of physical exercise. Brain Res. Bull. 2018, 143, 181–193. [Google Scholar] [CrossRef]
- Malberg, J.E.; Eisch, A.J.; Nestler, E.J.; Duman, R.S. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J. Neurosci. 2000, 20, 9104–9110. [Google Scholar] [CrossRef]
- Hunsberger, J.; Austin, D.R.; Hanter, I.D. The neurotrophic and neuroprotective effects of psychotropic agents. Dialogues Clin. Neurosci. 2009, 11, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, Z.; He, J.; Haimanot, S.; Li, X.; Dyck, L.; Li, X.M. Synergetic effects of quetiapine and venlafaxine in preventing the chronic restraint stressinduced decrease in cell proliferation and BDNF expression in rat hippocampus. Hippocampus 2006, 16, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Encinas, J.M.; Vaahtokari, A.; Enikolopov, G. Fluoxetine targets early progenitor cells in the adult brain. Proc. Natl. Acad. Sci. USA 2006, 103, 8233–8238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.V.; Shamy, J.L.; Bedi, G.; Choi, C.W.; Wall, M.M.; Arango, V.; Boldrini, M.; Foltin, R.W.; Hen, R. Impact of social status and antidepressant treatment on neurogenesis in the baboon hippocampus. Neuropsychopharmacology 2014, 39, 1861–1871. [Google Scholar] [CrossRef] [Green Version]
- Kohler, S.J.; Williams, N.I.; Stanton, G.B.; Cameron, J.L.; Greenough, W.T. Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months. Proc. Natl. Acad. Sci. USA 2011, 108, 10326–10331. [Google Scholar] [CrossRef] [Green Version]
- Li, W.L.; Cai, H.H.; Wang, B.; Chen, L.; Zhou, Q.G.; Luo, C.X.; Liu, N.; Ding, X.S.; Zhu, D.Y. Chronic fluoxetine treatment improves ischemia-induced spatial cognitive deficits through increasing hippocampal neurogenesis after stroke. J. Neurosci. Res. 2009, 87, 112–122. [Google Scholar] [CrossRef]
- Warkus, E.L.L.; Marikawa, Y. Fluoxetine Inhibits Canonical Wnt Signaling to Impair Embryoid Body Morphogenesis: Potential Teratogenic Mechanisms of a Commonly Used Antidepressant. Toxicol. Sci. 2018, 165, 372–388. [Google Scholar] [CrossRef]
- Wang, W.; Yin, H.; Feng, N.; Wang, L.; Wang, X. Inhibitory effects of antidepressant fluoxetine on cloned Kv2.1 potassium channel expressed in HEK293 cells. Eur. J. Pharmacol. 2020, 878, 173097. [Google Scholar] [CrossRef]
- Tate, K.; Kirk, B.; Tseng, A.; Ulffers, A.; Litwa, K. Effects of the Selective Serotonin Reuptake Inhibitor Fluoxetine on Developing Neural Circuits in a Model of the Human Fetal Cortex. Int. J. Mol. Sci. 2021, 22, 10457. [Google Scholar] [CrossRef]
- Molteni, R.; Calabrese, F.; Bedogni, F.; Tongiorgi, E.; Fumagalli, F.; Racagni, G.; Andrea Riva, M. Chronic treatment with fluoxetine up-regulates cellular BDNF mRNA expression in rat dopaminergic regions. Int. J. Neuropsychopharmacol. 2006, 9, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Outhred, T.; Das, P.; Felmingham, K.L.; Bryant, R.A.; Nathan, P.J.; Malhi, G.S.; Kemp, A.H. Impact of acute administration of escitalopram on the processing of emotional and neutral images: A randomized crossover fMRI study of healthy women. J. Psychiatry Neurosci. 2014, 39, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pringle, A.; Browning, M.; Cowen, P.J.; Harmer, C.J. A cognitive neuropsychological model of antidepressant drug action. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Chawana, R.; Alagaili, A.; Patzke, N.; Spocter, M.A.; Mohammed, O.B.; Kaswera, C.; Gilissen, E.; Bennett, N.C.; Ihunwo, A.O.; Manger, P.R. Microbats appear to have adult hippocampal neurogenesis, but post-capture stress causes a rapid decline in the number of neurons expressing doublecortin. Neuroscience 2014, 277, 724–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, N.; Habay, S.A.; Zeng, J.; Chamberlin, A.R.; Reinscheid, R.K. Synthesis and pharmacological in vitro and in vivo profile of 3-oxo-1,1-diphenyl-tetrahydro-oxazolo[3,4-a]pyrazine-7-carboxylic acid 4-fluoro-benzylamide (SHA68), a selective antagonist of the neuropeptide S receptor. J. Pharmacol. Exp. Ther. 2008, 325, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, H.; Liu, L.; Reinhart, C.; Michel, H. Heterologous expression of human neuromedin U receptor 1 and its subsequent solubilization and purification. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 2203–2209. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.K.; Reinhart, C.; Michel, H. Dimethylsulphoxide as a tool to increase functional expression of heterologously produced GPCRs in mammalian cells. FEBS Lett. 2006, 580, 4261–4265. [Google Scholar] [CrossRef] [Green Version]
- Cavaletti, G.; Oggioni, N.; Sala, F.; Pezzoni, G.; Cavalletti, E.; Marmiroli, P.; Petruccioli, M.G.; Frattola, L.; Tredici, G. Effect on the peripheral nervous system of systemically administered dimethylsulfoxide in the rat: A neurophysiological and pathological study. Toxicol. Lett. 2000, 118, 103–107. [Google Scholar] [CrossRef]
- Cavas, M.; Beltrán, D.; Navarro, J.F. Behavioural effects of dimethyl sulfoxide (DMSO): Changes in sleep architecture in rats. Toxicol. Lett. 2005, 157, 221–232. [Google Scholar] [CrossRef]
- Tamagnini, F.; Scullion, S.; Brown, J.T.; Randall, A.D. Low concentrations of the solvent dimethyl sulphoxide alter intrinsic excitability properties of cortical and hippocampal pyramidal cells. PLoS ONE 2014, 9, e92557. [Google Scholar] [CrossRef]
- Hanslick, J.L.; Lau, K.; Noguchi, K.K.; Olney, J.W.; Zorumski, C.F.; Mennerick, S.; Farber, N.B. Dimethyl sulfoxide (DMSO) produces widespread apoptosis in the developing central nervous system. Neurobiol. Dis. 2009, 34, 1–10. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, A.; Lange, S.; Rotheneichner, P.; Bieler, L.; Aigner, L.; Rivera, F.J.; Couillard-Despres, S. Dimethylsulfoxide Inhibits Oligodendrocyte Fate Choice of Adult Neural Stem and Progenitor Cells. Front. Neurosci. 2019, 26, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Authier, N.; Dupuis, E.; Kwasiborski, A.; Eschalier, A.; Coudoré, F. Behavioural assessment of dimethylsulfoxide neurotoxicity in rats. Toxicol. Lett. 2002, 32, 117–121. [Google Scholar] [CrossRef]
- Iwai, T.; Iinuma, Y.; Kodani, R.; Oka, J. Neuromedin U inhibits inflammation-mediated memory impairment and neuronal cell-death in rodents. Neurosci. Res. 2008, 61, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Telegdy, G. Neurotransmissions of antidepressant-like effects of neuromedin U-23 in mice. Behav. Brain Res. 2014, 259, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Telegdy, G.; Adamik, A. Anxiolytic action of neuromedin-U and neurotransmitters involved in mice. Regul. Pept. 2013, 186, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Vallöf, D.; Kalafateli, A.L.; Jerlhag, E. Brain region-specific neuromedin U signalling regulates alcohol-related behaviours and food intake in rodents. Addict. Biol. 2020, 25, e12764. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Gragerov, A.; Hohmann, J.G.; Pavlova, M.N.; Schimpf, B.A.; Xu, H.; Wu, L.J.; Toyoda, H.; Zhao, M.G.; Rohde, A.D.; et al. Neuromedin U receptor 2-deficient mice display differential responses in sensory perception, stress, and feeding. Mol. Cell. Biol. 2006, 26, 9352–9363. [Google Scholar] [CrossRef] [Green Version]
- Todd, T.P.; Bucci, D.J. Retrosplenial Cortex and Long-Term Memory: Molecules to Behavior. Neural Plast. 2015, 2015, 414173. [Google Scholar] [CrossRef] [Green Version]
- Agasse, F.; Bernardino, L.; Kristiansen, H.; Christiansen, S.H.; Ferreira, R.; Silva, B.; Grade, S.; Woldbye, D.P.; Malva, J.O. Neuropeptide Y promotes neurogenesis in murine subventricular zone. Stem Cells 2008, 26, 1636–1645. [Google Scholar] [CrossRef]
- Decressac, M.; Wright, B.; David, B.; Tyers, P.; Jaber, M.; Barker, R.A.; Gaillard, A. Exogenous neuropeptide Y promotes in vivo hippocampal neurogenesis. Hippocampus 2011, 21, 233–238. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: London, UK, 2007. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piwowarczyk-Nowak, A.; Pałasz, A.; Suszka-Świtek, A.; Błaszczyk, I.; Bogus, K.; Łasut-Szyszka, B.; Krzystanek, M.; Worthington, J.J. Effect of Escitalopram on the Number of DCX-Positive Cells and NMUR2 Receptor Expression in the Rat Hippocampus under the Condition of NPSR Receptor Blockade. Pharmaceuticals 2022, 15, 631. https://doi.org/10.3390/ph15050631
Piwowarczyk-Nowak A, Pałasz A, Suszka-Świtek A, Błaszczyk I, Bogus K, Łasut-Szyszka B, Krzystanek M, Worthington JJ. Effect of Escitalopram on the Number of DCX-Positive Cells and NMUR2 Receptor Expression in the Rat Hippocampus under the Condition of NPSR Receptor Blockade. Pharmaceuticals. 2022; 15(5):631. https://doi.org/10.3390/ph15050631
Chicago/Turabian StylePiwowarczyk-Nowak, Aneta, Artur Pałasz, Aleksandra Suszka-Świtek, Iwona Błaszczyk, Katarzyna Bogus, Barbara Łasut-Szyszka, Marek Krzystanek, and John J. Worthington. 2022. "Effect of Escitalopram on the Number of DCX-Positive Cells and NMUR2 Receptor Expression in the Rat Hippocampus under the Condition of NPSR Receptor Blockade" Pharmaceuticals 15, no. 5: 631. https://doi.org/10.3390/ph15050631
APA StylePiwowarczyk-Nowak, A., Pałasz, A., Suszka-Świtek, A., Błaszczyk, I., Bogus, K., Łasut-Szyszka, B., Krzystanek, M., & Worthington, J. J. (2022). Effect of Escitalopram on the Number of DCX-Positive Cells and NMUR2 Receptor Expression in the Rat Hippocampus under the Condition of NPSR Receptor Blockade. Pharmaceuticals, 15(5), 631. https://doi.org/10.3390/ph15050631