
The Role of Interventional Irisin on Heart Molecular Physiology

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. General Clinical Observations
Body Weights
2.2. CBC and Clinical Biochemistry
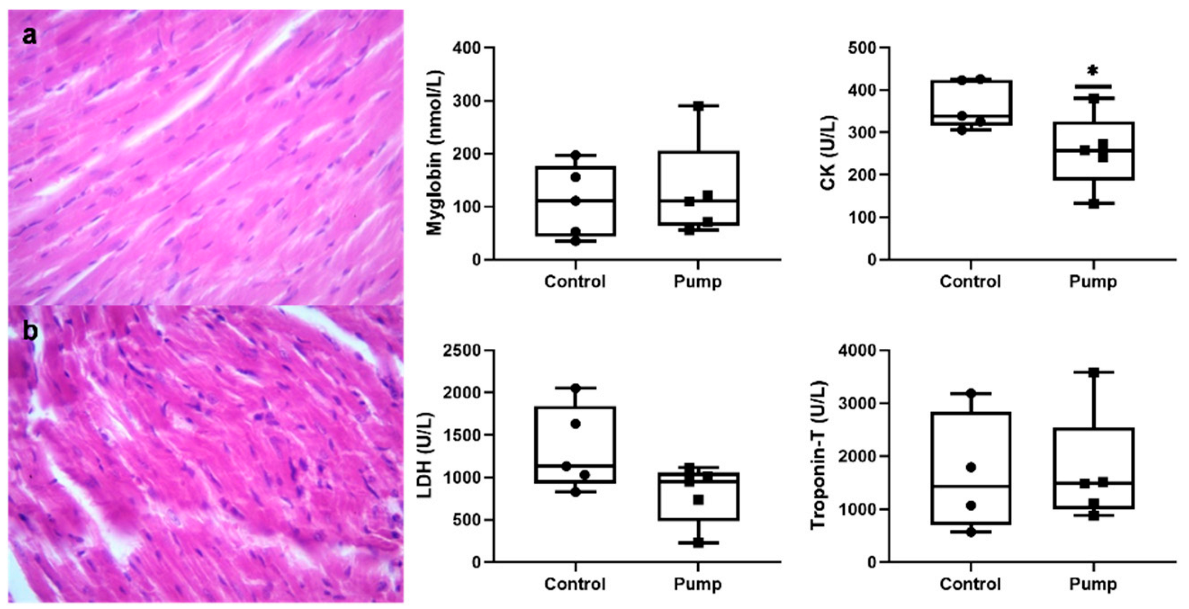
2.3. Histopathology
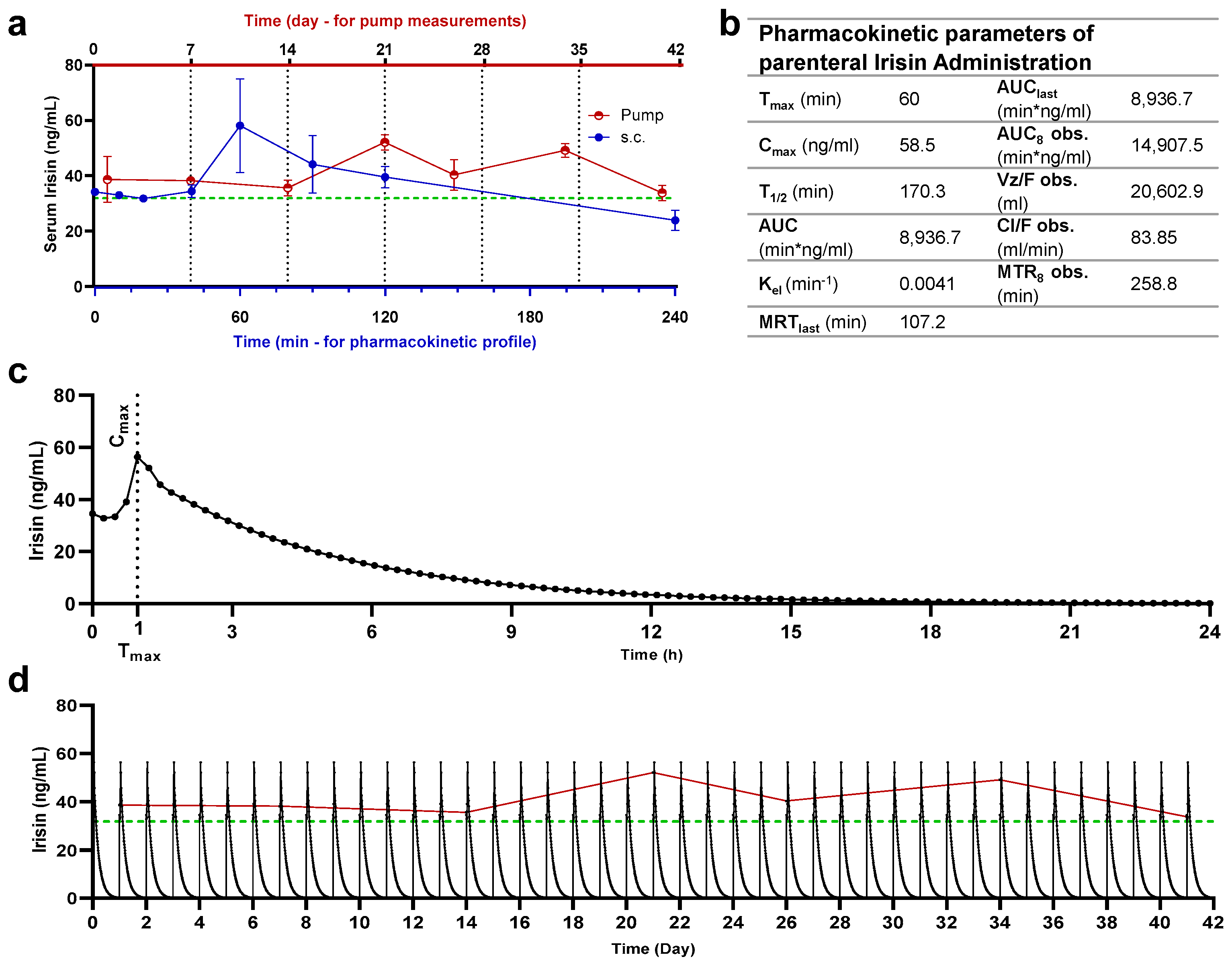
2.4. Quantification of Serum Irisin
2.5. Gene Expression Levels
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Pump Implantation and Irisin Administration
4.3. Parenteral Irisin Administration and Quantification
4.4. Sampling
4.5. Irisin Quantification
4.6. Superposition Principle Simulation Modelling
4.7. RNA Isolation and cDNA Synthesis
4.8. Relative Quantitative Real-Time PCR (RT-qPCR)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fagard, R. Athlete’s heart. Heart 2003, 89, 1455–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, B.J. Sudden Death in Young Athletes. N. Engl. J. Med. 2003, 349, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Morganroth, J.; Maron, B.J.; Henry, W.L.; Epstein, S.E. Comparative Left Ventricular Dimensions in Trained Athletes. Ann. Intern. Med. 1975, 82, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Bueno, O.F.; De Windt, L.J.; Tymitz, K.M.; Witt, S.A.; Kimball, T.R.; Klevitsky, R.; Hewett, T.E.; Jones, S.; Lefer, D.J.; Peng, C.; et al. The MEK1-ERK1/2 signaling pathway promotes compensated cardiac hypertrophy in transgenic mice. EMBO J. 2000, 19, 6341–6350. [Google Scholar] [CrossRef] [Green Version]
- Cittadini, A.; Strömer, H.; Katz, S.E.; Clark, R.; Moses, A.C.; Morgan, J.P.; Douglas, P.S. Differential Cardiac Effects of Growth Hormone and Insulin-like Growth Factor1 in the Rat. Circulation 1996, 93, 800–809. [Google Scholar] [CrossRef]
- Whitehead, N.; Gill, J.F.; Brink, M.; Handschin, C. Moderate Modulation of Cardiac PGC-1α Expression Partially Affects Age-Associated Transcriptional Remodeling of the Heart. Front. Physiol. 2018, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Hassaan, P.S.; Nassar, S.Z.; Issa, Y.; Zahran, N. Irisin vs. Treadmill Exercise in Post Myocardial Infarction Cardiac Rehabilitation in Rats. Arch. Med. Res. 2019, 50, 44–54. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463. [Google Scholar] [CrossRef]
- Spiegelman, B.M. Banting Lecture. Diabetes 2013, 62, 1774–1782. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Huh, J.Y.; Mougios, V.; Kabasakalis, A.; Fatouros, I.; Siopi, A.; Douroudos, I.I.; Filippaios, A.; Panagiotou, G.; Park, K.H.; Mantzoros, C.S. Exercise-Induced Irisin Secretion Is Independent of Age or Fitness Level and Increased Irisin May Directly Modulate Muscle Metabolism Through AMPK Activation. J. Clin. Endocrinol. Metab. 2014, 99, E2154–E2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, R.; Meng, Y.; Li, S.; Donelan, W.; Zhao, Y.; Qi, L.; Zhang, M.; Wang, X.; Cui, T.; et al. Irisin Stimulates Browning of White Adipocytes Through Mitogen-Activated Protein Kinase p38 MAP Kinase and ERK MAP Kinase Signaling. Diabetes 2014, 63, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, J.Y.; Mantzoros, C.S. Irisin physiology, oxidative stress, and thyroid dysfunction: What next? Metabolism 2015, 64, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M. Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [CrossRef]
- Majeed, S.; Shafi, R.; Moin, H.; Ashraf, I.; Irshad, K.; Liaquat, A. Effects of Recombinant Irisin on Body Mass Index, Serum Insulin, Luteinizing Hormone and Testosterone Levels in Obese Female BALB/c Mice. J. Coll. Physicians Surg. Pak. 2019, 29, 736–740. [Google Scholar] [CrossRef]
- Asadi, Y.; Gorjipour, F.; Behrouzifar, S.; Vakili, A. Irisin Peptide Protects Brain Against Ischemic Injury Through Reducing Apoptosis and Enhancing BDNF in a Rodent Model of Stroke. Neurochem. Res. 2018, 43, 1549–1560. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, X.; Xu, P.; Mi, J.-Y.; Rui, Y.-J. The protective effect of Irisin against ischemia-reperfusion injury after perforator flap grafting in rats. Injury 2018, 49, 2147–2153. [Google Scholar] [CrossRef]
- Duan, H.; Ma, B.; Ma, X.; Wang, H.; Ni, Z.; Wang, B.; Li, X.; Jiang, P.; Umar, M.; Li, M. Anti-diabetic activity of recombinant irisin in STZ-induced insulin-deficient diabetic mice. Int. J. Biol. Macromol. 2016, 84, 457–463. [Google Scholar] [CrossRef]
- Aydin, S.; Kuloglu, T.; Aydin, S.; Eren, M.N.; Celik, A.; Yilmaz, M.; Kalayci, M.; Sahin, I.; Gungor, O.; Gurel, A.; et al. Cardiac, skeletal muscle and serum irisin responses to with or without water exercise in young and old male rats: Cardiac muscle produces more irisin than skeletal muscle. Peptides 2014, 52, 68–73. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Guo, L.; Jiang, J.; Chen, H. Irisin: Linking metabolism with heart failure. Am. J. Transl. Res. 2020, 12, 6003–6014. [Google Scholar]
- Alzoughool, F.; Al-Zghoul, M.B.; Al-Nassan, S.; Alanagreh, L.; Mufleh, D.; Atoum, M. The optimal therapeutic irisin dose intervention in animal model: A systematic review. Vet. World 2020, 13, 2191–2196. [Google Scholar] [CrossRef] [PubMed]
- Bashar, S.M.; El-Sherbeiny, S.M.S.; Boraie, M.Z. Correlation between the blood level of irisin and the severity of acute myocardial infarction in exercise-trained rats. J. Basic Clin. Physiol. Pharmacol. 2019, 30, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Valaris, S.; Young, M.F.; Haley, E.B.; Luo, R.; Bond, S.F.; Mazuera, S.; Kitchen, R.R.; Caldarone, B.J.; Bettio, L.E.B.; et al. Exercise hormone irisin is a critical regulator of cognitive function. Nat. Metab. 2021, 3, 1058–1070, Correction in Nat. Metab. 2021, 3, 1432. [Google Scholar] [CrossRef]
- Fu, J.; Li, F.; Tang, Y.; Cai, L.; Zeng, C.; Yang, Y.; Yang, J. The Emerging Role of Irisin in Cardiovascular Diseases. J. Am. Heart Assoc. 2021, 10, e022453. [Google Scholar] [CrossRef] [PubMed]
- Ou-Yang, W.-L.; Guo, B.; Xu, F.; Lin, X.; Li, F.-X.; Shan, S.-K.; Wu, F.; Wang, Y.; Zheng, M.-H.; Xu, Q.-S.; et al. The Controversial Role of Irisin in Clinical Management of Coronary Heart Disease. Front. Endocrinol. 2021, 12, 678309. [Google Scholar] [CrossRef]
- Aydin, S. Is irisin a decisive protein in cancer cachexia and death of cancer cells? Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3727–3729. [Google Scholar]
- Welch, S.; Plank, D.; Witt, S.; Glascock, B.; Schaefer, E.; Chimenti, S.; Andreoli, A.M.; Limana, F.; Leri, A.; Kajstura, J.; et al. Cardiac-specific IGF-1 expression attenuates dilated cardiomyopathy in tropomodulin-overexpressing transgenic mice. Circ. Res. 2002, 90, 641–648. [Google Scholar] [CrossRef] [Green Version]
- González-Guerra, J.L.; Castilla-Cortazar, I.; Aguirre, G.A.; Muñoz, U.; Martin-Estal, I.; Ávila-Gallego, E.; Granado, M.; Puche, J.E.; García-Villalón, A.L. Partial IGF-1 deficiency is sufficient to reduce heart contractibility, angiotensin II sensibility, and alter gene expression of structural and functional cardiac proteins. PLoS ONE 2017, 12, e0181760. [Google Scholar] [CrossRef] [Green Version]
- Parra, V.; Rothermel, B.A. Calcineurin signaling in the heart: The importance of time and place. J. Mol. Cell. Cardiol. 2017, 103, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Chaklader, M.; Rothermel, B.A. Calcineurin in the heart: New horizons for an old friend. Cell. Signal. 2021, 87, 110134. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta 2016, 1863, 2422–2435. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid β-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2010, 1801, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Mikula, M.; Schreiber, M.; Husak, Z.; Kucerova, L.; Rüth, J.; Wieser, R.; Zatloukal, K.; Beug, H.; Wagner, E.F.; Baccarini, M. Embryonic lethality and fetal liver apoptosis in mice lacking the c-raf-1 gene. EMBO J. 2001, 20, 1952–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huser, M.; Luckett, J.; Chiloeches, A.; Mercer, K.; Iwobi, M.; Giblett, S.; Sun, X.; Brown, J.; Marais, R.; Pritchard, C. MEK kinase activity is not necessary for Raf-1 function. EMBO J. 2001, 20, 1940–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, O.; Watanabe, T.; Nishida, K.; Kashiwase, K.; Higuchi, Y.; Takeda, T.; Hikoso, S.; Hirotani, S.; Asahi, M.; Taniike, M.; et al. Cardiac-specific disruption of the c-raf-1 gene induces cardiac dysfunction and apoptosis. J. Clin. Investig. 2004, 114, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushima, S.; Kuroda, J.; Ago, T.; Zhai, P.; Ikeda, Y.; Oka, S.; Fong, G.-H.; Tian, R.; Sadoshima, J. Broad Suppression of NADPH Oxidase Activity Exacerbates Ischemia/Reperfusion Injury Through Inadvertent Downregulation of Hypoxia-inducible Factor-1α and Upregulation of Peroxisome Proliferator–activated Receptor-α. Circ. Res. 2013, 112, 1135–1149. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, F.L.M.; Walum, E.; Wikström, P.; Arner, A. A small molecule inhibitor of Nox2 and Nox4 improves contractile function after ischemia–reperfusion in the mouse heart. Sci. Rep. 2021, 11, 11970. [Google Scholar] [CrossRef]
- Hall, A.R.; Burke, N.; Dongworth, R.K.; Kalkhoran, S.B.; Dyson, A.; Vicencio, J.M.; Dorn, G.W.; Yellon, D.; Hausenloy, D.J. Hearts deficient in both Mfn1 and Mfn2 are protected against acute myocardial infarction. Cell Death Dis. 2016, 7, e2238. [Google Scholar] [CrossRef]
- Dinas, P.C.; Lahart, I.M.; Timmons, J.A.; Svensson, P.A.; Koutedakis, Y.; Flouris, A.D.; Metsios, G.S. Effects of physical activity on the link between PGC-1a and FNDC5 in muscle, circulating Ιrisin and UCP1 of white adipocytes in humans: A systematic review. F1000Research 2017, 6, 286. [Google Scholar] [CrossRef]
- Karamanlidis, G.; Garcia-Menendez, L.; Kolwicz, S.C., Jr.; Lee, C.F.; Tian, R. Promoting PGC-1α-driven mitochondrial biogenesis is detrimental in pressure-overloaded mouse hearts. Am. J. Physiol. Circ. Physiol. 2014, 307, H1307–H1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, I.C.; Ho, M.Y.; Wen, M.S.; Chen, C.C.; Hsieh, M.J.; Lin, C.P.; Yeh, J.K.; Tsai, M.L.; Yang, C.H.; Wu, V.C.C.; et al. Serum Irisin Levels Are Associated with Adverse Cardiovascular Outcomes in Patients with Acute Myocardial Infarction. Int. J. Cardiol. 2018, 261, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Khorasani, Z.M.; Bagheri, R.K.; Yaghoubi, M.A.; Chobkar, S.; Aghaee, M.A.; Abbaszadegan, M.R.; Sahebkar, A. The Association between Serum Irisin Levels and Cardiovascular Disease in Diabetic Patients. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Guo, Y.; Xia, Y.; Li, C.; Xu, X.; Qi, T.; Zhang, F.; Fan, M.; Hu, G.; Zhao, H.; et al. FNDC5/Irisin Attenuates Diabetic Cardiomyopathy in a Type 2 Diabetes Mouse Model by Activation of Integrin AV/Β5-AKT Signaling and Reduction of Oxidative/Nitrosative Stress. J. Mol. Cell Cardiol. 2021, 160, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Xin, C.; Zhang, Z.; Gao, G.; Ding, L.; Yang, C.; Wang, C.; Liu, Y.; Guo, Y.; Yang, X.; Zhang, L.; et al. Irisin Attenuates Myocardial Ischemia/Reperfusion Injury and Improves Mitochondrial Function Through AMPK Pathway in Diabetic Mice. Front. Pharmacol. 2020, 11, 1374. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, X.; Wang, F.; Ye, Q.; Yang, C.; Liu, D. Irisin Rescues Diabetic Cardiac Microvascular Injury via ERK1/2/Nrf2/HO-1 Mediated Inhibition of Oxidative Stress. Diabetes Res. Clin. Pract. 2022, 183, 109170. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Irisin-Pump | ||
|---|---|---|---|
| Complete blood count (CBC) | Leukocytes (WBCs) Count (×103) | 2.98 ± 0.35 | 2.34 ± 0.41 |
| Red blood cells (RBCs) Count (×106) | 5.92 ± 0.95 | 5.60 ± 0.68 | |
| Hemoglobin (g/dL) | 12.84 ± 0.52 | 13.08 ± 0.41 | |
| Hematocrit % | 33.8 ± 5.95 | 31.36 ± 4.51 | |
| Platelets Count (PCT) (×103) | 717 ± 54.58 | 781.60 ± 208.77 | |
| PCT % | 0.53 ± 0.09 | 0.58 ± 0.08 | |
| Mean cell volume (MCV) (μm3) | 55.8 ± 1.93 | 55.20 ± 1.83 | |
| Mean cell hemoglobin (MCH) (pg) | 26.22 ± 7.25 | 24.96 ± 3.42 | |
| Mean cell hemoglobin concentration (MCHC) (g/dL) | 48.8 ± 15.83 | 45.98 ± 7.74 | |
| Red cell distribution width (RDW) % | 17.22 ± 2.43 | 18.22 ± 2.58 | |
| Mean platelet volume (MPV) (μm3) | 7.44 ± 1.10 | 8.50 ± 1.49 | |
| Platelet distribution width (PDW) % | 13.22 ± 0.33 | 9.34 ± 1.46 * | |
| Lymphocytes % | 86.42 ± 1.11 | 82.98 ± 1.05 | |
| Monocytes % | 10.32 ± 0.67 | 11.80 ± 0.93 | |
| Granulocyte % | 3.26 ± 0.48 | 5.22 ± 0.24 | |
| Lymphocytes Count (×103) | 2.52 ± 0.31 | 1.88 ± 0.37 | |
| Monocytes Count (×103) | 0.26 ± 0.04 | 0.24 ± 0.04 * | |
| Granulocyte Count (×103) | 0.20 ± 0.00 | 0.22 ± 0.02 | |
| Clinical Biochemistry | |||
| Cholesterol (mg/dL) | 59.8 ± 5.3 | 67.8 ± 5.6 | |
| Triglycerides (mg/dL) | 53 ± 2.4 | 51.2 ± 7.3 | |
| High-Density Lipoprotein (HDL) (mmol/L) | 16.2 ± 1.7 | 18.6 ± 1.2 | |
| Low-Density Lipoprotein (LDL) (mmol/L) | 33.4 ± 3.9 | 39 ± 3.1 | |
| Cholesterol/HDL Ratio | 3.8 ± 0.4 | 3.62 ± 0.1 | |
| Very Low-Density Lipoprotein (VLDL) (U/L) | 10.2 ± 0.6 | 9 ± 2.1 | |
| Aspartate-aminotransferase (AST:GOT) (U/L) | 105 ± 3.7 | 96.4 ± 7.4 | |
| Alanine-aminotransferase (ALT:GPT) (U/L) | 24.6 ± 1.3 | 26 ± 2.1 | |
| Alkaline Phosphatase (ALP) (U/L) | 26.2 ± 5 | 30 ± 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzoughool, F.; Al-Zghoul, M.B.; Ghanim, B.Y.; Gollob, M.; Idkaidek, N.; Qinna, N.A. The Role of Interventional Irisin on Heart Molecular Physiology. Pharmaceuticals 2022, 15, 863. https://doi.org/10.3390/ph15070863
Alzoughool F, Al-Zghoul MB, Ghanim BY, Gollob M, Idkaidek N, Qinna NA. The Role of Interventional Irisin on Heart Molecular Physiology. Pharmaceuticals. 2022; 15(7):863. https://doi.org/10.3390/ph15070863
Chicago/Turabian StyleAlzoughool, Foad, Mohammad Borhan Al-Zghoul, Bayan Y. Ghanim, Michael Gollob, Nasir Idkaidek, and Nidal A. Qinna. 2022. "The Role of Interventional Irisin on Heart Molecular Physiology" Pharmaceuticals 15, no. 7: 863. https://doi.org/10.3390/ph15070863
APA StyleAlzoughool, F., Al-Zghoul, M. B., Ghanim, B. Y., Gollob, M., Idkaidek, N., & Qinna, N. A. (2022). The Role of Interventional Irisin on Heart Molecular Physiology. Pharmaceuticals, 15(7), 863. https://doi.org/10.3390/ph15070863