
Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Isolates
2.2. Phenotypic Antimicrobial Susceptibility Testing
2.3. Genomic Sequence Features and Strains Typing
2.4. Detection of Antibiotic-Resistance Genes (ARGs)
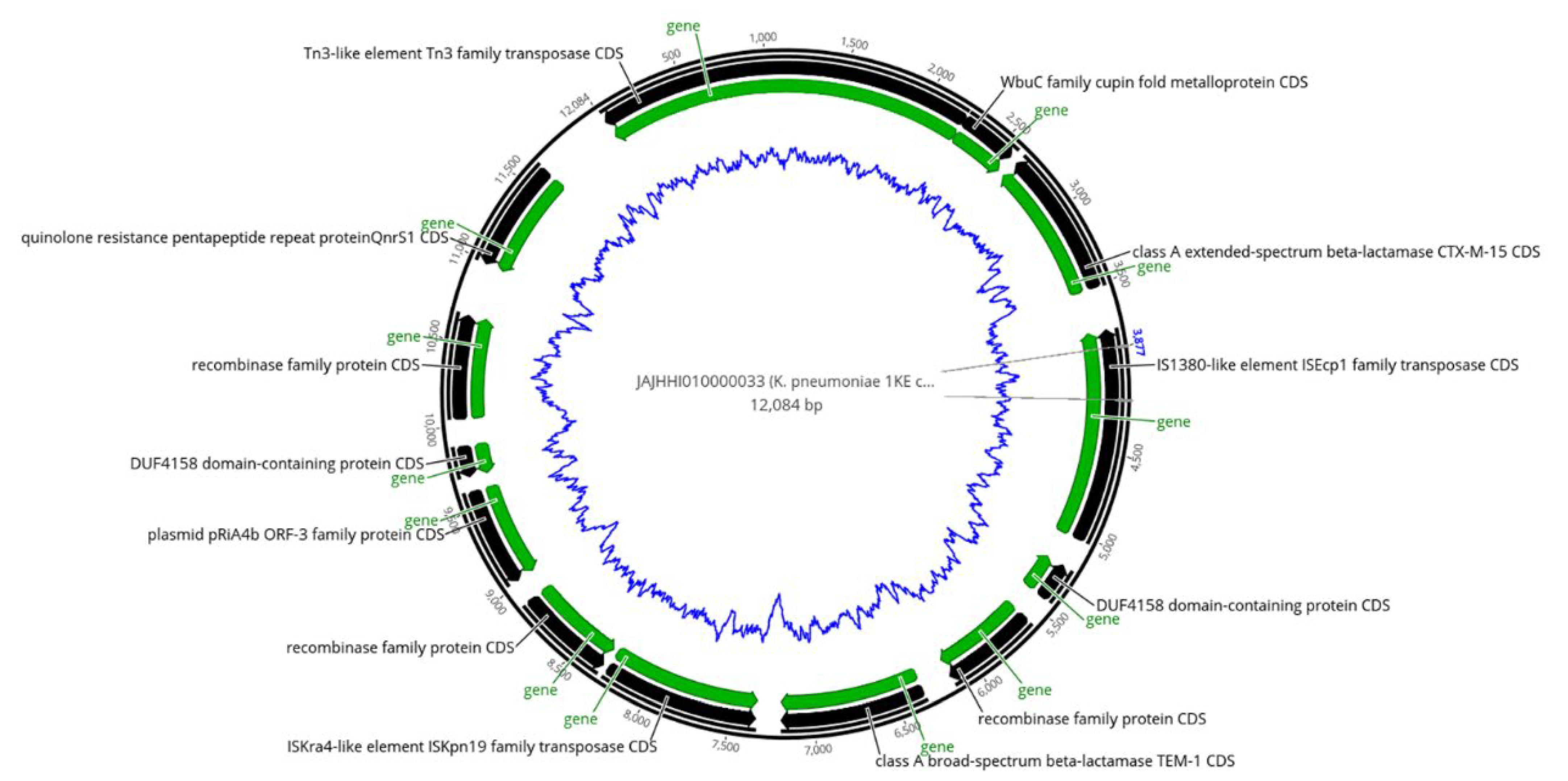
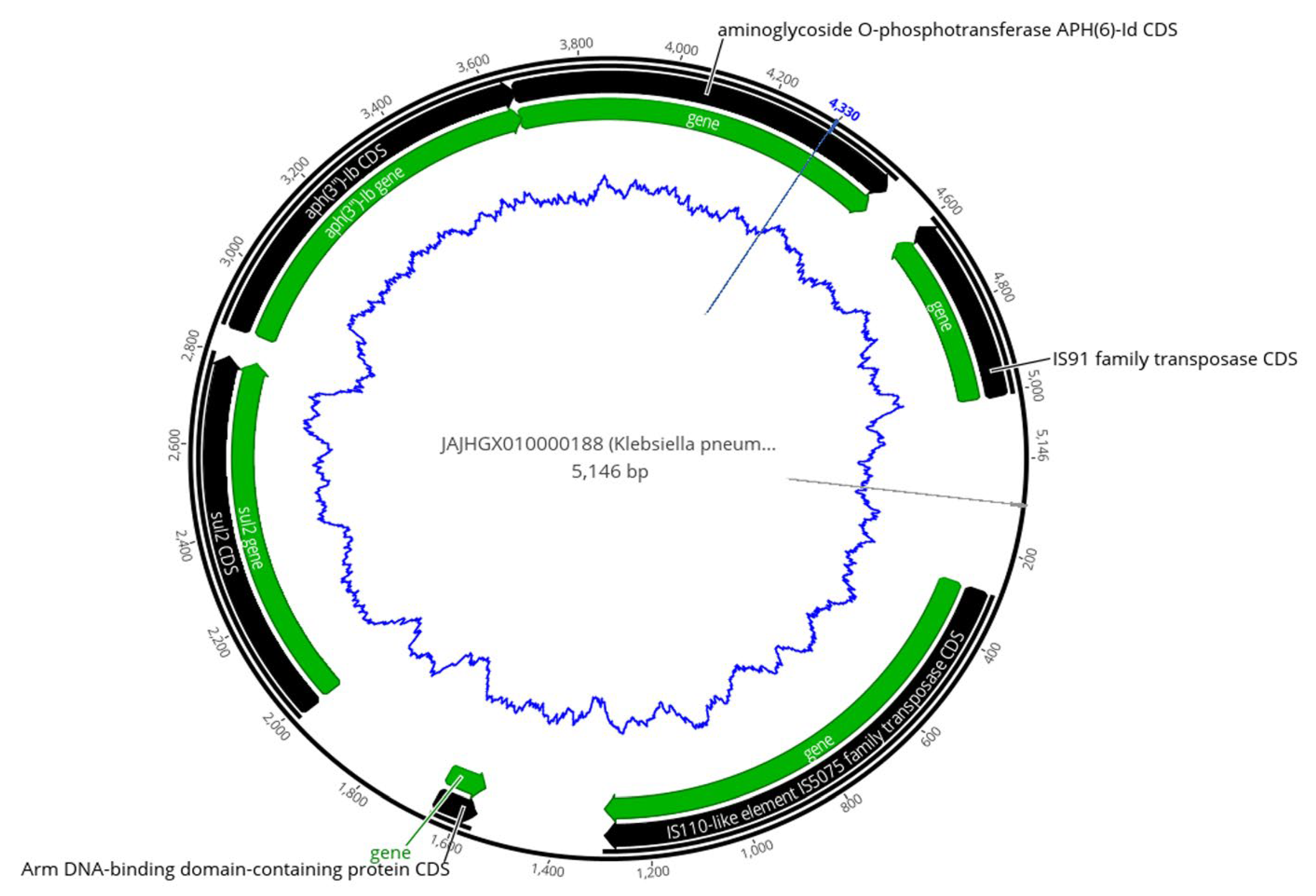
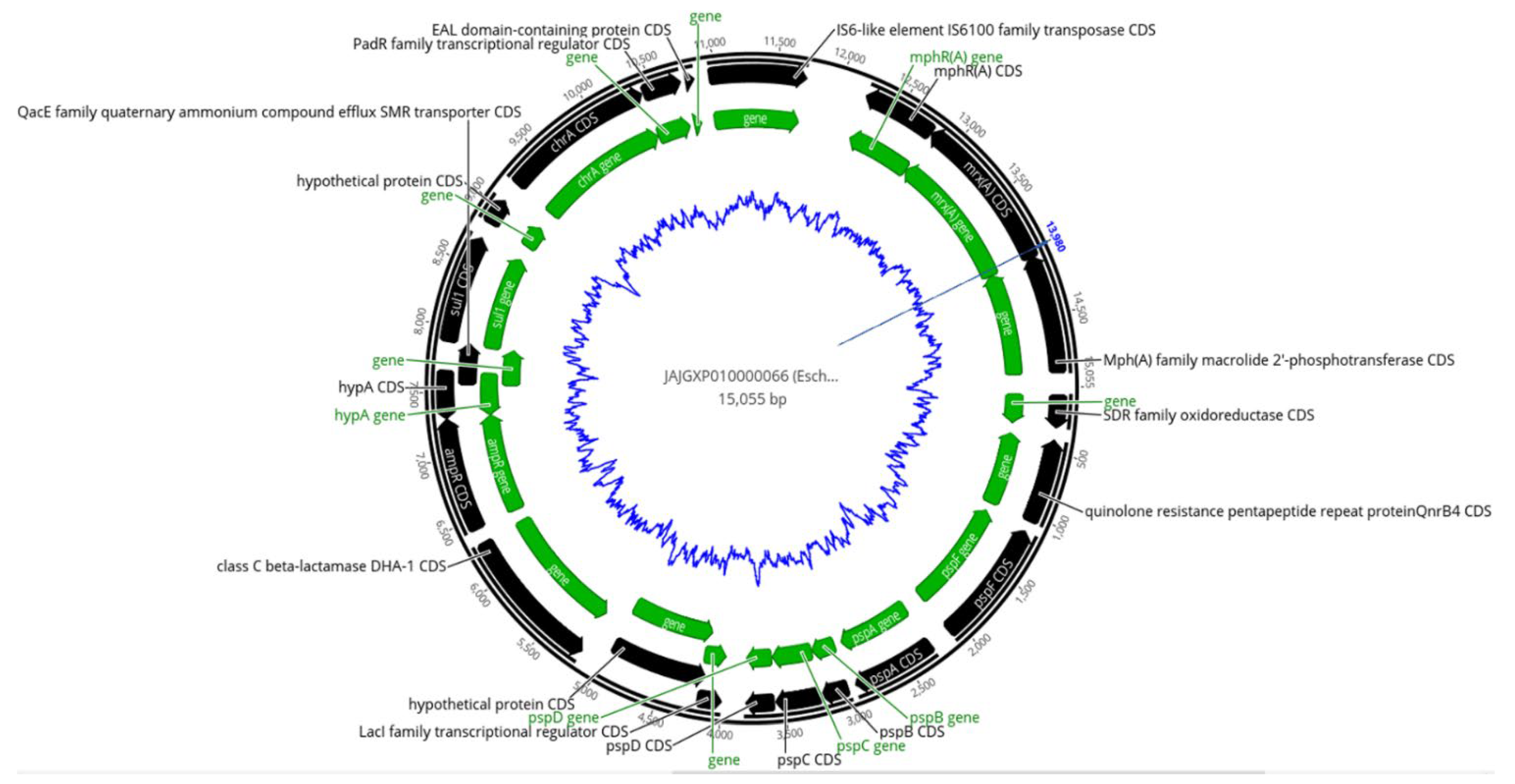
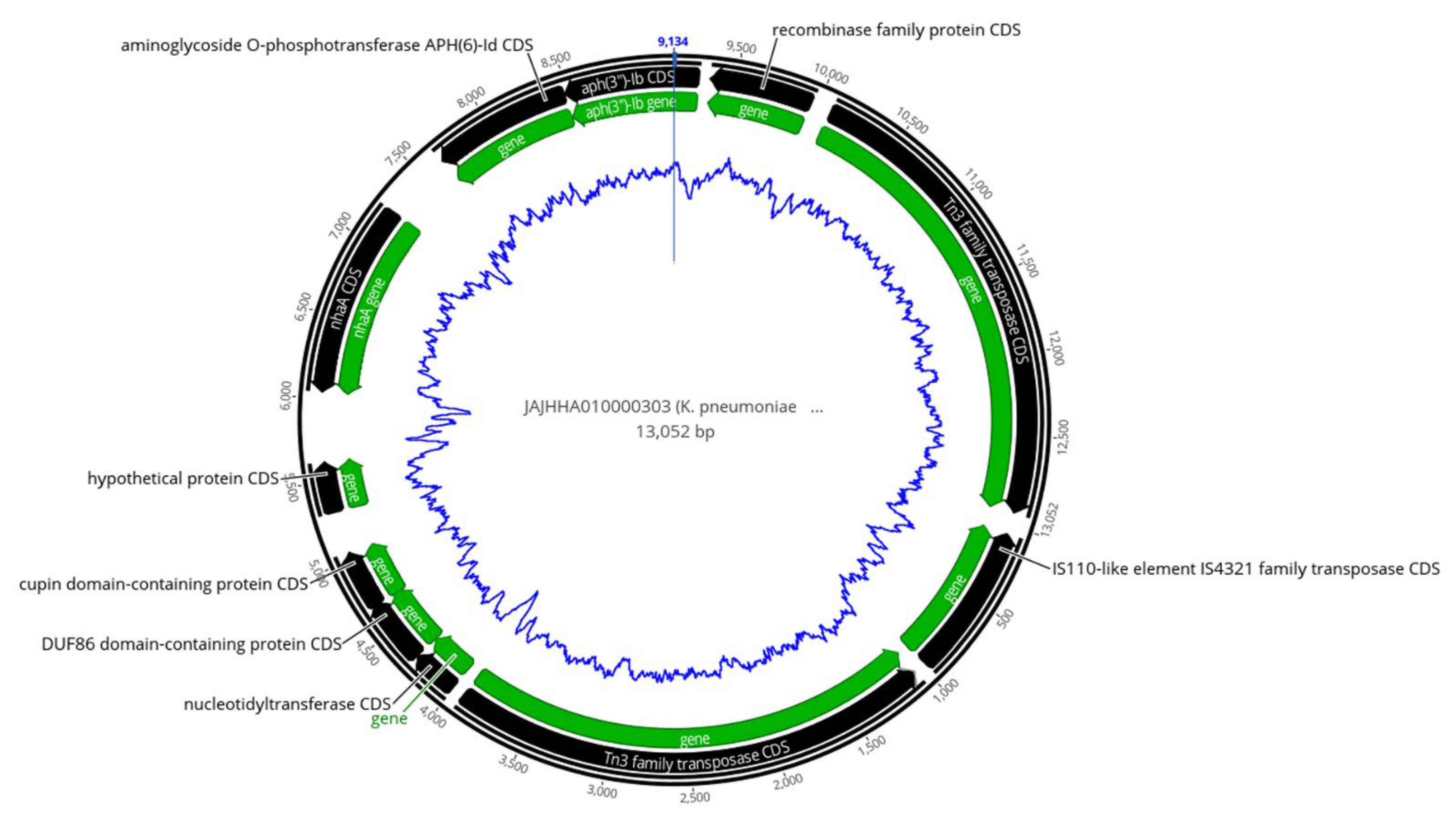
2.5. Analysis of Mobile Genetic Elements (MGEs)
2.6. Co-Occurrence of ARGs with Transposases
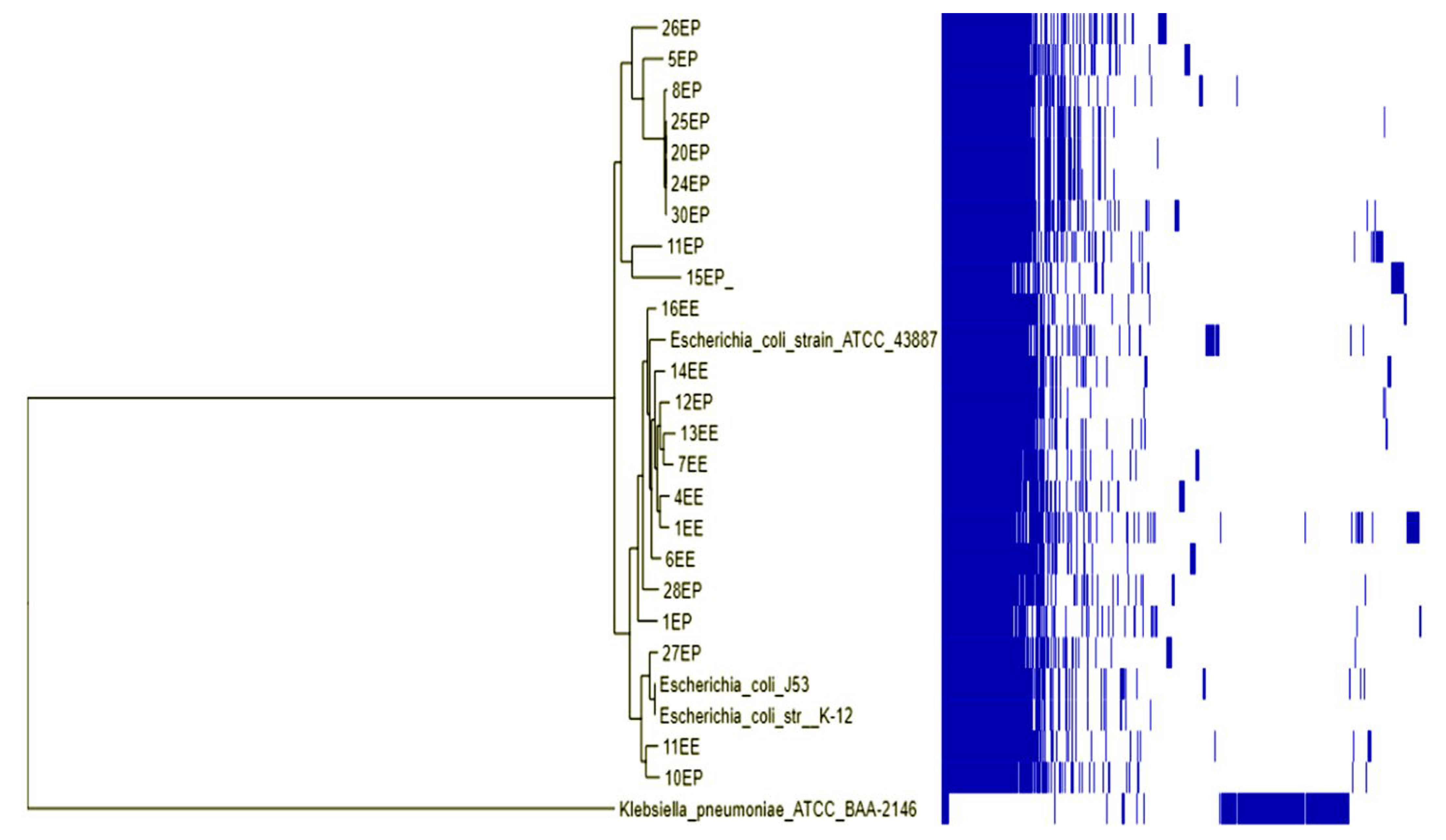
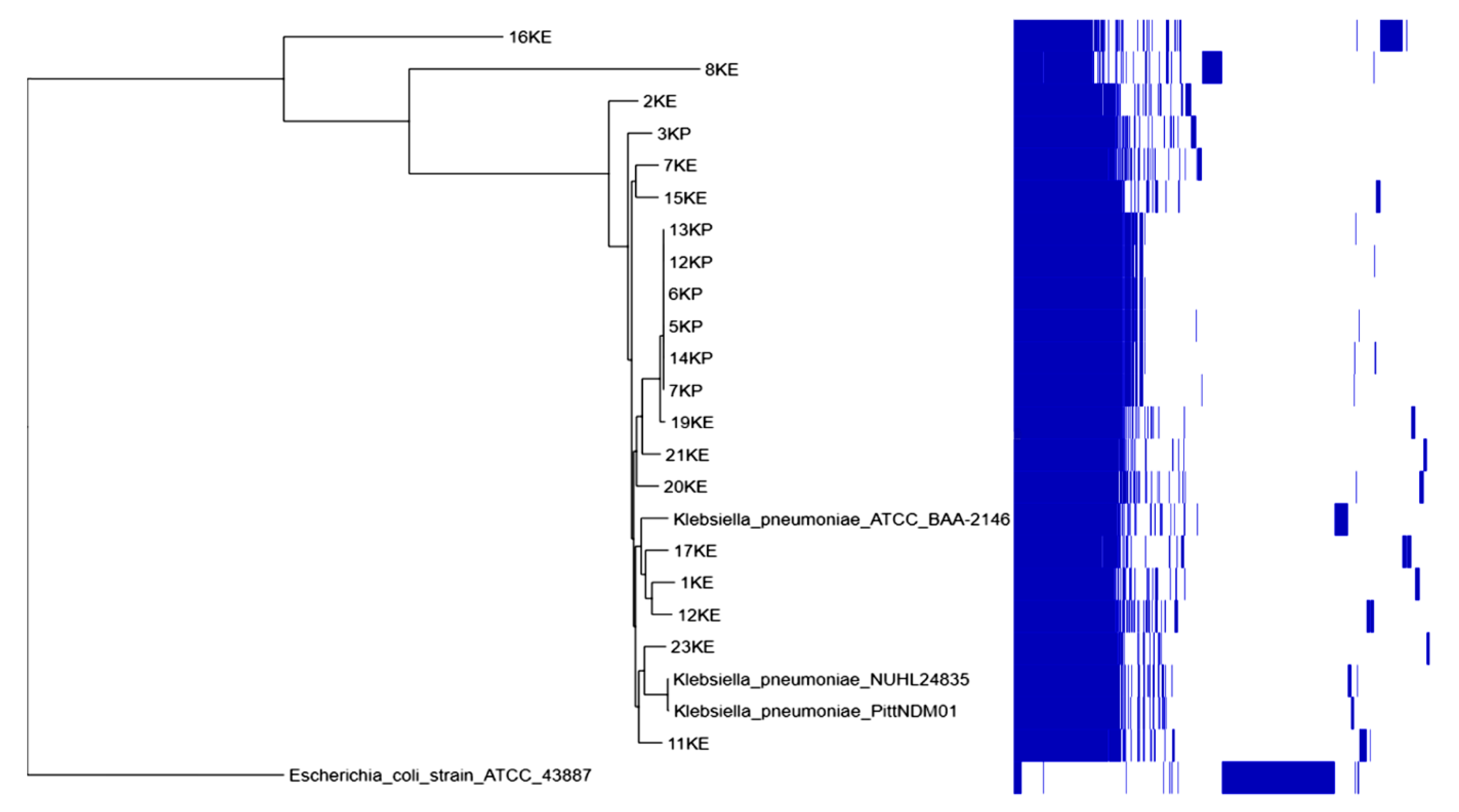
2.7. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Antimicrobial Susceptibility Testing
4.2. Whole-Genome Sequencing and Molecular Analysis
4.3. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Tacconelli, E.; Magrini, N.; Kahlmeter, G.; Singh, N. Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. World Health Organ. 2017, 27, 318–327. [Google Scholar]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.R.; Griffith, M.P.; Sundermann, A.J.; Shutt, K.A.; Saul, M.I.; Mustapha, M.M.; Marsh, J.W.; Cooper, V.S.; Harrison, L.H.; Van Tyne, D. Systematic detection of horizontal gene transfer across genera among multidrug-resistant bacteria in a single hospital. Elife 2020, 9, e53886. [Google Scholar] [CrossRef] [PubMed]
- Xanthopoulou, K.; Carattoli, A.; Wille, J.; Biehl, L.M.; Rohde, H.; Farowski, F.; Krut, O.; Villa, L.; Feudi, C.; Seifert, H. Antibiotic Resistance and Mobile Genetic Elements in Extensively Drug-Resistant Klebsiella pneumoniae Sequence Type 147 Recovered from Germany. Antibiotics 2020, 9, 675. [Google Scholar] [CrossRef]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Ahmed, A.; Sirag, B.; Raees, F.; Kidir, E.-S.; Ali, T.; Rahman, M.A.; Ashgar, S.; Barhameen, A.; Elsawy, A.; Mostafa, A. Bacterial whole genome sequencing as powerful tool for hospital molecular epidemiology: Acinetobacter baumannii as a model. Clin. Microbiol. Infect. Dis. 2016, 1, 10–16. [Google Scholar] [CrossRef]
- Stadler, T.; Meinel, D.; Aguilar-Bultet, L.; Huisman, J.S.; Schindler, R.; Egli, A.; Seth-Smith, H.M.; Eichenberger, L.; Brodmann, P.; Hübner, P. Transmission of ESBL-producing Enterobacteriaceae and their mobile genetic elements—Identification of sources by whole genome sequencing: Study protocol for an observational study in Switzerland. BMJ Open 2018, 8, e021823. [Google Scholar] [CrossRef]
- Palzkill, T. Structural and mechanistic basis for extended-spectrum drug-resistance mutations in altering the specificity of TEM, CTX-M, and KPC β-lactamases. Front. Mol. Biosci. 2018, 5, 16. [Google Scholar] [CrossRef]
- Elbadawi, H.S.; Elhag, K.M.; Mahgoub, E.; Altayb, H.N.; Ntoumi, F.; Elton, L.; McHugh, T.D.; Tembo, J.; Ippolito, G.; Osman, A.Y. Detection and characterization of carbapenem resistant Gram-negative bacilli isolates recovered from hospitalized patients at Soba University Hospital, Sudan. BMC Microbiol. 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Osman, E.A.; El-Amin, N.I.; Al-Hassan, L.L.; Mukhtar, M. Multiclonal spread of Klebsiella pneumoniae across hospitals in Khartoum, Sudan. J. Glob. Antimicrob. Resist. 2021, 24, 241–245. [Google Scholar] [CrossRef]
- Mohamed, S.B.; Kambal, S.; Munir, A.; Abdalla, N.; Hassan, M.; Hamad, A.; Mohammed, S.; Ahmed, F.; Hamid, O.; Ismail, A. First Whole-Genome Sequence of a Highly Resistant Klebsiella pneumoniae Sequence Type 14 Strain Isolated from Sudan. Microbiol. Resour. Announc. 2019, 8, e00552-19. [Google Scholar] [CrossRef]
- Mbanga, J.; Amoako, D.G.; Abia, A.L.; Allam, M.; Ismail, A.; Essack, S.Y. Genomic Insights of Multidrug-Resistant Escherichia coli From Wastewater Sources and Their Association With Clinical Pathogens in South Africa. Front. Vet. Sci. 2021, 8, 137. [Google Scholar] [CrossRef]
- Madni, O.; Amoako, D.G.; Abia, A.L.K.; Rout, J.; Essack, S.Y. Genomic Investigation of Carbapenem-Resistant Klebsiella pneumonia Colonization in an Intensive Care Unit in South Africa. Genes 2021, 12, 951. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.; Fischer, J.; Wagenaar, J.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Mayor, S. Doctors to get real time data to support antibiotic prescribing and reduce resistance. BMJ 2019, 25, l406. [Google Scholar] [CrossRef]
- Balali, G.I.; Yar, D.D.; Dela, V.G.A.; Adjei-Kusi, P. Microbial contamination, an increasing threat to the consumption of fresh fruits and vegetables in today’s world. Int. J. Microbiol. 2020, 2020, 3029295. [Google Scholar] [CrossRef]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic pollution in the environment: From microbial ecology to public policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H. Antibiotic resistance—The need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Van Duin, D.; Paterson, D.L. Multidrug-resistant bacteria in the community: Trends and lessons learned. Infect. Dis. Clin. 2016, 30, 377–390. [Google Scholar] [CrossRef]
- Elhadary, Y.; Ali, S. A new trend in urban housing: Gated communities in Khartoum, Sudan. Am. J. Sociol. Res. 2017, 7, 45–55. [Google Scholar]
- Gatari, M.J. Air pollution over East Africa. In Proceedings of the First International Workshop on Climate Variability over Africa, Alexandria, Egypt, 15–26 May 2005. [Google Scholar]
- Kheder, S.I. Physcians knowledge and perception of antimicrobial resistance: A survey in Khartoum Stata Hospital settings. Br. J. Pharmaceut. Res. 2013, 3, 347–362. [Google Scholar] [CrossRef]
- Musa, M.M.Y.A. Medicine Prices, Availability and Affordability in Sudan. In Partial Fulfillment of Master of Sciences of Health Economics and Health Care Management; Chulalongkorn University: Bangkok, Thailand, 2013. [Google Scholar]
- Alfadil, A.; Ahmed, N.; Mohamed, M.S.; Ali, M.M.; El Nima, E.A.I. Characterization of pathogenic bacteria isolated from Sudanese banknotes and determination of their resistance profile. Int. J. Microbiol. 2018, 2018, 4375164. [Google Scholar] [CrossRef]
- Mohammed, S.A.S.; Musa, A.; Mohammed, A.A.; Mohammed, H. CTX-M B-lactamase–producing Escherichia coli in Sudan tertiary hospitals: Detection genotypes variants and bioinformatics analysis. Int. J. Med. Biomed. Stud. 2019, 3, 146–157. [Google Scholar]
- Altayb, H.N.; Salih, E.K.; Moglad, E.H. Molecular detection of beta-lactamase blaCTX-M group 1 in Escherichia coli isolated from drinking water in Khartoum State. J. Water Health 2020, 18, 1091–1097. [Google Scholar] [CrossRef]
- Runcharoen, C.; Moradigaravand, D.; Blane, B.; Paksanont, S.; Thammachote, J.; Anun, S.; Parkhill, J.; Chantratita, N.; Peacock, S.J. Whole genome sequencing reveals high-resolution epidemiological links between clinical and environmental Klebsiella pneumoniae. Genome Med. 2017, 9, 1–10. [Google Scholar] [CrossRef]
- Malik, I.; Elhag, K.; Gustaw, K. Characterisation of extended-spectrum β-lactamases among multidrug resistant Enterobacteriaceae from Sudan. J. Pure Appl. Microbiol. 2019, 13, 8. [Google Scholar]
- Ahmed, O.B.; Omar, A.O.; Asghar, A.H.; Elhassan, M.M.; Al-Munawwarah, A.-M.; Arabia, S. Prevalence of TEM, SHV and CTX-M genes in Escherichia coli and Klebsiella spp Urinary Isolates from Sudan with confirmed ESBL phenotype. Life Sci. J. 2013, 10, 191–195. [Google Scholar]
- Bougnom, B.; Thiele-Bruhn, S.; Ricci, V.; Zongo, C.; Piddock, L. Raw wastewater irrigation for urban agriculture in three African cities increases the abundance of transferable antibiotic resistance genes in soil, including those encoding extended spectrum β-lactamases (ESBLs). Sci. Total Environ. 2020, 698, 134201. [Google Scholar] [CrossRef]
- Yaici, L.; Haenni, M.; Métayer, V.; Saras, E.; Zekar, F.M.; Ayad, M.; Touati, A.; Madec, J.-Y. Spread of ESBL/AmpC-producing Escherichia coli and Klebsiella pneumoniae in the community through ready-to-eat sandwiches in Algeria. Int. J. Food Microbiol. 2017, 245, 66–72. [Google Scholar] [CrossRef]
- Moremi, N.; Manda, E.V.; Falgenhauer, L.; Ghosh, H.; Imirzalioglu, C.; Matee, M.; Chakraborty, T.; Mshana, S.E. Predominance of CTX-M-15 among ESBL producers from environment and fish gut from the shores of Lake Victoria in Mwanza, Tanzania. Front. Microbiol. 2016, 7, 1862. [Google Scholar] [CrossRef]
- Ojer-Usoz, E.; González, D.; Vitas, A.I. Clonal diversity of ESBL-producing Escherichia coli isolated from environmental, human and food samples. Int. J. Environ. Res. Public Health 2017, 14, 676. [Google Scholar] [CrossRef] [PubMed]
- van Hoek, A.H.; Veenman, C.; van Overbeek, W.M.; Lynch, G.; de Roda Husman, A.M.; Blaak, H. Prevalence and characterization of ESBL-and AmpC-producing Enterobacteriaceae on retail vegetables. Int. J. Food Microbiol. 2015, 204, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Bliziotis, I.A. Pandrug-resistant Gram-negative bacteria: The dawn of the post-antibiotic era? Int. J. Antimicrob. Agents 2007, 29, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Darphorn, T.S.; Bel, K.; Koenders-van Sint Anneland, B.B.; Brul, S.; Ter Kuile, B.H. Antibiotic resistance plasmid composition and architecture in Escherichia coli isolates from meat. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Babiker, W.; Ahmed, A.; Babiker, T.; Ibrahim, E.; Almugadam, B. Prevalence and Causes of Neonatal Sepsis in Soba University Hospital, Sudan. Med. Microbiol. Rep. 2018, 3, 11–13. [Google Scholar]
- Chowdhury, P.R.; McKinnon, J.; Liu, M.; Djordjevic, S.P. Multidrug resistant uropathogenic Escherichia coli ST405 with a novel, composite IS26 transposon in a unique chromosomal location. Front. Microbiol. 2019, 9, 3212. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, N.E.; Altayb, H.N.; Gurashi, R.M. Detection of Carbapenem-Resistant Genes in Escherichia coli Isolated from Drinking Water in Khartoum, Sudan. J. Environ. Public Health 2020, 2020, 2571293. [Google Scholar] [CrossRef]
- Moglad, E.; Adam, O.J.; Alnosh, M.; Altayb, H. Detection of virulence genes of diarrheagenic Escherichia coli strains from drinking water in Khartoum State. J. Water Health 2020, jwh2020097. [Google Scholar] [CrossRef]
- Stercz, B.; Farkas, F.B.; Tóth, Á.; Gajdács, M.; Domokos, J.; Horváth, V.; Ostorházi, E.; Makra, N.; Kocsis, B.; Juhász, J. The influence of antibiotics on transitory resistome during gut colonization with CTX-M-15 and OXA-162 producing Klebsiella pneumoniae ST15. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Alonso, C.; Zarazaga, M.; Ben Sallem, R.; Jouini, A.; Ben Slama, K.; Torres, C. Antibiotic resistance in Escherichia coli in husbandry animals: The African perspective. Lett. Appl. Microbiol. 2017, 64, 318–334. [Google Scholar] [CrossRef]
- Pitout, J.D.; Peirano, G.; Kock, M.M.; Strydom, K.-A.; Matsumura, Y. The global ascendency of OXA-48-type carbapenemases. Clin. Microbiol. Rev. 2019, 33, e00102–e00119. [Google Scholar] [CrossRef]
- Mata, C.; Miró, E.; Mirelis, B.; Garcillán-Barcia, M.P.; De La Cruz, F.; Coll, P.; Navarro, F. In vivo transmission of a plasmid coharbouring bla DHA-1 and qnr B genes between Escherichia coli and Serratia marcescens. FEMS Microbiol. Lett. 2010, 308, 24–28. [Google Scholar] [CrossRef]
- Guo, Q.; Spychala, C.N.; McElheny, C.L.; Doi, Y. Comparative analysis of an IncR plasmid carrying armA, bla DHA-1 and qnrB4 from Klebsiella pneumoniae ST37 isolates. J. Antimicrob. Chemother. 2016, 71, 882–886. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, K.; Xiao, S.; Xie, L.; Gu, F.; Li, X.; Ni, Y.; Sun, J.; Han, L. A Multidrug Resistance Plasmid pIMP26, Carrying bla IMP-26, fosA5, bla DHA-1, and qnrB 4 in Enterobacter cloacae. Sci. Rep. 2019, 9, 1–7. [Google Scholar]
- Takayama, Y.; Sekizuka, T.; Matsui, H.; Adachi, Y.; Eda, R.; Nihonyanagi, S.; Wada, T.; Matsui, M.; Suzuki, S.; Takaso, M. Characterization of the IncFII-IncFIB (pB171) plasmid carrying blaNDM-5 in Escherichia coli ST405 clinical isolate in Japan. Infect. Drug Resist. 2020, 13, 561. [Google Scholar] [CrossRef]
- Maurya, A.P.; Dhar, D.; Basumatary, M.K.; Paul, D.; Ingti, B.; Choudhury, D.; Talukdar, A.D.; Chakravarty, A.; Mishra, S.; Bhattacharjee, A. Expansion of highly stable bla OXA-10 β-lactamase family within diverse host range among nosocomial isolates of Gram-negative bacilli within a tertiary referral hospital of Northeast India. BMC Res. Notes 2017, 10, 1–6. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, X.; Guo, Q.; Wang, P.; Wang, W.; Wang, M. Characterization of fosA5, a new plasmid-mediated fosfomycin resistance gene in E scherichia coli. Lett. Appl. Microbiol. 2015, 60, 259–264. [Google Scholar] [CrossRef]
- Wang, H.; Min, C.; Li, J.; Yu, T.; Hu, Y.; Dou, Q.; Zou, M. Characterization of fosfomycin resistance and molecular epidemiology among carbapenem-resistant Klebsiella pneumoniae strains from two tertiary hospitals in China. BMC Microbiol. 2021, 21, 1–8. [Google Scholar] [CrossRef]
- Sorlozano-Puerto, A.; Lopez-Machado, I.; Albertuz-Crespo, M.; Martinez-Gonzalez, L.J.; Gutierrez-Fernandez, J. Characterization of fosfomycin and nitrofurantoin resistance mechanisms in Escherichia coli isolated in clinical urine samples. Antibiotics 2020, 9, 534. [Google Scholar] [CrossRef]
- Botelho, J.; Roberts, A.P.; León-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the move: It’s good to be on ICEs. Mob. DNA 2018, 9, 1–11. [Google Scholar] [CrossRef]
- He, J.; Li, C.; Cui, P.; Wang, H. Detection of Tn7-Like Transposons and Antibiotic Resistance in Enterobacterales from Animals Used for Food Production With Identification of Three Novel Transposons Tn6813, Tn6814, and Tn6765. Front. Microbiol. 2020, 11, 2049. [Google Scholar] [CrossRef]
- Biswas, B.; Azad, M.A.K.; Absar, N.; Islam, S.; Amin, S. Isolation and Identification of Pathogenic Bacteria from Fresh Fruits and Vegetables in Chittagong, Bangladesh. J. Microbiol. Res. 2020, 10, 55–58. [Google Scholar]
- Wauters, G.; Vaneechoutte, M. Approaches to the identification of aerobic Gram-negative bacteria. In Manual of Clinical Microbiology; Wiley: Hoboken, NJ, USA, 2015; pp. 613–634. [Google Scholar]
- Wayne, P. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Inform. Suppl. 2020, 31, 100–121. [Google Scholar]
- Sabeel, S.; Salih, M.A.; Ali, M.; El-Zaki, S.-E.; Abuzeid, N.; Elgadi, Z.A.M.; Altayb, H.N.; Elegail, A.; Ibrahim, N.Y.; Elamin, B.K. Phenotypic and genotypic analysis of multidrug-resistant Mycobacterium tuberculosis isolates from Sudanese patients. Tuberc. Res. Treat. 2017, 2017, 8340746. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Johansson, M.H.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, S.; Larsen, M.V.; Aarestrup, F.M.; Lund, O. PathogenFinder-distinguishing friend from foe using bacterial whole genome sequence data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Source | Specimen Type | PubMLST | ST | Accession No. |
|---|---|---|---|---|---|
| 1EP | Clinical sample | Wound swab | E. coli | 120 | JAJGXP000000000 |
| 5EP | Clinical sample | CSF | E. coli | 38 | JAJGXO000000000 |
| 8EP | Clinical sample | Urine | E. coli | 405 | JAJHQC000000000 |
| 10EP | Clinical sample | Wound swab | E. coli | 773 | JAJGXN000000000 |
| 11EP | Clinical sample | Wound swab | E. coli | 648 | JAJGXM000000000 |
| 15EP | Clinical sample | Urine | E. coli | 73 | JAJGXK000000000 |
| 20EP | Clinical sample | Urine | E. coli | 405 | JAJGXJ000000000 |
| 24EP | Clinical sample | Acetic fluid | E. coli | 405 | JAJGXI000000000 |
| 25EP | Clinical sample | Urine | E. coli | 405 | JAJGXH000000000 |
| 26EP | Clinical sample | Wound Swab | E. coli | 340 | JAJHGQ000000000 |
| 27EP | Clinical sample | Urine | E. coli | 656 | JAJGXG000000000 |
| 28EP | Clinical sample | Urine | E. coli | 410 | JAJGXF000000000 |
| 30EP | Clinical sample | Urine | E. coli | 405 | JAJGXE000000000 |
| 1EE | Hands | Swab | E. coli | 2522 | JAJGXX000000000 |
| 4EE | Hands | Swab | E. coli | 58 | JAJGXW000000000 |
| 6EE | Water | Water sample | E. coli | 4038 | JAJGXV000000000 |
| 7EE | Water | Water sample | E. coli | 448 | JAJGXU000000000 |
| 11EE | Vegetables | Swab | E. coli | 522 | JAJGXU000000000 |
| 12EP | Surface | Swab | E. coli | 2280 | JAJGXL000000000 |
| 13EE | Surface | Swab | E. coli | 1308 | JAJGXS000000000 |
| 14EE | Water | Water sample | E. coli | 1508 | JAJGXR000000000 |
| 16EE | Water | Water sample | E. coli | 1146 | JAJGXQ000000000 |
| 1KE | Vegetables | Swab | K. pneumoniae | 2365 | JAJHHI000000000 |
| 2KE | Surface | Swab | K. pneumoniae | 2177 | JAJHHH000000000 |
| 3KP | Clinical sample | Urine | K. pneumoniae | 76 | JAJHHG000000000 |
| 5KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHHF000000000 |
| 6KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHHE000000000 |
| 7KE | Surface | Swab | K. pneumoniae | 700 | JAJHHD000000000 |
| 7KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHHC000000000 |
| 8KE | Water | Water sample | K. pneumoniae | 1584 | JAJHNR000000000 |
| 11KE | Vegetables | Swab | K. pneumoniae | 1507 | JAJHHB000000000 |
| 12KE | Fruits | Swab | K. pneumoniae | 45 | JAJHHA000000000 |
| 12KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHGZ000000000 |
| 13KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHGY000000000 |
| 14KP | Clinical sample | Blood | K. pneumoniae | 45 | JAJHGX000000000 |
| 16KE | Surface | Swab | K. pneumoniae | 22,233 * | JAJHNQ000000000 |
| 17KE | Vegetables | Swab | K. pneumoniae | 22,234 * | JAJHGW000000000 |
| 19KE | Vegetables | Swab | K. pneumoniae | 45 | JAJHGV000000000 |
| 20KE | Vegetables | Swab | K. pneumoniae | 5624 | JAJHGU000000000 |
| 15KE | Surface | Swab | K. pneumoniae | 5808 | JAJHGT000000000 |
| 21KE | Surface | Swab | K. pneumoniae | 3335 | JAJHGS000000000 |
| 23KE | Water | Water samples | K. pneumoniae | 1504 * | JAJHGR000000000 |
| ID | Phenotypic AST | |||||||
|---|---|---|---|---|---|---|---|---|
| Beta-Lactam | Aminoglycosides and Fluoroquinolones | |||||||
| MR | IMP | CAZ | CTR | AMC | AK | GEN | CIP | |
| 1EP | S | S | R | R | R | S | S | S |
| 5EP | S | S | R | R | R | S | R | S |
| 8EP | S | S | R | R | R | S | S | R |
| 10EP | S | S | R | R | R | S | S | S |
| 11EP | S | S | R | R | R | R | R | S |
| 12EP | S | S | S | S | R | I | I | S |
| 15EP | S | S | S | S | R | S | S | S |
| 20EP | S | S | R | R | R | R | R | R |
| 24EP | S | S | S | S | R | S | S | S |
| 25EP | S | S | R | R | R | S | S | R |
| 26EP | S | S | R | R | R | S | S | R |
| 27EP | S | S | R | R | R | S | S | S |
| 28EP | S | S | R | R | R | S | S | S |
| 30EP | S | S | R | R | R | R | R | R |
| 1EE | S | S | S | S | S | S | S | S |
| 4EE | S | S | S | S | S | S | S | S |
| 6EE | S | S | S | S | S | S | S | S |
| 7EE | S | S | S | R | R | S | S | S |
| 11EE | S | S | S | S | R | S | R | S |
| 13EE | S | S | S | S | R | S | S | S |
| 14EE | S | S | S | S | R | I | I | S |
| 16EE | S | S | S | S | R | S | R | R |
| 3KP | S | S | S | S | R | S | S | S |
| 5KP | S | S | R | R | R | S | R | S |
| 6KP | S | S | R | R | R | S | R | R |
| 7KP | S | S | R | R | R | S | R | R |
| 12KP | S | S | R | R | R | S | S | R |
| 13KP | S | S | R | R | R | S | S | S |
| 14KP | S | S | R | R | R | S | S | S |
| 1KE | S | S | S | S | R | S | S | S |
| 2KE | S | S | S | S | R | S | S | S |
| 7KE | S | S | S | S | R | S | S | S |
| 8KE | S | S | S | S | R | S | S | S |
| 11KE | S | S | S | S | R | S | S | S |
| 12KE | S | S | S | S | R | S | S | S |
| 16KE | S | S | S | S | R | S | S | S |
| 15KE | S | S | S | S | R | S | S | S |
| 17KE | S | S | S | S | R | S | S | S |
| 19KE | S | S | R | R | R | S | S | S |
| 20KE | S | S | S | S | R | S | S | S |
| 21KE | S | S | S | S | R | S | S | S |
| 23KE | S | S | S | S | R | S | S | S |
| ID | Beta Lactam | Aminoglycosides and Fluoroquinolones | Others | ||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| blaCTX-M-15 | blaCTX-M-216 | blaOXA-1 | blaDHA-1 | blaTEM-35 | blaTEM-1 | blaCMY-141 | blaCMY-42 | blaNDM-5 | blaSHV-1 | blaSHV-12 | blaSHV-11 | blaSHV-26 | blaSHV-38 | blaSHV-71 | blaLEN16 | ompK37 | ompK36 | blaLEN16 | blaEC | qnrB4 | aac(6′)-Ib-cr | aac(3)-IIa | aadA5 | aph(3″)-Ib | aph(6)-Id | mdf(A) | Mdf | rmtB | qepA4 | qacE | qnrS1 | OqxB | OqxA | acrR | sul2 | Sul1 | dfrA | mph(A) | catB3 | catA1 | tet(A) | tet(B) | tet(39) | sitABCD | fosA | erm(C) | |
| 1EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 5EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 8EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 10EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 11EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 12EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 15EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 20EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 24EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 25EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 26EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 27EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 28EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 30EP | |||||||||||||||||||||||||||||||||||||||||||||||
| 1EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 4EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 6EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 7EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 11EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 13EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 14EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 16EE | |||||||||||||||||||||||||||||||||||||||||||||||
| 1KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 2KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 3KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 5KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 6KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 7KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 7KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 8KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 11KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 12KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 12KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 13KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 14KP | |||||||||||||||||||||||||||||||||||||||||||||||
| 15KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 16KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 17KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 19KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 20KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 21KE | |||||||||||||||||||||||||||||||||||||||||||||||
| 23KE | |||||||||||||||||||||||||||||||||||||||||||||||
| ID | IncFIA | IncFIB | IncFII | IncI1-I(Alpha) | IncY | IncR | IncFIB(H89-PhagePlasmid) | IncFIB(pB171) | IncFIB(pKPHS1) | IncFII(pHN7A8) | IncFIC(FII) | ncFII(pRSB107) | IncFIB(AP001918) | Col156 | Col(BS512) | Col440I | IncI(Gamma) | IncFII(pAMA1167-NDM-5) | IncFIB(pNDM-Mar) | IncHI1B(pNDM-MAR) | IncFII(pCoo) | IncFIA(HI1) | p0111 | IncFII(29) | IncFIB(K)(pCAV1099-114) | FIA(pBK30683) | IncFIB(K) | IncFII(K) | IncFII(pKP91) | ColpVC | pKP1433 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1EP | |||||||||||||||||||||||||||||||
| 5EP | |||||||||||||||||||||||||||||||
| 8EP | |||||||||||||||||||||||||||||||
| 10EP | |||||||||||||||||||||||||||||||
| 11EP | |||||||||||||||||||||||||||||||
| 12EP | |||||||||||||||||||||||||||||||
| 15EP | |||||||||||||||||||||||||||||||
| 20EP | |||||||||||||||||||||||||||||||
| 24EP | |||||||||||||||||||||||||||||||
| 25EP | |||||||||||||||||||||||||||||||
| 26EP | |||||||||||||||||||||||||||||||
| 27EP | |||||||||||||||||||||||||||||||
| 28EP | |||||||||||||||||||||||||||||||
| 30EP | |||||||||||||||||||||||||||||||
| 1EE | |||||||||||||||||||||||||||||||
| 4EE | |||||||||||||||||||||||||||||||
| 6EE | |||||||||||||||||||||||||||||||
| 7EE | |||||||||||||||||||||||||||||||
| 11EE | |||||||||||||||||||||||||||||||
| 13EE | |||||||||||||||||||||||||||||||
| 14EE | |||||||||||||||||||||||||||||||
| 16EE | |||||||||||||||||||||||||||||||
| 1KE | |||||||||||||||||||||||||||||||
| 2KE | |||||||||||||||||||||||||||||||
| 3KP | |||||||||||||||||||||||||||||||
| 5KP | |||||||||||||||||||||||||||||||
| 6KP | |||||||||||||||||||||||||||||||
| 7KE | |||||||||||||||||||||||||||||||
| 7KP | |||||||||||||||||||||||||||||||
| 8KE | |||||||||||||||||||||||||||||||
| 11KE | |||||||||||||||||||||||||||||||
| 12KE | |||||||||||||||||||||||||||||||
| 12KP | |||||||||||||||||||||||||||||||
| 13KP | |||||||||||||||||||||||||||||||
| 14KP | |||||||||||||||||||||||||||||||
| 16KE | |||||||||||||||||||||||||||||||
| 17KE | |||||||||||||||||||||||||||||||
| 19KE | |||||||||||||||||||||||||||||||
| 20KE | |||||||||||||||||||||||||||||||
| 15KE | |||||||||||||||||||||||||||||||
| 21KE | |||||||||||||||||||||||||||||||
| 23KE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altayb, H.N.; Elbadawi, H.S.; Alzahrani, F.A.; Baothman, O.; Kazmi, I.; Nadeem, M.S.; Hosawi, S.; Chaieb, K. Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach. Pharmaceuticals 2022, 15, 1011. https://doi.org/10.3390/ph15081011
Altayb HN, Elbadawi HS, Alzahrani FA, Baothman O, Kazmi I, Nadeem MS, Hosawi S, Chaieb K. Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach. Pharmaceuticals. 2022; 15(8):1011. https://doi.org/10.3390/ph15081011
Chicago/Turabian StyleAltayb, Hisham N., Hana S. Elbadawi, Faisal A. Alzahrani, Othman Baothman, Imran Kazmi, Muhammad Shahid Nadeem, Salman Hosawi, and Kamel Chaieb. 2022. "Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach" Pharmaceuticals 15, no. 8: 1011. https://doi.org/10.3390/ph15081011
APA StyleAltayb, H. N., Elbadawi, H. S., Alzahrani, F. A., Baothman, O., Kazmi, I., Nadeem, M. S., Hosawi, S., & Chaieb, K. (2022). Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach. Pharmaceuticals, 15(8), 1011. https://doi.org/10.3390/ph15081011