
Bacteriophage-Derived Double-Stranded RNA Exerts Anti-SARS-CoV-2 Activity In Vitro and in Golden Syrian Hamsters In Vivo


 , , ,
, , ,
Abstract
:
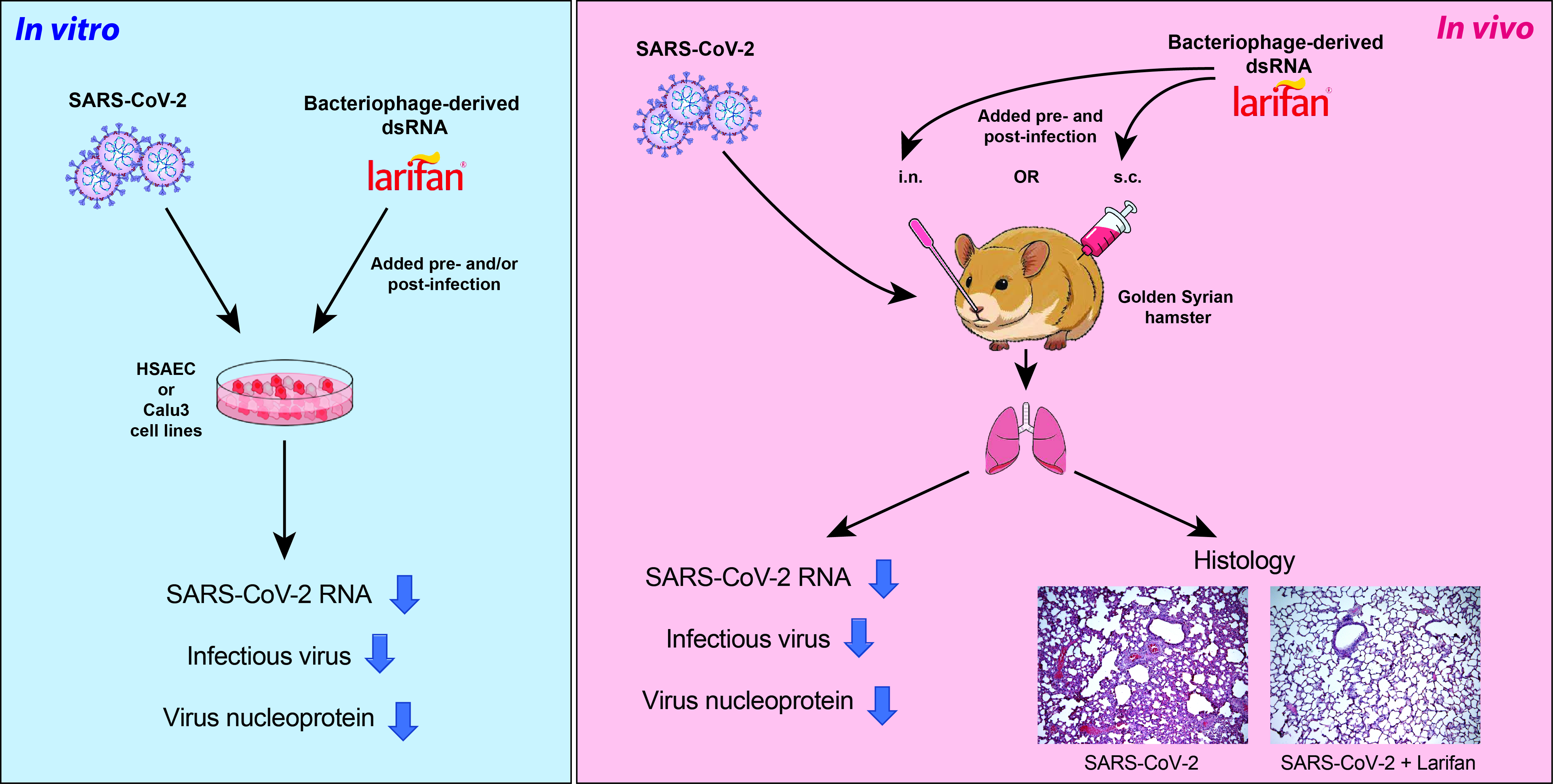{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Replication of SARS-CoV-2 in Calu3 and HSAEC Cells
2.2. Larifan Exserts an Antiviral Activity against SARS-CoV-2 In Vitro
2.3. Administration of Larifan Decreases SARS-CoV-2 Virus Numbers In Vivo
2.4. Administration of Larifan Reduces SARS-CoV-2 Antigen in Lungs and Improves Histological Lung Pathology
3. Discussion
4. Materials and Methods
4.1. SARS-CoV-2
4.2. Cell Lines
4.3. Compound
4.4. Cytotoxicity Assay
4.5. Infection of Cells with SARS-CoV-2 and Treatment with Larifan
4.6. SARS-CoV-2 Infection Model in Hamsters
4.7. RNA Isolation and Viral RNA Quantification by Digital Droplet PCR (ddPCR) Analysis
4.8. End-Point Virus Titration
4.9. Immunofluorescence Detection of SARS-CoV-2 in HSEAC and Hamster Lungs
4.10. Histology
4.11. Immunofluorescence Detection of SARS-CoV-2 in Hamster Lungs
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, C.; Ye, C.; Ruan, Z.; Liang, Y.; Li, Y.; Wu, J.; Luo, Z. Advances in the Development of Therapeutic Strategies against COVID-19 and Perspectives in the Drug Design for Emerging SARS-CoV-2 Variants. Comput. Struct. Biotechnol. J. 2022, 20, 824–837. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, I.; Sehgal, A.; Singh, S.; Sharma, N.; Anwer, M.K.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Bhatia, S.; et al. There Is Nothing Exempt from the Peril of Mutation—The Omicron Spike. Biomed. Pharmacother. 2022, 148, 112756. [Google Scholar] [CrossRef] [PubMed]
- Grein, J.; Ohmagari, N.; Shin, D.; Diaz, G.; Asperges, E.; Castagna, A.; Feldt, T.; Green, G.; Green, M.L.; Lescure, F.-X.; et al. Compassionate Use of Remdesivir for Patients with Severe COVID-19. N. Engl. J. Med. 2020, 382, 2327–2336. [Google Scholar] [CrossRef]
- Service, R.F. Bad News for Paxlovid? Resistance May Be Coming. Science 2022, 377, 138–139. [Google Scholar] [CrossRef]
- Sokolova, T.M.; Antonov, A.I.; Makhov, A.M. The dynamics of 2′,5′-oligoadenylate synthetase activity in the nuclear and cytoplasmic fractions of human fibroblasts treated by double-stranded RNAs, by their complexes with DEAE-dextran and by type-I recombinant interferons. The ratio of double-stran. Antibiot. i Khimioterapiia = Antibiot. Chemoterapy [Sic] 1991, 36, 31–35. [Google Scholar]
- Sokolova, T.M.; Nosik, N.N.; Lopatina, O.A.; Krylov, V.F. Evaluation of 2,5-oligoadenylate synthetase and protein kinase activities in clinical studies of larifan and recombinant interferon alpha in volunteers. Antibiot. I Khimioterapiia = Antibiot. Chemoterapy [Sic] 1991, 36, 42–45. [Google Scholar]
- Gebauer, F.; Hentze, M.W. Molecular Mechanisms of Translational Control. Nat. Rev. Mol. Cell Biol. 2004, 5, 827–835. [Google Scholar] [CrossRef]
- Finkel, Y.; Gluck, A.; Nachshon, A.; Winkler, R.; Fisher, T.; Rozman, B.; Mizrahi, O.; Lubelsky, Y.; Zuckerman, B.; Slobodin, B.; et al. SARS-CoV-2 Uses a Multipronged Strategy to Impede Host Protein Synthesis. Nature 2021, 594, 240–245. [Google Scholar] [CrossRef]
- Alavi Darazam, I.; Shokouhi, S.; Pourhoseingholi, M.A.; Naghibi Irvani, S.S.; Mokhtari, M.; Shabani, M.; Amirdosara, M.; Torabinavid, P.; Golmohammadi, M.; Hashemi, S.; et al. Role of Interferon Therapy in Severe COVID-19: The COVIFERON Randomized Controlled Trial. Sci. Rep. 2021, 11, 8059. [Google Scholar] [CrossRef] [PubMed]
- Nakhlband, A.; Fakhari, A.; Azizi, H. Interferon-Beta Offers Promising Avenues to COVID-19 Treatment: A Systematic Review and Meta-Analysis of Clinical Trial Studies. Naunyn. Schmiedebergs. Arch. Pharmacol. 2021, 394, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Nakhlband, A.; Fakhari, A.; Azizi, H. Interferon-Alpha Position in Combating with COVID-19: A Systematic Review. J. Med. Virol. 2021, 93, 5277–5284. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Riva, L.; Pu, Y.; Martin-Sancho, L.; Kanamune, J.; Yamamoto, Y.; Sakai, K.; Gotoh, S.; Miorin, L.; De Jesus, P.D.; et al. MDA5 Governs the Innate Immune Response to SARS-CoV-2 in Lung Epithelial Cells. Cell Rep. 2021, 34, 108628. [Google Scholar] [CrossRef]
- Lowery, S.A.; Sariol, A.; Perlman, S. Innate Immune and Inflammatory Responses to SARS-CoV-2: Implications for COVID-19. Cell Host Microbe 2021, 29, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhuang, M.-W.; Han, L.; Zhang, J.; Nan, M.-L.; Zhan, P.; Kang, D.; Liu, X.; Gao, C.; Wang, P.-H. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Membrane (M) Protein Inhibits Type I and III Interferon Production by Targeting RIG-I/MDA-5 Signaling. Signal Transduct. Target. Ther. 2020, 5, 299. [Google Scholar] [CrossRef] [PubMed]
- Tamir, H.; Melamed, S.; Erez, N.; Politi, B.; Yahalom-Ronen, Y.; Achdout, H.; Lazar, S.; Gutman, H.; Avraham, R.; Weiss, S.; et al. Induction of Innate Immune Response by TLR3 Agonist Protects Mice against SARS-CoV-2 Infection. Viruses 2022, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Sia, S.F.; Yan, L.-M.; Chin, A.W.H.; Fung, K.; Choy, K.-T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and Transmission of SARS-CoV-2 in Golden Hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Mautner, L.; Hoyos, M.; Dangel, A.; Berger, C.; Ehrhardt, A.; Baiker, A. Replication Kinetics and Infectivity of SARS-CoV-2 Variants of Concern in Common Cell Culture Models. Virol. J. 2022, 19, 76. [Google Scholar] [CrossRef]
- Do, T.N.D.; Claes, S.; Schols, D.; Neyts, J.; Jochmans, D. SARS-CoV-2 Virion Infectivity and Cytokine Production in Primary Human Airway Epithelial Cells. Viruses 2022, 14, 951. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Yuan, S.; Kok, K.-H.; To, K.K.-W.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.-Y.; Poon, R.W.-S.; et al. A Familial Cluster of Pneumonia Associated with the 2019 Novel Coronavirus Indicating Person-to-Person Transmission: A Study of a Family Cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef]
- Iwasaki, A.; Pillai, P.S. Innate Immunity to Influenza Virus Infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makris, S.; Johnston, S. Recent Advances in Understanding Rhinovirus Immunity. F1000Research 2018, 7, 1537. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Li, X.; Mou, T.; Zhou, X.; Li, D.; Wang, L.; Zhang, Y.; Dong, X.; Zheng, H.; Guo, L.; et al. Distinct Infection Process of SARS-CoV-2 in Human Bronchial Epithelial Cell Lines. J. Med. Virol. 2020, 92, 2830–2838. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, T.M.; Fatkhutdinova, N.G.; Nosik, N.N. Effect of double-stranded RNA and interferon type I on reproduction of cytomegalovirus in human fibroblasts. Biull. Eksp. Biol. Med. 1991, 112, 80–83. [Google Scholar] [CrossRef]
- Nosik, N.N.; Nosik, D.N.; Kuznetsova, N.V. The inhibiting action of interferon inducers on the multiplication of the human immunodeficiency virus. Vopr. Virusol. 1992, 37, 92–94. [Google Scholar]
- Alimbarova, L.M.; Barinskiĭ, I.F. Efficacy of herpetic vaccine, larifan, myelopid and their combinations during herpesvirus infection in irradiated animals. Vopr. Virusol. 1996, 41, 270–274. [Google Scholar]
- Loginova, S.I.; Bogatikov, G.V.; Prikhod’ko, A.V.; Polozov, A.I. Study of the effectiveness of recombinant alpha2-interferon and its inducers in primates with orthopoxvirus infection. Vopr. Virusol. 1997, 42, 186–188. [Google Scholar]
- Loginova, S.I.; Efanova, T.N.; Koval’chuk, A.V.; Faldina, V.N.; Androshchuk, I.A.; Pistsov, M.N.; Borisevich, S.V.; Kopylova, N.K.; Pashchenko, I.I.; Khamitov, P.A.; et al. Effectiveness of virazol, realdiron and interferon inductors in experimental Omsk hemorrhagic fever. Vopr. Virusol. 2002, 47, 27–30. [Google Scholar]
- Ershov, F.I.; Barinskiĭ, I.F.; Podcherniaeva, R.I.; Gribencha, S.V.; Popova, O.M. Immunostimulating effect of interferon inducers. Vopr. Virusol. 1983, 28, 74–79. [Google Scholar]
- Gribencha, S.V.; Nosik, N.N.; Ershov, F.I.; Barinskiĭ, I.F. Prophylactic and therapeutic activity of an interferon inducer in mice infected intracerebrally with rabies street virus. Vopr. Virusol. 1983, 28, 71–74. [Google Scholar] [PubMed]
- Mamedova, S.A.; Gashimov, R.G.; Bikbulatov, R.M. The use of the ultrasonic spraying of interferon and larifan solutions for treating acute herpetic stomatitis in children. Stomatologiia 1991, 5, 68–70. [Google Scholar]
- Rogovskaya, S.I.; Zhdanov, A.V.; Loginova, N.S.; Faizullin, L.Z.; Prilepskaya, V.N.; Van’ko, L.V.; Sukhikh, G.T. Interferon System in Women with Genital Papillomavirus Infection Receiving Immunomodulatory Therapy. Bull. Exp. Biol. Med. 2002, 134, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Potekaev, N.S.; Nosik, N.N.; Samgin, M.A.; Kudratullaev, K.N.; Lavrukhina, L.A. Experience with the combined treatment of recurrent herpes using larifan and a herpetic vaccine. Klin. Med. 1992, 70, 67–70. [Google Scholar]
- Veinalde, R.; Petrovska, R.; Brūvere, R.; Feldmane, G.; Pjanova, D. Ex Vivo Cytokine Production in Peripheral Blood Mononuclear Cells after Their Stimulation with DsRNA of Natural Origin. Biotechnol. Appl. Biochem. 2014, 61, 65–73. [Google Scholar] [CrossRef]
- Pjanova, D.; Mandrika, L.; Petrovska, R.; Vaivode, K.; Donina, S. Comparison of the Effects of Bacteriophage-Derived DsRNA and Poly(I:C) on Ex Vivo Cultivated Peripheral Blood Mononuclear Cells. Immunol. Lett. 2019, 212, 114–119. [Google Scholar] [CrossRef]
- Lemaire, P.A.; Anderson, E.; Lary, J.; Cole, J.L. Mechanism of PKR Activation by DsRNA. J. Mol. Biol. 2008, 381, 351–360. [Google Scholar] [CrossRef]
- Liu, G.; Lee, J.-H.; Parker, Z.M.; Acharya, D.; Chiang, J.J.; van Gent, M.; Riedl, W.; Davis-Gardner, M.E.; Wies, E.; Chiang, C.; et al. ISG15-Dependent Activation of the Sensor MDA5 Is Antagonized by the SARS-CoV-2 Papain-like Protease to Evade Host Innate Immunity. Nat. Microbiol. 2021, 6, 467–478. [Google Scholar] [CrossRef]
- Beyer, D.K.; Forero, A. Mechanisms of Antiviral Immune Evasion of SARS-CoV-2. J. Mol. Biol. 2022, 434, 167265. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian Hamsters as a Small Animal Model for SARS-CoV-2 Infection and Countermeasure Development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]
- Arifin, W.N.; Zahiruddin, W.M. Sample Size Calculation in Animal Studies Using Resource Equation Approach. Malays. J. Med. Sci. 2017, 24, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.F.; Jacobs, S.; Langendries, L.; Seldeslachts, L.; Ter Horst, S.; Liesenborghs, L.; Hens, B.; Vergote, V.; Heylen, E.; Barthelemy, K.; et al. Favipiravir at High Doses Has Potent Antiviral Activity in SARS-CoV-2-Infected Hamsters, Whereas Hydroxychloroquine Lacks Activity. Proc. Natl. Acad. Sci. USA 2020, 117, 26955–26965. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaivode, K.; Verhovcova, I.; Skrastina, D.; Petrovska, R.; Kreismane, M.; Lapse, D.; Kalnina, Z.; Salmina, K.; Rubene, D.; Pjanova, D. Bacteriophage-Derived Double-Stranded RNA Exerts Anti-SARS-CoV-2 Activity In Vitro and in Golden Syrian Hamsters In Vivo. Pharmaceuticals 2022, 15, 1053. https://doi.org/10.3390/ph15091053
Vaivode K, Verhovcova I, Skrastina D, Petrovska R, Kreismane M, Lapse D, Kalnina Z, Salmina K, Rubene D, Pjanova D. Bacteriophage-Derived Double-Stranded RNA Exerts Anti-SARS-CoV-2 Activity In Vitro and in Golden Syrian Hamsters In Vivo. Pharmaceuticals. 2022; 15(9):1053. https://doi.org/10.3390/ph15091053
Chicago/Turabian StyleVaivode, Kristine, Irina Verhovcova, Dace Skrastina, Ramona Petrovska, Madara Kreismane, Daira Lapse, Zane Kalnina, Kristine Salmina, Diana Rubene, and Dace Pjanova. 2022. "Bacteriophage-Derived Double-Stranded RNA Exerts Anti-SARS-CoV-2 Activity In Vitro and in Golden Syrian Hamsters In Vivo" Pharmaceuticals 15, no. 9: 1053. https://doi.org/10.3390/ph15091053
APA StyleVaivode, K., Verhovcova, I., Skrastina, D., Petrovska, R., Kreismane, M., Lapse, D., Kalnina, Z., Salmina, K., Rubene, D., & Pjanova, D. (2022). Bacteriophage-Derived Double-Stranded RNA Exerts Anti-SARS-CoV-2 Activity In Vitro and in Golden Syrian Hamsters In Vivo. Pharmaceuticals, 15(9), 1053. https://doi.org/10.3390/ph15091053