
Extraction, Isolation, Characterization, and Biological Activity of Sulfated Polysaccharides Present in Ascidian Viscera Microcosmus exasperatus
 and
and
Abstract
:
1. Introduction
2. Results
2.1. Morphology of the Ascidian Microcosmus Exasperatus
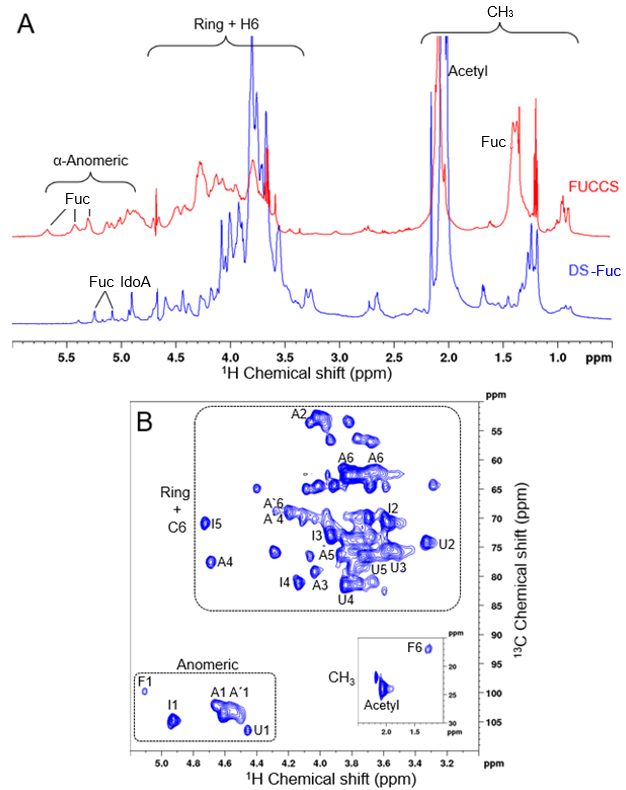
2.2. Microcosmus Exasperatus Viscera Presents a Dermatan Sulfate and a Fucosylated-Dermatan Sulfate
2.3. Microcosmus Exasperatus-Derived Dermatan Sulfate-Containing Fraction (SP2) Presents Anticoagulant Activity
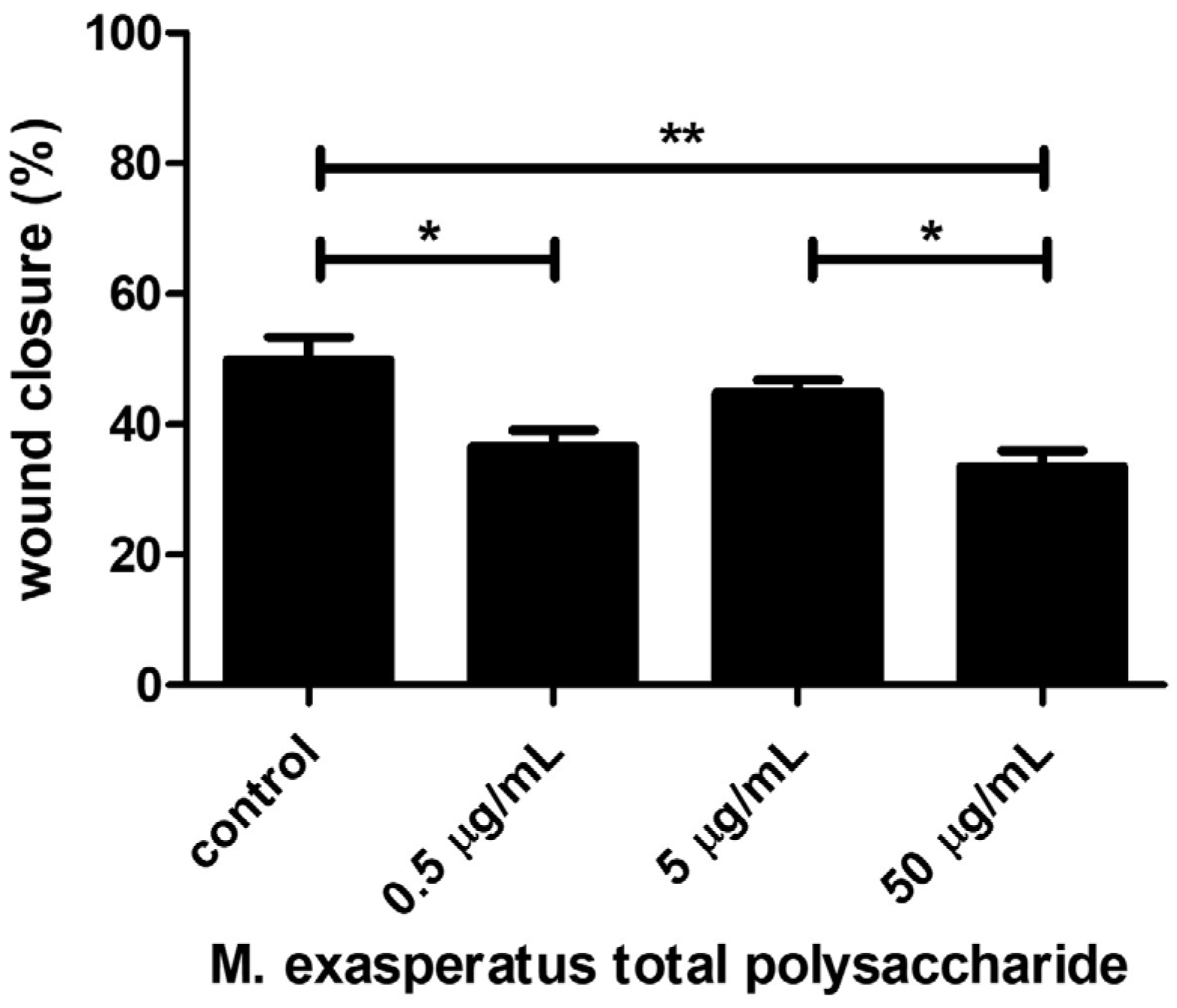
2.4. Microcosmus Exasperatus Total Polysaccharides (TP) Presents Antitumor Activity
3. Discussion
4. Materials and Methods
4.1. Specimens Collection and Dissection
4.2. GAGs Extraction from the Microcosmus Exasperatus Viscera
4.2.1. Delipidation and Depigmentation
4.2.2. Proteolytic Digestion
4.2.3. GAGs Precipitation
4.3. Sulfated Polysaccharides Electrophoretic Profile Analysis
4.4. Sulfated Polysaccharides Purification and Fractionation by Ion Exchange Liquid Chromatography
4.5. Lyases Specific Degradation and Nitrous Acid Deamination
4.6. Uronic Acid Analysis
4.7. Nuclear Magnetic Resonance (NMR) Analyses
4.8. Activated Partial Thromboplastin Time (APTT)
4.9. Cell Culture
4.10. Cell Viability Assays (MTT)
4.11. Clonogenic Assay
4.12. Wound Healing—Cell Migration—Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holland, L.Z. A Chordate with a Difference. Nature 2007, 447, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Jacobs, M.W.; Swalla, B.J. Coloniality Has Evolved Once in Stolidobranch Ascidians. Integr. Comp. Biol. 2006, 46, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, R146–R152. [Google Scholar] [CrossRef]
- Bettoni, R.; Hudson, C.; Williaume, G.; Sirour, C.; Yasuo, H.; de Buyl, S.; Dupont, G. Model of Neural Induction in the Ascidian Embryo. PLoS Comput. Biol. 2023, 19, e1010335. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, P.; Yi, C.-H. Biodiversity and Spatial Distribution of Ascidian Using Environmental DNA Metabarcoding. Mar. Environ. Res. 2023, 185, 105893. [Google Scholar] [CrossRef] [PubMed]
- Zemann, B.; Le, M.-L.V.; Sherlock, R.E.; Baum, D.; Katija, K.; Stach, T. Evolutionary Traces of Miniaturization in a Giant—Comparative Anatomy of Brain and Brain Nerves in Bathochordaeus stygius (Tunicata, Appendicularia). J. Morphol. 2023, 284, e21598. [Google Scholar] [CrossRef]
- Nishino, A.; Satou, Y.; Morisawa, M.; Satoh, N. Brachyury (T) Gene Expression and Notochord Development in Oikopleura longicauda (Appendicularia, Urochordata). Dev. Genes. Evol. 2001, 211, 219–231. [Google Scholar] [CrossRef]
- Matthysse, A.G.; Deschet, K.; Williams, M.; Marry, M.; White, A.R.; Smith, W.C. A Functional Cellulose Synthase from Ascidian Epidermis. Proc. Natl. Acad. Sci. USA 2004, 101, 986–991. [Google Scholar] [CrossRef]
- Kozlowski, E.O.; Lima, P.C.; Vicente, C.P.; Lotufo, T.; Bao, X.; Sugahara, K.; Pavão, M.S.G. Dermatan Sulfate in Tunicate Phylogeny: Order-Specific Sulfation Pattern and the Effect of [→4IdoA(2-Sulfate)β-1→3GalNAc(4-Sulfate)β-1→] Motifs in Dermatan Sulfate on Heparin Cofactor II Activity. BMC Biochem. 2011, 12, 29. [Google Scholar] [CrossRef]
- Yanagida, T.; Nakayama, K.; Sawada, T.; Honjo, M.; Murakami, S.; Iida, T.; Hirose, E.; Kitamura, S.-I. Innate Immunity in the Edible Ascidian Halocynthia roretzi Developing Soft Tunic Syndrome: Hemolymph Can Eliminate the Causative Flagellates and Discriminate Allogeneic Hemocytes. Fish. Shellfish. Immunol. 2022, 127, 659–665. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L.; Cima, F. Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri. Mar. Drugs 2023, 21, 74. [Google Scholar] [CrossRef] [PubMed]
- Satake, H. Kobayashi Award 2021: Neuropeptides, Receptors, and Follicle Development in the Ascidian, Ciona Intestinalis Type A: New Clues to the Evolution of Chordate Neuropeptidergic Systems from Biological Niches. Gen. Comp. Endocrinol. 2023, 337, 114262. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, S.; Bozzo, M.; Pennati, A.; Candiani, S.; Pennati, R. Serotonin Receptors and Their Involvement in Melanization of Sensory Cells in Ciona Intestinalis. Cells 2023, 12, 1150. [Google Scholar] [CrossRef] [PubMed]
- Nanglu, K.; Lerosey-Aubril, R.; Weaver, J.C.; Ortega-Hernández, J. A Mid-Cambrian Tunicate and the Deep Origin of the Ascidiacean Body Plan. Nat. Commun. 2023, 14, 3832. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, M.; Saito, Y.; Ogawa, M.; Takeuchi, A.; Jing, Z.; Naganuma, T.; Hirose, E. Morphological Studies on the Bathyal Ascidian, Megalodicopia hians Oka 1918 (Octacnemidae, Phlebobranchia), with Remarks on Feeding and Tunic Morphology. Zool. Sci. 2002, 19, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lee, C.; Rho, S.; Lee, Y. Reproductive Cycle and Spawning Rhythm of the Ascidian, Halocynthia hilgendorfi Ritteri. Korean J. Biol. Sci. 2004, 8, 33–40. [Google Scholar] [CrossRef]
- Hickman, C.P., Jr.; Cleveland, H.J.; Keen, S.L.; Eisenhour, D.J.; Larson, A.; I’Anson, H. Integrated Principles of Zoology; McGraw Hill: New York, NY, USA, 2023; ISBN 978-1-266-57724-6. [Google Scholar]
- Di Bella, M.A.; Carbone, M.C.; De Leo, G. Aspects of Cell Production in Mantle Tissue of Ciona intestinalis L. (Tunicata, Ascidiacea). Micron 2005, 36, 477–481. [Google Scholar] [CrossRef]
- Gasparini, F.; Manni, L.; Cima, F.; Zaniolo, G.; Burighel, P.; Caicci, F.; Franchi, N.; Schiavon, F.; Rigon, F.; Campagna, D.; et al. Sexual and Asexual Reproduction in the Colonial Ascidian Botryllus schlosseri. Genesis 2015, 53, 105–120. [Google Scholar] [CrossRef]
- Sawada, H.; Saito, T. Mechanisms of Sperm–Egg Interactions: What Ascidian Fertilization Research Has Taught Us. Cells 2022, 11, 2096. [Google Scholar] [CrossRef]
- Kuplik, Z.; Novak, L.; Shenkar, N. Proteomic Profiling of Ascidians as a Tool for Biomonitoring Marine Environments. PLoS ONE 2019, 14, e0215005. [Google Scholar] [CrossRef]
- Vered, G.; Kaplan, A.; Avisar, D.; Shenkar, N. Using Solitary Ascidians to Assess Microplastic and Phthalate Plasticizers Pollution among Marine Biota: A Case Study of the Eastern Mediterranean and Red Sea. Mar. Pollut. Bull. 2019, 138, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Nagar, L.R.; Shenkar, N. Temperature and Salinity Sensitivity of the Invasive Ascidian Microcosmus exasperatus Heller, 1878. Aquat. Invasions 2016, 11, 33–43. [Google Scholar] [CrossRef]
- Navon, G.; Kaplan, A.; Avisar, D.; Shenkar, N. Assessing Pharmaceutical Contamination along the Mediterranean and Red Sea Coasts of Israel: Ascidians (Chordata, Ascidiacea) as Bioindicators. Mar. Pollut. Bull. 2020, 160, 111510. [Google Scholar] [CrossRef]
- Anderson, G.; Shenkar, N. Potential Effects of Biodegradable Single-Use Items in the Sea: Polylactic Acid (PLA) and Solitary Ascidians. Environ. Pollut. 2021, 268, 115364. [Google Scholar] [CrossRef]
- Bereza, D.; Shenkar, N. Shipping Voyage Simulation Reveals Abiotic Barriers to Marine Bioinvasions. Sci. Total Environ. 2022, 837, 155741. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Espinosa, D.C.; Román, Y.; Colorado-Ríos, J.; de Santana-Filho, A.P.; Sassaki, G.L.; Cipriani, T.R.; Martínez, A.; Iacomini, M.; Pavão, M.S.G. Structural Analysis of a Sulfated Galactan from the Tunic of the Ascidian Microcosmus exasperatus and Its Inhibitory Effect of the Intrinsic Coagulation Pathway. Int. J. Biol. Macromol. 2017, 105, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Espinosa, D.C.; Motta, J.M.; de Teixeira, F.C.O.B.; Román, Y.; Colorado-Ríos, J.; Pavão, M.S.G.; Martínez, A. Restrepo-Espinosa, D.C.; Motta, J.M.; Teixeira, F.C.O. de B.; et al. Anticoagulant Properties of a High Molecular Weight Polysaccharide Fraction (1000RS) of the Ascidian Microcosmus exasperatus. Ars Pharm. (Internet) 2017, 58, 95–101. [Google Scholar] [CrossRef]
- Santos, J.C.; Mesquita, J.M.F.; Belmiro, C.L.R.; da Silveira, C.B.M.; Viskov, C.; Mourier, P.A.; Pavão, M.S.G. Isolation and Characterization of a Heparin with Low Antithrombin Activity from the Body of Styela plicata (Chordata-Tunicata). Distinct Effects on Venous and Arterial Models of Thrombosis. Thromb. Res. 2007, 121, 213–223. [Google Scholar] [CrossRef]
- Valcarcel, J.; Vázquez, J.A.; Varela, U.R.; Reis, R.L.; Novoa-Carballal, R. Isolation and Characterization of Polysaccharides from the Ascidian Styela clava. Polymers 2021, 14, 16. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Ozbek, S. Evolution of Glycosaminoglycans: Comparative Biochemical Study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef]
- Yasuoka, Y. Tissue-Specific Expression of Carbohydrate Sulfotransferases Drives Keratan Sulfate Biosynthesis in the Notochord and Otic Vesicles of Xenopus Embryos. Front. Cell Dev. Biol. 2023, 11, 957805. [Google Scholar] [CrossRef]
- Neri, T.A.N.; Palmos, G.N.; Park, S.Y.; Jung, T.S.; Choi, B.-D. Hair Growth-Promoting Activities of Glycosaminoglycans Extracted from the Tunics of Ascidian (Halocynthia roretzi). Polymers 2022, 14, 1096. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-D.; Choi, Y.J. Nutraceutical Functionalities of Polysaccharides from Marine Invertebrates. Adv. Food Nutr. Res. 2012, 65, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Pavão, M.S.G. Glycosaminoglycans Analogs from Marine Invertebrates: Structure, Biological Effects, and Potential as New Therapeutics. Front. Cell Infect. Microbiol. 2014, 4, 123. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, G.F.; Palmero, C.Y.; de Souza-Menezes, J.; Araujo, A.K.; Guimarães, A.G.; de Barros, C.M. Dermatan Sulfate Obtained from the Phallusia Nigra Marine Organism Is Responsible for Antioxidant Activity and Neuroprotection in the Neuroblastoma-2A Cell Lineage. Int. J. Biol. Macromol. 2020, 164, 1099–1111. [Google Scholar] [CrossRef]
- Li, J.; Han, S.; Zhu, Y.; Dong, B. Halorotetin A: A Novel Terpenoid Compound Isolated from Ascidian Halocynthia Rotetzi Exhibits the Inhibition Activity on Tumor Cell Proliferation. Mar. Drugs 2023, 21, 51. [Google Scholar] [CrossRef]
- Medeiros, T.B.; Cosendey, P.; Gerin, D.R.; de Sousa, G.F.; Portal, T.M.; Monteiro-de-Barros, C. The Effect of the Sulfation Patterns of Dermatan and Chondroitin Sulfate from Vertebrates and Ascidians on Their Neuritogenic and Neuroprotective Properties. Int. J. Biol. Macromol. 2023, 247, 125830. [Google Scholar] [CrossRef]
- Hogwood, J.; Mulloy, B.; Lever, R.; Gray, E.; Page, C.P. Pharmacology of Heparin and Related Drugs: An Update. Pharmacol. Rev. 2023, 75, 328–379. [Google Scholar] [CrossRef]
- Vitiello, A.; Ferrara, F. Low Molecular Weight Heparin, Anti-Inflammatory/Immunoregulatory and Antiviral Effects, a Short Update. Cardiovasc. Drugs Ther. 2023, 37, 277–281. [Google Scholar] [CrossRef]
- Vasconcelos, A.A.; Pomin, V.H. The Sea as a Rich Source of Structurally Unique Glycosaminoglycans and Mimetics. Microorganisms 2017, 5, 51. [Google Scholar] [CrossRef]
- Mourão, P.A.; Pereira, M.S. Searching for Alternatives to Heparin: Sulfated Fucans from Marine Invertebrates. Trends Cardiovasc. Med. 1999, 9, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Stelling, M.P.; de Bento, A.A.; Caloba, P.; Vilanova, E.; Pavão, M.S.G. Methods for Isolation and Characterization of Sulfated Glycosaminoglycans from Marine Invertebrates. Methods Mol. Biol. 2019, 1952, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Pavão, M.S.; Aiello, K.R.; Werneck, C.C.; Silva, L.C.; Valente, A.P.; Mulloy, B.; Colwell, N.S.; Tollefsen, D.M.; Mourão, P.A. Highly Sulfated Dermatan Sulfates from Ascidians. Structure versus Anticoagulant Activity of These Glycosaminoglycans. J. Biol. Chem. 1998, 273, 27848–27857. [Google Scholar] [CrossRef] [PubMed]
- Lambert, G. Ecology and Natural History of the Protochordates. Can. J. Zool. 2005, 83, 34–50. [Google Scholar] [CrossRef]
- Davidson, B.; Swalla, B.J. A Molecular Analysis of Ascidian Metamorphosis Reveals Activation of an Innate Immune Response. Development 2002, 129, 4739–4751. [Google Scholar] [CrossRef]
- Schlosser, G.; Thieffry, D. Modularity in Development and Evolution. Bioessays 2000, 22, 1043–1045. [Google Scholar] [CrossRef]
- Tiozzo, S.; Brown, F.D.; De Tomaso, A.W. Regeneration and Stem Cells in Ascidians. In Stem Cells; Bosch, T.C.G., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 95–112. ISBN 978-1-4020-8273-3. [Google Scholar]
- Mayzel, B.; Haber, M.; Ilan, M. Chemical Defense against Fouling in the Solitary Ascidian Phallusia nigra. Biol. Bull. 2014, 227, 232–241. [Google Scholar] [CrossRef]
- Pomin, V.H. Holothurian Fucosylated Chondroitin Sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef]
- Bitter, T.; Muir, H.M. A Modified Uronic Acid Carbazole Reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Eggleton, C.A.; Barrowcliffe, T.W.; Merton, R.E.; Thomas, D.P. In Vitro and in Vivo Studies of the Anti-Xa Activity of Heparin. Thromb. Res. 1981, 24, 319–328. [Google Scholar] [CrossRef]
- Thomas, D.P.; Merton, R.E.; Barrowcliffe, T.W.; Thunberg, L.; Lindahl, U. Effects of Heparin Oligosaccharides with High Affinity for Antithrombin III in Experimental Venous Thrombosis. Thromb. Haemost. 1982, 47, 244–248. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Units | C-H1 | C-H2 | C-H3 | C-H4 | C-H5 | C-H6 |
|---|---|---|---|---|---|---|
| GalNAC-4S # | 4.58–103.6 | 4.02–54.0 | 4.00–78.5 | 4.76–79.2 | 3.83–77.4 | 3.77–63.8 |
| GalNAC-4S a | 4.67/4.55–103.2 | 4.01–53.0 | 4.03–79.4 | 4.70–77.9 | 3.87–76.3 | 3.82/3.66–62.6 |
| GalNAC-6S # | 4.58–103.6 | 4.02–53.4 | 3.98–81.7 | 4.20–70.3 | 3.97–75.1 | 4.24–70.1 |
| GalNAC-6S a | 4.67/4.55–103.2 | 3.98–53.0 | 3.98–81.7 | 4.19–69.1 | - | 4.19–69.1 |
| GlcA # | 4.46–106.5 | 3.37–75.2 | 3.57–76.2 | 3.77–83.8 | 3.66–79.5 | - |
| GlcA a | 4.46–106.6 | 3.34–74.3 | 3.57–75.3 | 3.82–82.0 | 3.72–76.5 | - |
| IdoA * | 4.90–102.8 | 3.53–71.6 | 3.95–73.2 | 4.10–82.3 | 4.72–72.4 | - |
| IdoA a | 4.92–104.0 | 3.57–71.1 | 3.94–72.9 | 4.13–81.1 | 4.73–71.2 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bento, A.d.A.; Maciel, M.C.; Bezerra, F.F.; Mourão, P.A.d.S.; Pavão, M.S.G.; Stelling, M.P. Extraction, Isolation, Characterization, and Biological Activity of Sulfated Polysaccharides Present in Ascidian Viscera Microcosmus exasperatus. Pharmaceuticals 2023, 16, 1401. https://doi.org/10.3390/ph16101401
Bento AdA, Maciel MC, Bezerra FF, Mourão PAdS, Pavão MSG, Stelling MP. Extraction, Isolation, Characterization, and Biological Activity of Sulfated Polysaccharides Present in Ascidian Viscera Microcosmus exasperatus. Pharmaceuticals. 2023; 16(10):1401. https://doi.org/10.3390/ph16101401
Chicago/Turabian StyleBento, Ananda de Araujo, Marianna Cardoso Maciel, Francisco Felipe Bezerra, Paulo Antônio de Souza Mourão, Mauro Sérgio Gonçalves Pavão, and Mariana Paranhos Stelling. 2023. "Extraction, Isolation, Characterization, and Biological Activity of Sulfated Polysaccharides Present in Ascidian Viscera Microcosmus exasperatus" Pharmaceuticals 16, no. 10: 1401. https://doi.org/10.3390/ph16101401
APA StyleBento, A. d. A., Maciel, M. C., Bezerra, F. F., Mourão, P. A. d. S., Pavão, M. S. G., & Stelling, M. P. (2023). Extraction, Isolation, Characterization, and Biological Activity of Sulfated Polysaccharides Present in Ascidian Viscera Microcosmus exasperatus. Pharmaceuticals, 16(10), 1401. https://doi.org/10.3390/ph16101401