


Ligand-Based Drug Design of Genipin Derivatives with Cytotoxic Activity against HeLa Cell Line: A Structural and Theoretical Study

 , , , , and
, , , , and 
Highlights
- The importance of the dipole moment for the cytotoxic activity of iridoids.
- The lower potency of Genipin in HeLa cell line.
- The lack of activity of Geniposide in HeLa cell line.
- The importance of the double bond in C7 and C8 for the cytotoxic activity of iridoids.
Abstract
:1. Introduction
2. Results and Discussion
2.1. In Vitro Cytotoxic Activity of Geniposide and Genipin
2.2. Cytotoxic Iridoids Assessed against the HeLa Cell Line
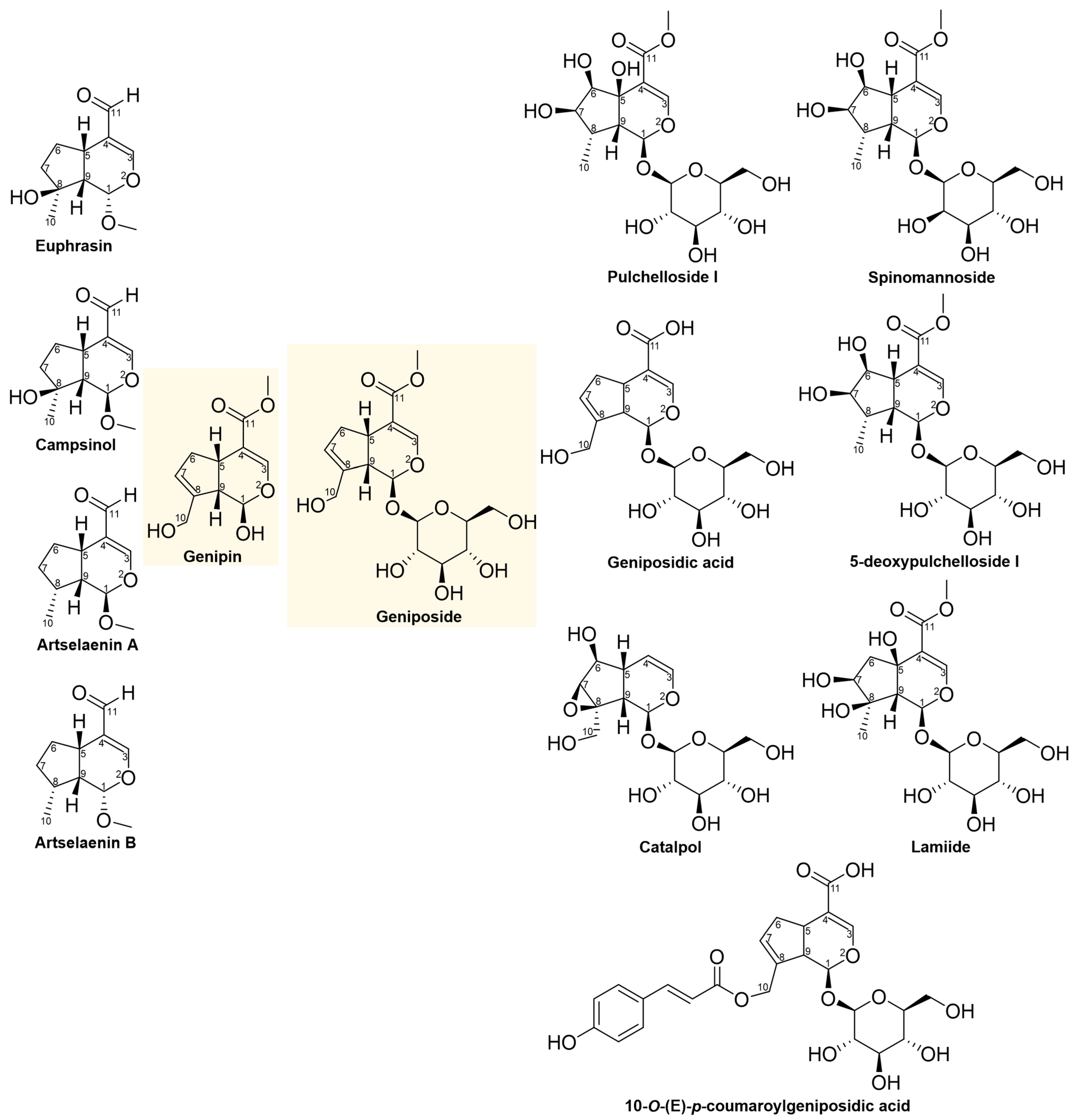
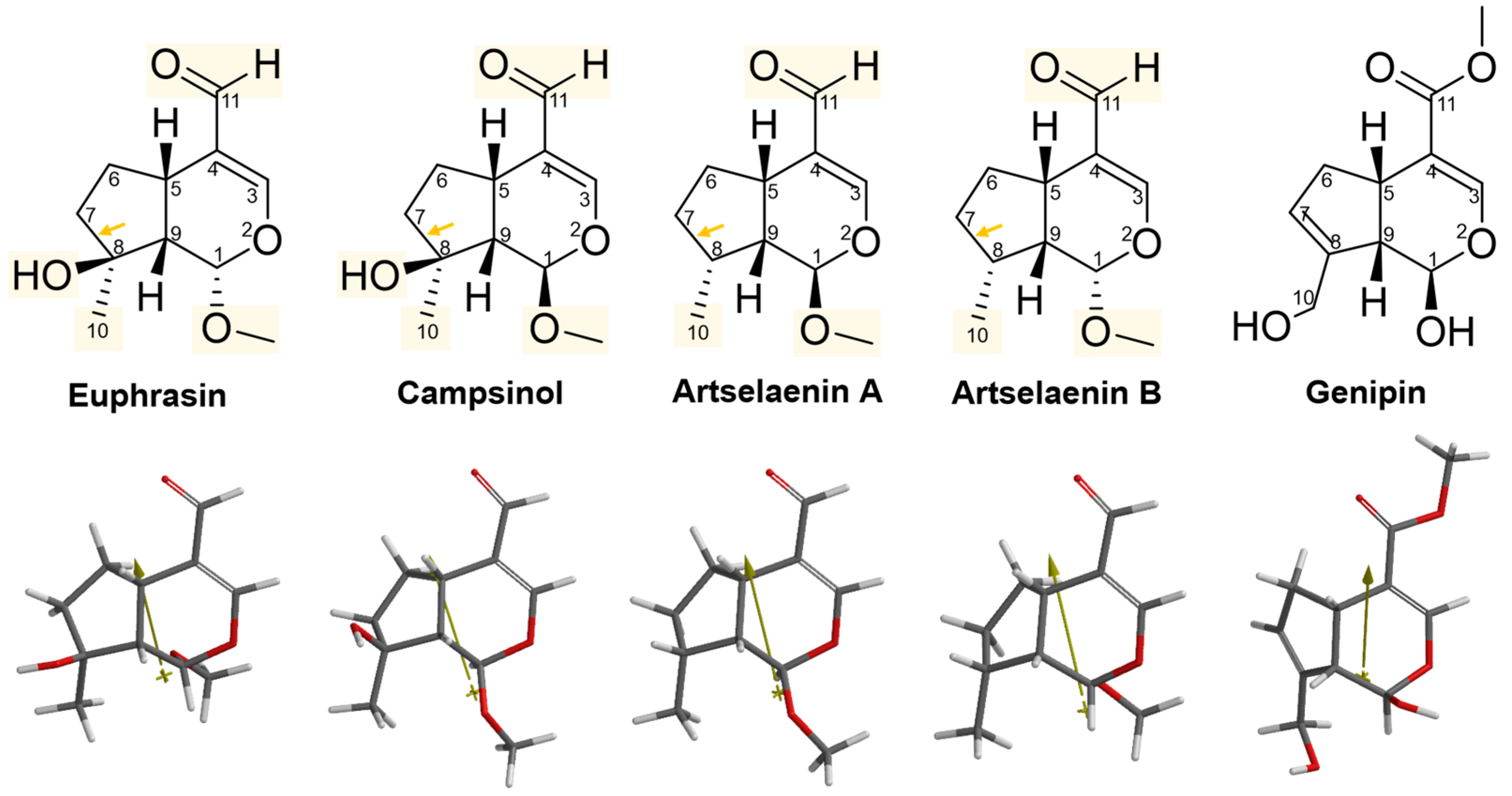
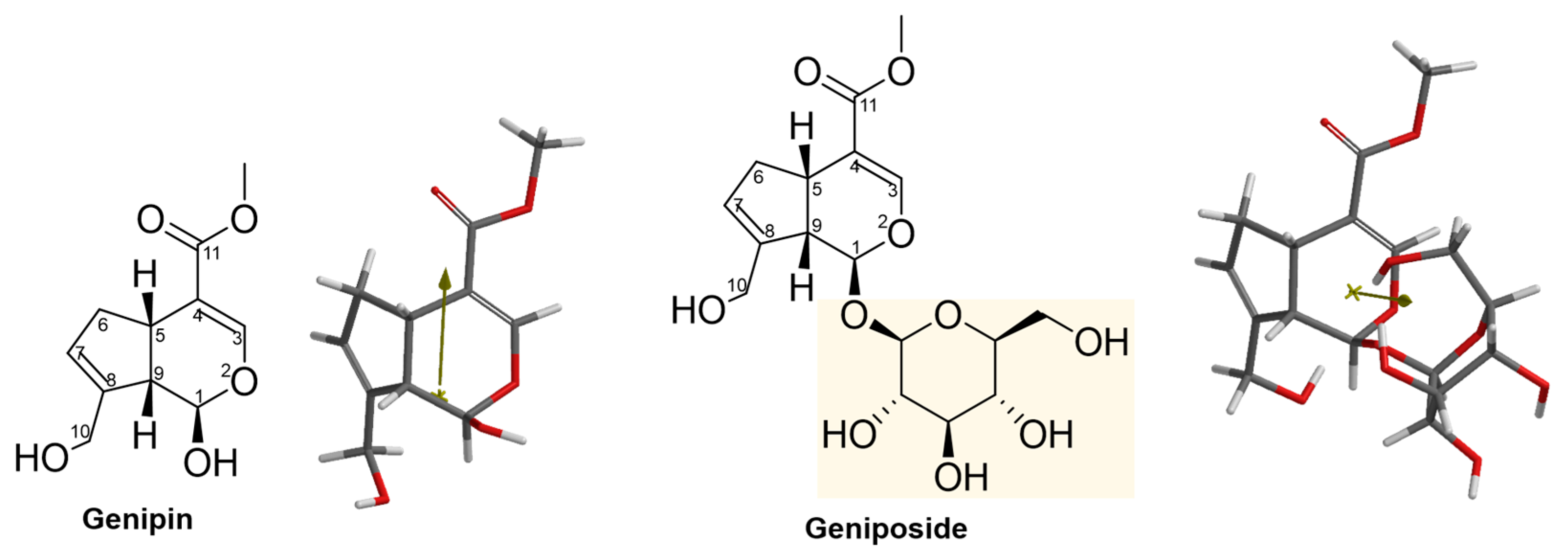
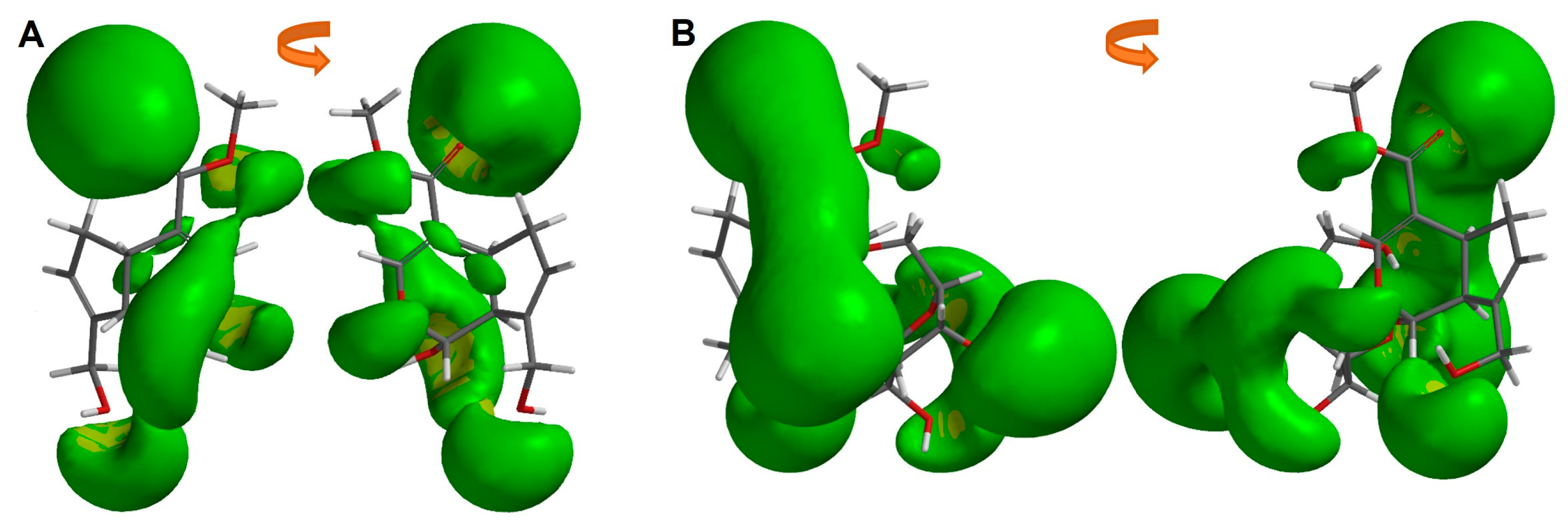
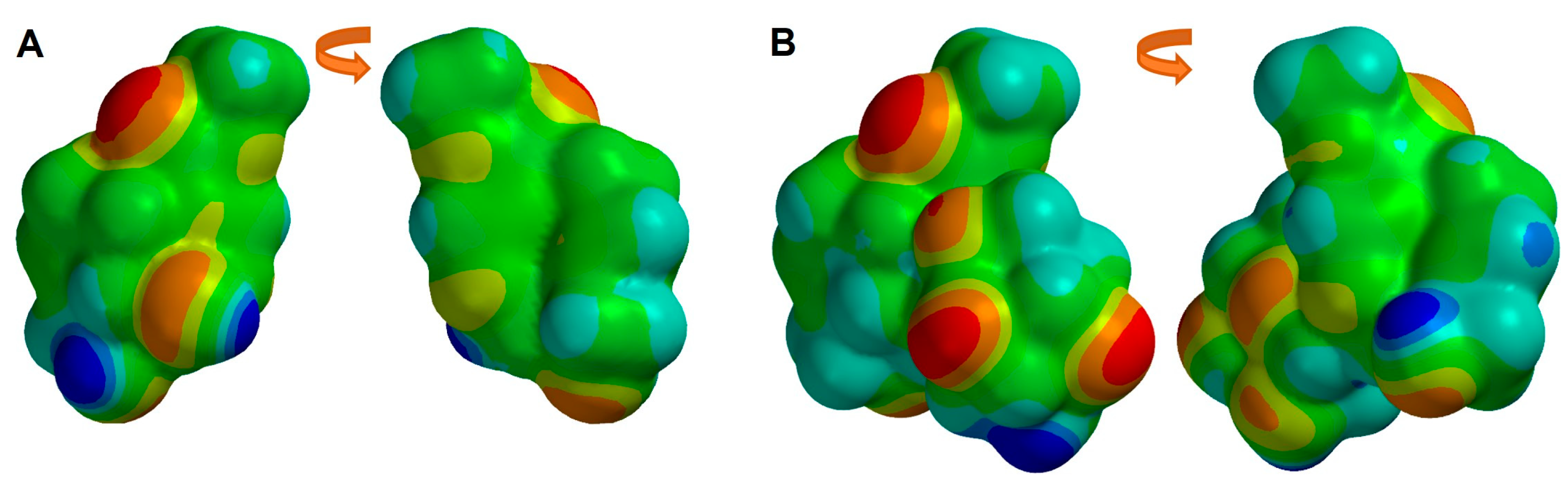
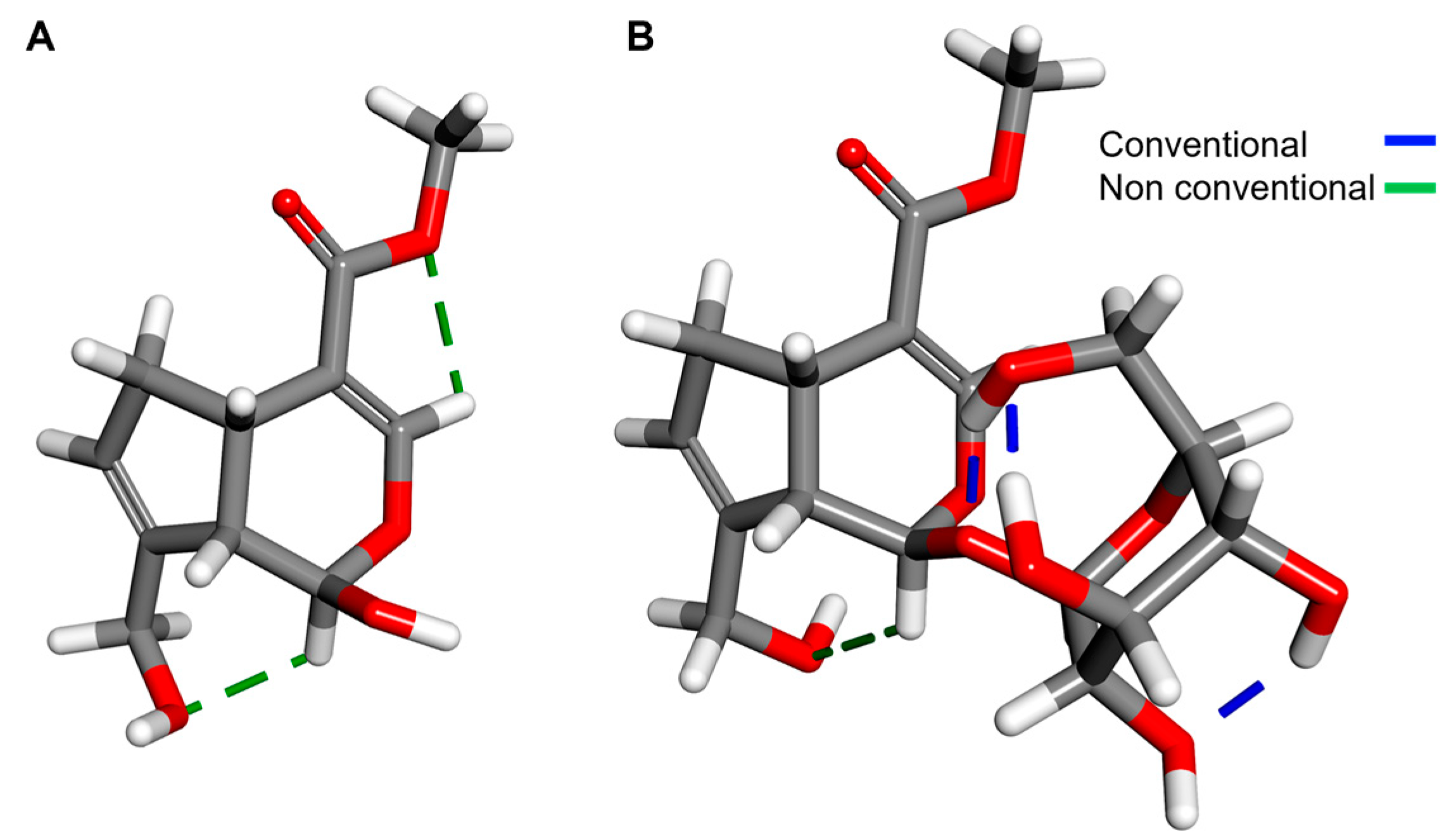
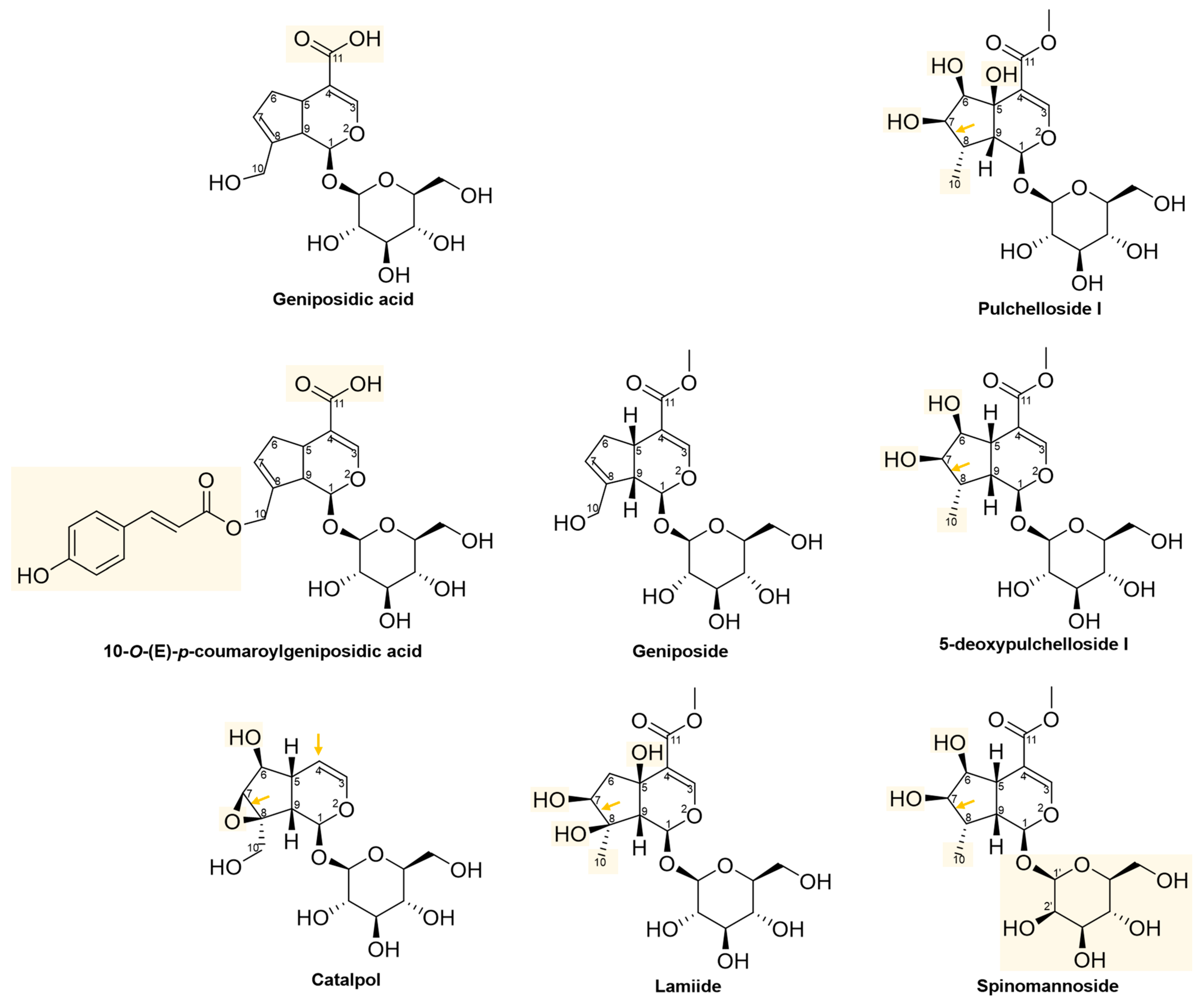
2.3. Structural and Molecular Analysis of Genipin and Other Simple Iridoids
2.4. Structural and Molecular Analysis of Genipin and Geniposide
2.5. Structural and Molecular Analysis of Geniposide and Other Iridoid Glycosides
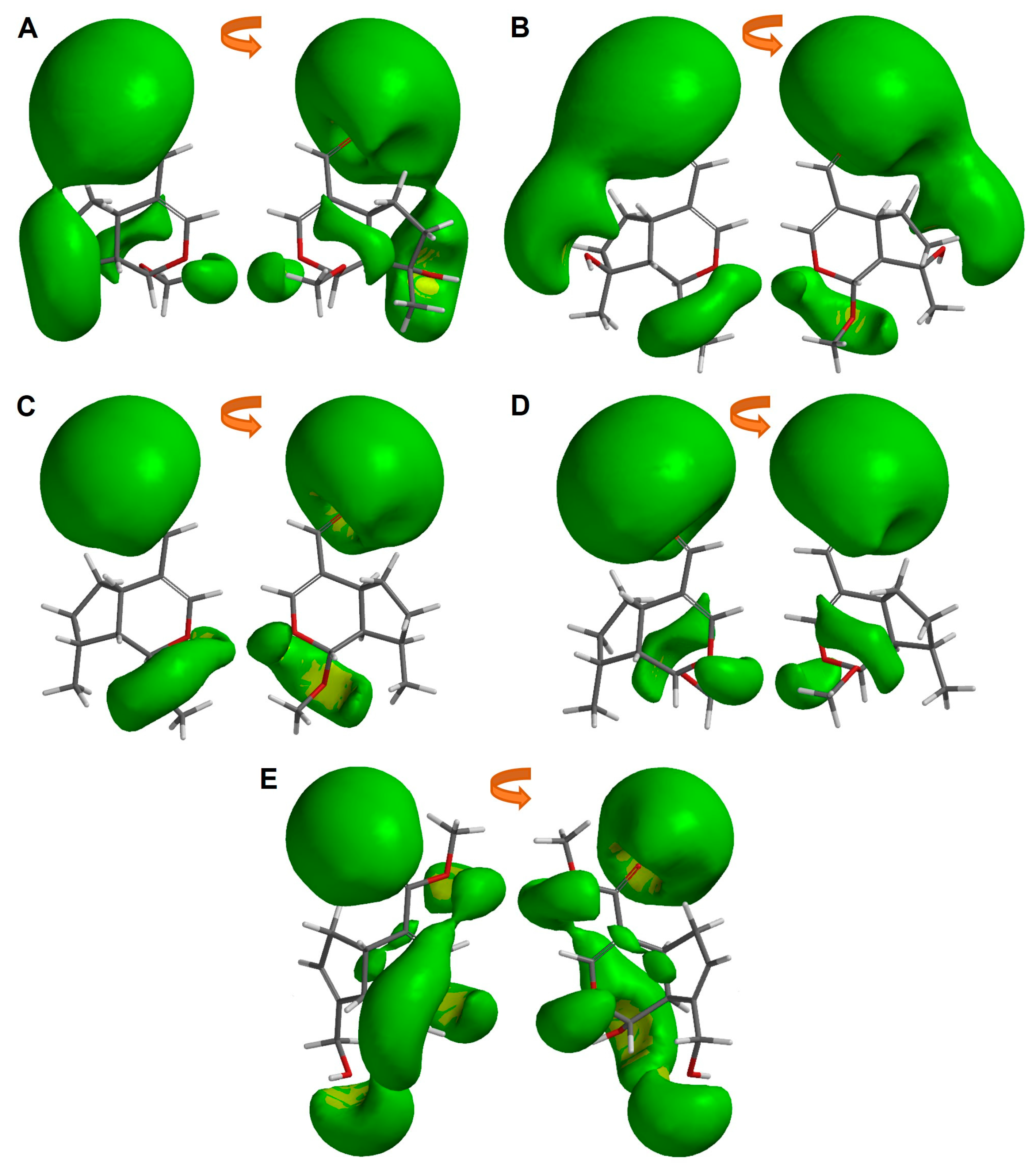
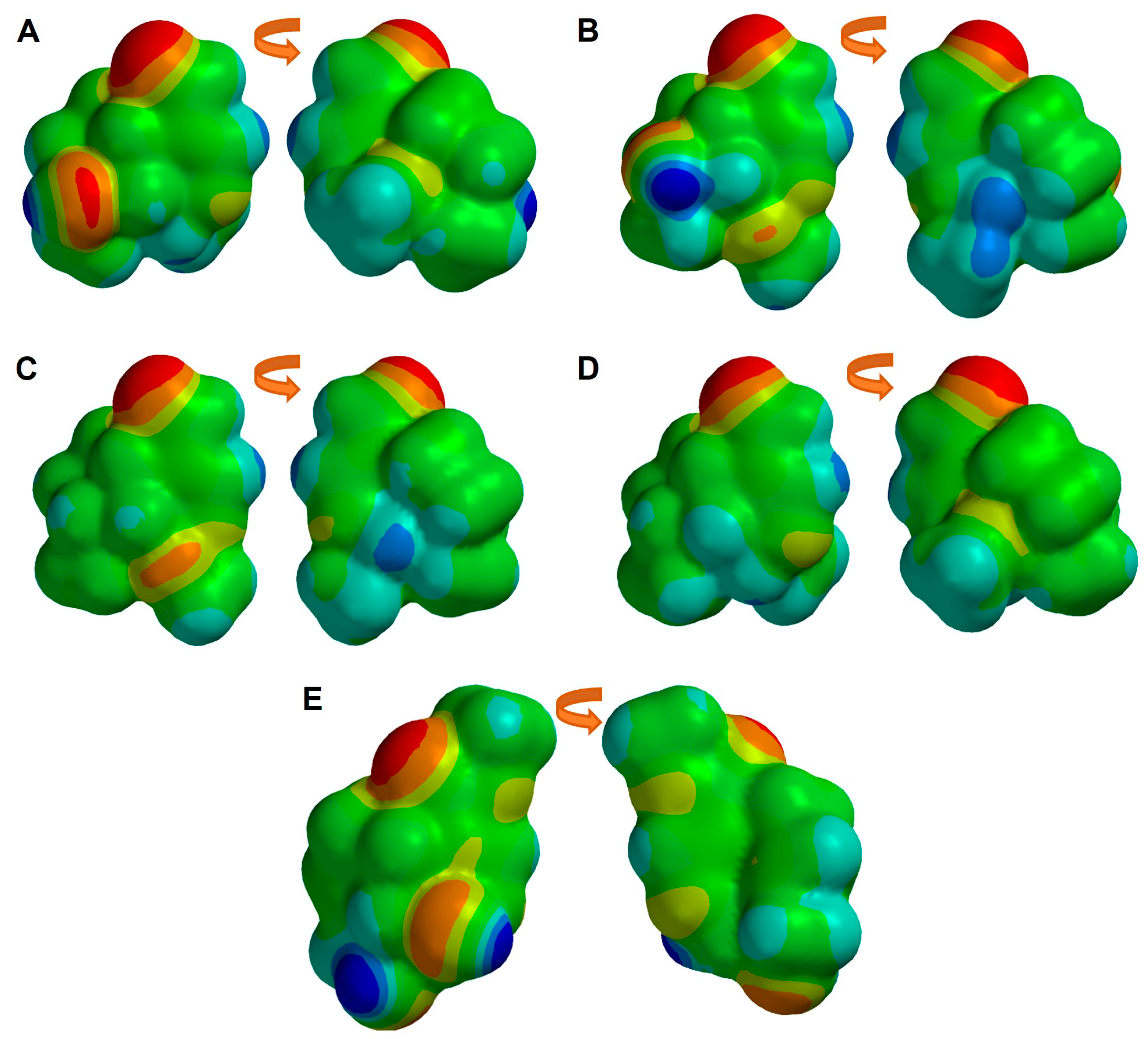
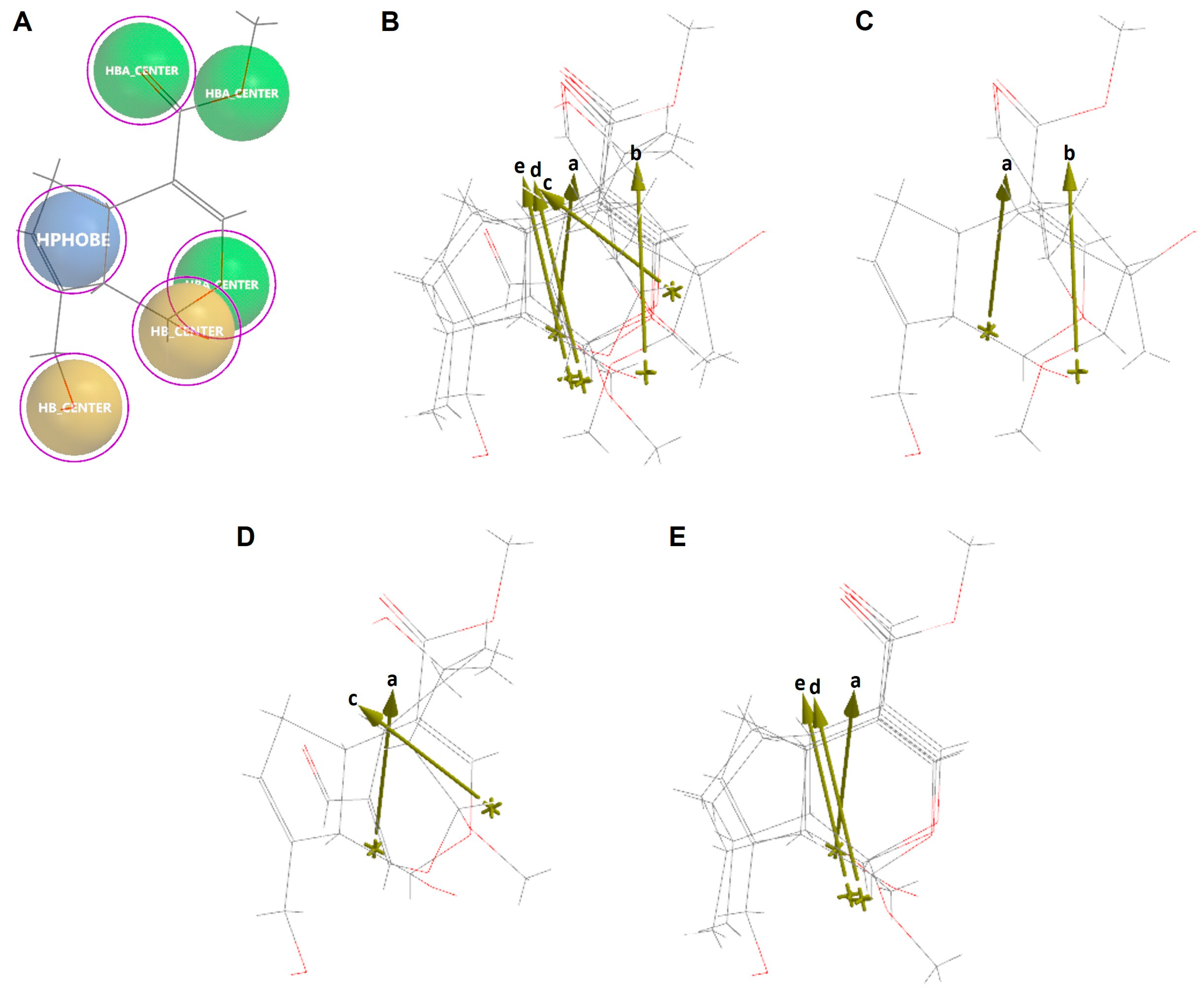
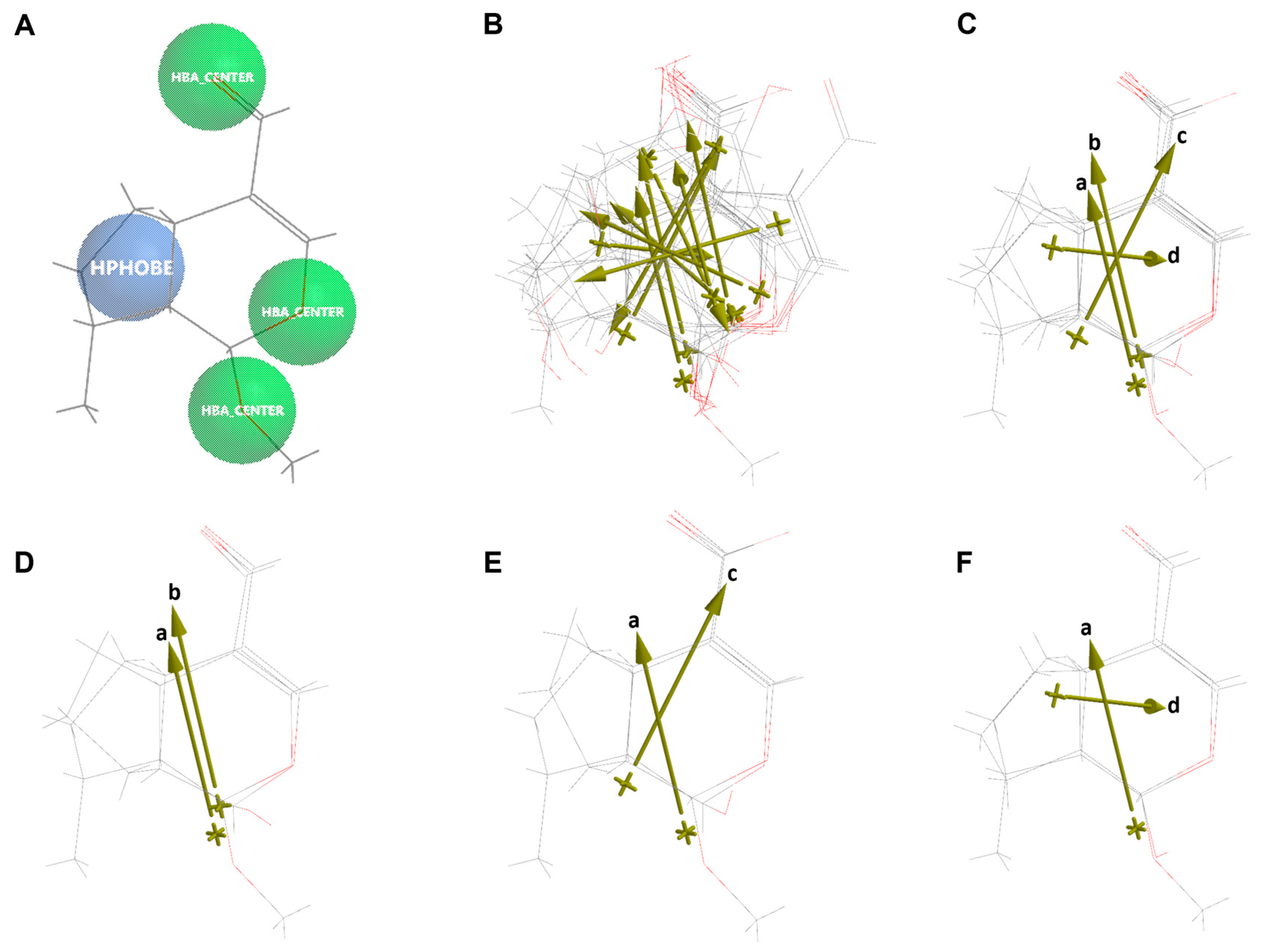
2.6. Molecular Similarity Analysis of Iridoids
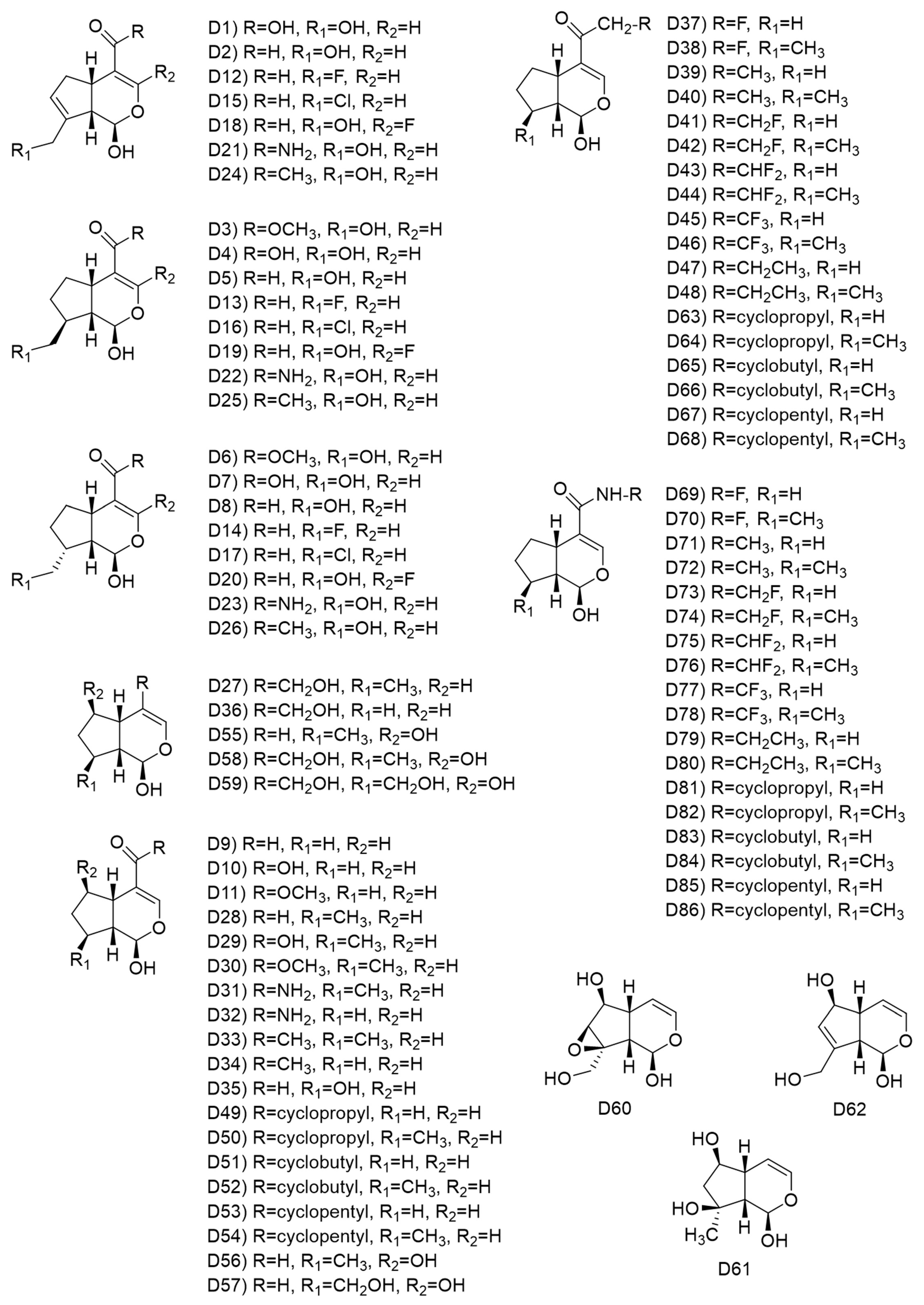
2.7. Ligand-Based Drug Design of Genipin Derivatives
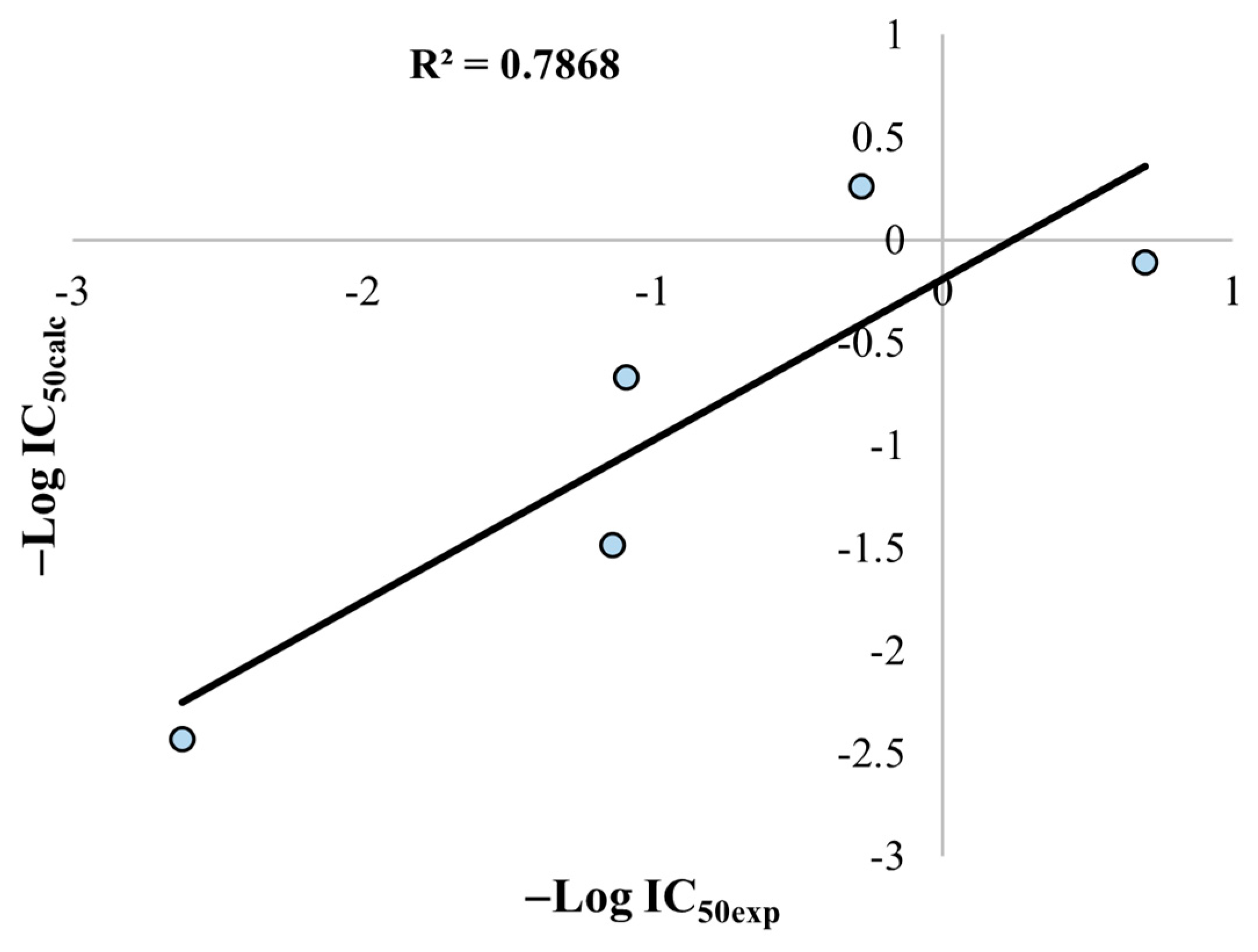
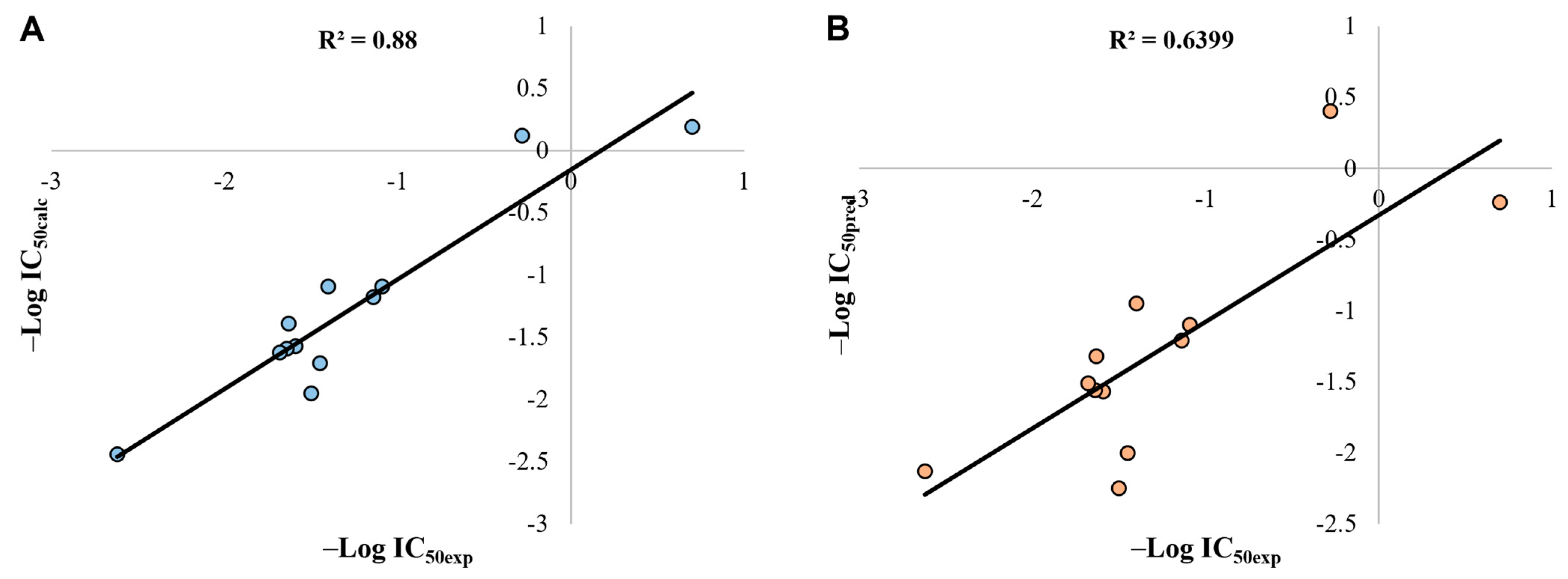
2.8. QSAR Analysis
3. Materials and Methods
3.1. In Vitro Cytotoxic Activity of Geniposide and Genipin
3.2. Searching of Cytotoxic Reports of Iridoids in Cervical Cancer
3.3. Conformation Analysis, Geometry Optimization, and Energy Calculation
3.4. Molecular Descriptors
3.5. Molecular Representation of Electron Density Properties
3.6. Molecular Similarity Analysis of Iridoids
3.7. QSAR Construction and Validation
Molecular Descriptors
3.8. Mathematical Model Generation
3.9. Statistical Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 5 July 2023).
- GLOBOCAN-Global Cancer Observatory. Available online: https://gco.iarc.fr/ (accessed on 10 July 2023).
- Okunade, K.S. Human papillomavirus and cervical cancer. J. Obstet. Gynaecol. 2020, 40, 602–608. [Google Scholar] [CrossRef]
- Anand, U.; Dey, A.; Chandel, A.K.S.; Sanyal, R.; Mishra, A.; Pandey, D.K.; De Falco, V.; Upadhyay, A.; Kandimalla, R.; Chaudhary, A.; et al. Cancer chemotherapy and beyond: Current status, drug candidates, associated risks and progress in targeted therapeutics. Genes Dis. 2022, 10, 1367–1401. [Google Scholar] [CrossRef]
- Çakır, C.; Kılıç, F.; Dur, R.; Yüksel, D.; Ünsal, M.; Korkmaz, V.; Kılıç, Ç.; Kimyon Cömert, G.; Boran, N.; Türkmen, O.; et al. Neoadjuvant chemotherapy for locally advanced stage (IB2-IIA2-IIB) cervical carcinoma: Experience of a tertiary center and comprehensive review of the literature. Turk. J. Obstet. Gynecol. 2021, 18, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liang, L.Z.; Cao, L.P.; Min, Z.; Liu, J.H. Clinical effects of irinotecan hydrochloride in combination with cisplatin as neoadjuvant chemotherapy in locally advanced cervical cancer. Gynecol Oncol. 2011, 123, 99–104. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y. Application of vincristine and cisplatin combined with intensity-modulated radiation therapy in the treatment of patients with advanced cervical cancer. Am. J. Transl. Res. 2021, 13, 13894–13901. [Google Scholar] [PubMed]
- Bajaj, A.; Gopalakrishnan, M.; Harkenrider, M.M.; Lurain, J.R.; Small, W., Jr. Advanced small cell carcinoma of the cervix-Successful treatment with concurrent etoposide and cisplatin chemotherapy and extended field radiation: A case report and discussion. Gynecol. Oncol. Rep. 2017, 23, 4–6. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, M.; Lee, S.; Jung, W.; Kim, B. Therapeutic Potential of Natural Products in Treatment of Cervical Cancer: A Review. Nutrients 2021, 13, 154. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect. 2021, 11, 5–13. [Google Scholar] [CrossRef]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural Products as Anticancer Agents: Current Status and Future Perspectives. Molecules 2022, 27, 8367. [Google Scholar] [CrossRef]
- Wang, C.; Gong, X.; Bo, A.; Zhang, L.; Zhang, M.; Zang, E.; Zhang, C.; Li, M. Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics. Molecules 2020, 25, 287. [Google Scholar] [CrossRef]
- Hussain, H.; Green, I.R.; Saleem, M.; Raza, M.L.; Nazir, M. Therapeutic Potential of Iridoid Derivatives: Patent Review. Inventions 2019, 4, 29. [Google Scholar] [CrossRef]
- Bianco, A. Recent developments in iridoids chemistry. Pure Appl. Chem. 1994, 86, 2335–2338. [Google Scholar] [CrossRef]
- Ornano, L.; Feroci, M.; Guarcini, L.; Venditti, A.; Bianco, A. Anti-HIV agents from nature: Natural compounds from Hypericum hircinum and carbocyclic nucleosides from iridoids. Stud. Nat. Prod. Chem. 2018, 56, 173–228. [Google Scholar] [CrossRef]
- Kim, C.W.; Choi, K.C. Potential roles of iridoid glycosides and their underlying mechanisms against diverse cancer growth and metastasis: Do they have an inhibitory effect on cancer progression? Nutrients 2021, 13, 2974. [Google Scholar] [CrossRef]
- Ndongwe, T.; Witika, B.A.; Mncwangi, N.P.; Poka, M.S.; Skosana, P.P.; Demana, P.H.; Summers, B.; Siwe-Noundou, X. Iridoid Derivatives as Anticancer Agents: An Updated Review from 1970–2022. Cancers 2023, 15, 770. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, H.; Wang, X.; Liu, Z. Lactobacillus raises in vitro anticancer effect of Geniposide in HSC-3 human oral squamous cell carcinoma cells. Exp. Ther. Med. 2017, 14, 4586–4594. [Google Scholar] [CrossRef]
- Bai, G.; Chen, B.; Xiao, X.; Li, Y.; Liu, X.; Zhao, D.; Zhang, L.; Zhao, D. Geniposide inhibits cell proliferation and migration in human squamous carcinoma cells via AMPK and JNK signaling pathways. Exp. Ther. Med. 2022, 24, 706. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Lentini, G. Plant-derived anticancer agents: Lessons from the pharmacology of Geniposide and its aglycone, Genipin. Biomedicines 2018, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Cao, H.L.; Xu, W.; Li, X.R.; Ren, Y.Q.; Du, L.F. Apoptosis induced by Genipin in human leukemia K562 cells: Involvement of c-Jun N-terminal kinase in G2/M arrest. Acta Pharmacol. Sin. 2011, 31, 519–527. [Google Scholar] [CrossRef]
- Kim, E.S.; Jeong, C.S.; Moon, A. Genipin, a constituent of Gardenia jasminoides Ellis, induces apoptosis and inhibits invasion in MDA-MB-231 breast cancer cells. Oncol. Rep. 2012, 27, 567–572. [Google Scholar] [CrossRef]
- Ko, H.; Kim, J.M.; Kim, S.J.; Shim, S.H.; Ha, C.H.; Chang, H.I. Induction of apoptosis by Genipin inhibits cell proliferation in AGS human gastric cancer cells via Egr1/p21 signaling pathway. Bioorganic Med. Chem. Lett. 2015, 25, 4191–4196. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, T.B.; Jia, D.H.; Sun, W.Q.; Wang, C.L.; Gu, A.Z.; Yang, X.M. Genipin inhibits the growth of human bladder cancer cells via inactivation of PI3K/Akt signaling. Oncol. Lett. 2018, 15, 2619–2624. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Shen, H.; Tang, F.R.; Arfuso, F.; Rajesh, M.; Wang, L.; Kumar, A.P.; Bian, J.; Goh, B.C.; Bishayee, A.; et al. Potential role of Genipin in cancer therapy. Pharmacol. Res. 2018, 133, 195–200. [Google Scholar] [CrossRef]
- Maggiora, G.; Vogt, M.; Stumpge, D.; Bajorath, J. Molecular Similarity in Medicinal Chemistry. J. Med. Chem. 2014, 57, 3186–3204. [Google Scholar] [CrossRef]
- Daina, A.; Röhrig, U.F.; Zoete, V. Computer-aided drug design for cancer therapy. In Systems Medicine: Integrative, Qualitative and Computation Approaches, 1st ed.; Wolkenhauer, O., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 386–401. [Google Scholar] [CrossRef]
- Zuñiga Martinez, M.D.L.; López Mendoza, C.M.; Tenorio Salazar, J.; García Carrancá, A.M.; Cerbón Cervantes, M.A.; Alcántara-Quintana, L.E. Establishment, authenticity, and characterization of cervical cancer cell lines. Mol. Cell. Oncol. 2022, 9, 2078628. [Google Scholar] [CrossRef]
- Neri-Numa, I.A.; Pessôa, M.G.; Arruda, H.S.; Pereira, G.A.; Paulino, B.N.; Angolini, C.F.F.; Ruiz, A.L.T.G.; Pastore, G.M. Genipap (Genipa americana L.) fruit extract as a source of antioxidant and antiproliferative iridoids. Food Res. Int. 2020, 134, 109252. [Google Scholar] [CrossRef]
- Hwang, H.; Kim, C.; Kim, S.M.; Kim, W.S.; Choi, S.H.; Chang, I.M.; Ahn, K.S. The hydrolyzed products of iridoid glycoside with β-glucosidase treatment exert anti-proliferative effects through suppression of STAT3 activation and STAT3-regulated gene products in several human cancer cells. Pharm. Biol. 2012, 50, 8–17. [Google Scholar] [CrossRef]
- Marcotullio, M.C.; Loizzo, M.R.; Messina, F.; Temperini, A.; Tundis, R.; Menichini, F.; Curini, M. Bioassay-guided fractionation of Euphrasia pectinata Ten. and isolation of iridoids with antiproliferative activity. Phytochem. Lett. 2015, 12, 252–256. [Google Scholar] [CrossRef]
- Feng, S.X.; Yi, B.; Zhang, M.; Xu, J.; Lin, H.; Xu, W.T. Iridoid glycosides from Callicarpa nudiflora Hook. Nat. Prod. Res. 2017, 31, 181–189. [Google Scholar] [CrossRef]
- Wang, C.; Xin, P.; Wang, Y.; Zhou, X.; Wei, D.; Deng, C.; Sun, S. Iridoids and sfingolipids from Hedyotis diffusa. Fitoterapia 2018, 124, 152–159. [Google Scholar] [CrossRef]
- Cao, H.; Feng, Q.; Xu, W.; Li, X.; Kang, Z.; Ren, Y.; Du, L. Genipin induced apoptosis associated with activation of the c-Jun NH2-terminal kinase and p53 protein in HeLa cells. Biol. Pharm. Bull. 2010, 33, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Di, L.; Gao, W.C.; Wang, K.J.; Zu, L.B. Cytotoxic iridoids from the roots of Patrinia scabra. J. Nat. Prod. 2012, 72, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Gehle, D.; Dey, S.K.; Nahar, N.; Mosihuzzaman, M.; Sutana, N.; Sohrab, M.H.; Stephens, P.J.; Pan, J.J.; Sasse, F. Prismatomerin, a new iridoid from Prismatomeris tetranda. Structure elucidation, determination of absolute configuration, and cytotoxicity. J. Nat. Prod. 2007, 70, 1339–1343. [Google Scholar] [CrossRef]
- Kırmızıbekmez, H.; Kúsz, N.; Bérdi, P.; Zupkó, I.; Hohmann, J. New iridoids from the roots of Valeriana dioscoridis Sm. Fitoterapia 2018, 130, 73–78. [Google Scholar] [CrossRef]
- Sheng, L.; Yang, Y.; Zhang, Y.; Li, N. Chemical constituents of Patrinia heterophylla Bunge and selective cytotoxicity against six human tumor cells. J. Ethnopharmacol. 2019, 236, 129–135. [Google Scholar] [CrossRef]
- Saidi, I.; Baccari, W.; Marchal, A.; Waffo-Téguo, W.; Harrath, A.H.; Mansour, L.; Jannet, H.B. Iridoid glycosides from the Tunisian Citharexylum spinosum L.: Isolation, structure elucidation, biological evaluation, molecular docking and SAR analysis. Ind. Crops Prod. 2020, 151, 112440. [Google Scholar] [CrossRef]
- Saidi, I.; Nimbarte, V.D.; Schwalbe, H.; Wafoo-Téguo, P.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Jannet, H.B. Anti-tyrosinase, anti-cholinesterase and cytotoxic activities of extracts and phytochemicals from the Tunisian Citharexylum spinosum L.: Molecular docking and SAR analysis. Bioorganic Chem. 2020, 102, 104093. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, H.; Fu, Y.; Ma, K.; Ma, X.; Wang, J. Gentiopicroside inhibits cell growth and migration on cervical cancer via the recriprocal MAPK/Akt signaling pathways. Nutr. Cancer 2021, 73, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Ma, L.; Zhang, Q.; Yang, R.; Hou, G.; Wang, Y.; Zhao, F. Characterization, crystal structure and cytotoxic activity of a rare iridoid glycoside from Lonicera saccata. Acta Crystallogr. Sect. C Struct. Chem. 2020, C76, 269–275. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Minoshima, Y.; Kishimoto, Y.; Chen, I.S.; Takahashi, H.; Esumi, T. Iridoid glucosides and p-coumaroyl iridoids from Viburnum luzonicum and their cytotoxicity. J. Nat. Prod. 2004, 67, 1833–1838. [Google Scholar] [CrossRef]
- Discovery Studio 2021. Available online: https://discover.3ds.com/discovery-studio-visualizer-download (accessed on 5 August 2023).
- Yang, Y.; Yang, Y.; Hou, J.; Ding, Y.; Zhang, T.; Zhang, Y.; Wang, J.; Shi, C.; Fu, W.; Cai, Z. The hydroxyl at position C1 of Genipin is the active inhibitory group that affects mitochondrial uncoupling protein 2 in Panc-1 cells. PLoS ONE 2016, 11, e0147026. [Google Scholar] [CrossRef]
- Brown, S.; Clastre, M.; Courdavault, V.; O’Connor, S.E. De novo production of the plant-derived alkaloid strictosidine in yeast. Proc. Natl. Acad. Sci. USA 2015, 112, 3205–3210. [Google Scholar] [CrossRef] [PubMed]
- Nagatoshi, M.; Terasaka, K.; Nagatsu, A.; Mizukami, H. Iridoid-specific glucosyltransferase from Gardenia jasminoides. J. Biol. Chem. 2011, 286, 32866–32874. [Google Scholar] [CrossRef] [PubMed]
- Pungitore, C.R.; García, C.; Sotero Martín, V.; Tonn, C.E. Inhibition of Taq DNA polymerase by iridoid aglycone derivates. Cell. Mol. Biol. 2012, 58, 1786–1790. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxic assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Peña-Morán, O.A.; Villarreal, M.L.; Álvarez-Berber, L.; Meneses-Acosta, A.; Rodríguez-López, V. Cytotoxicity, Post-Treatment Recovery, and Selectivity Analysis of Naturally Occurring Podophyllotoxins from Bursera fagaroides var. fagaroides on Breast Cancer Cell Lines. Molecules 2016, 21, 1013. [Google Scholar] [CrossRef]
- Sanchez-Carranza, J.N.; González-Maya, L.; Razo-Hernández, R.S.; Salas-Vidal, E.; Nolasco-Quintana, N.Y.; Clemente-Soto, A.F.; García-Arizmendi, L.; Sánchez-Ramos, M.; Marquina, S.; Alvarez, L. Achillin Increases Chemosensitivity to Paclitaxel, Overcoming Resistance and Enhancing Apoptosis in Human Hepatocellular Carcinoma Cell Line Resistant to Paclitaxel (Hep3B/PTX). Pharmaceutics 2019, 11, 512. [Google Scholar] [CrossRef]
- Halgren, T.A. Merck Molecular Force Field. I. Basis, form, scope, parameterization and performance of MMFF94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Stewart, J.P.J. Optimization of parameters for semiempirical methods I. Method. J. Comput. Chem. 1989, 10, 209–220. [Google Scholar] [CrossRef]
- Stephens, P.J.; Devlin, F.; Chabalowski, C.F.; Frisch, M.J. Ab Initio calculation of vibrational absorption and circular dichroism spectra using density functional force fields: A comparison of local, nonlocal, and hybrid density functionals. J. Phys. Chem. 1994, 45, 11623–11627. [Google Scholar] [CrossRef]
- Hay, P.J.; Wadt, W.R. Ab initio effective core potentials for molecular calculations. Potentials for main group elements Na to Bi. J. Chem. Phys. 1985, 82, 284–298. [Google Scholar] [CrossRef]
- Chambers, C.C.; Hawkins, G.D.; Cramer, C.J.; Truhlar, D.G. Model for aqueous solvation based on class IV atomic charges and first solvation shell effects. J. Chem. Phys. 1996, 100, 16385–16398. [Google Scholar] [CrossRef]
- Petersson, G.A.; Tensfeldt, T.G.; Montgomery, J.A. A complete basis set model chemistry. III. The complete basis set-quadratic configuration interaction family of methods. J. Chem. Phys. 1991, 94, 6091–6101. [Google Scholar] [CrossRef]
- Wavefunction, Inc. Spartan’20. Available online: https://www.wavefun.com (accessed on 30 July 2023).
- Virtual Computational Chemistry Laboratory. Available online: https://vcclab.org/lab/alogps/ (accessed on 25 August 2023).
- Razo-Hernández, R.S.; Pineda-Urbina, K.; Velazco-Medel, M.A.; Villanueva-García, M.; Sumaya-Martínez, M.T.; Martínez-Martínez, F.J.; Gómez-Sandoval, Z. QSAR study of the DPPH· radical scavenging activity of coumarin derivatives and xanthine oxidase inhibition by molecular docking. Cent. Eur. J. Chem. 2014, 12, 1067–1080. [Google Scholar] [CrossRef]
- Marquina, S.; Maldonado-Santiago, M.; Sánchez-Carranza, J.N.; Antúnez-Mojica, M.; González-Maya, L.; Razo-Hernández, R.S.; Alvarez, L. Design, synthesis and QSAR study of 2′-hydroxy-4′-alkoxy chalcone derivatives that exert cytotoxic activity by the mitochondrial apoptotic pathway. Bioorganic Med. Chem. 2019, 27, 43–54. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Geniposide | Genipin | Cisplatin | Podophyllotoxin |
|---|---|---|---|---|
| CaSki | >1000 | 65.930 ± 4.420 | 0.640 ± 0.080 | 0.025 ± 0.001 |
| CaLo | >1000 | 58.970 ± 9.040 | 0.240 ± 0.020 | 0.014 ± 0.002 |
| INBL | >1000 | 178.800 ± 12.990 | 1.340 ± 0.160 | 0.011 ± 0.001 |
| HaCaT 1 | >1000 | 106.250 ± 9.290 | 1.150 ± 0.140 | 0.050 ± 0.006 |
| Compound | Cell Line | Exposure Time (h) | IC50 (µM) |
|---|---|---|---|
| Euphrasin 1 | HeLa | 48 | 0.20 |
| Campsinol 1 | HeLa | 48 | 1.90 |
| Artselaenin B 1 | HeLa | 48 | 12.30 |
| Artselaenin A 1 | HeLa | 48 | 13.80 |
| Pulchelloside I 2 | HeLa | 48 | 25.22 |
| Catalpol 3 | HeLa | 48 | 28.20 |
| Lamiide 2 | HeLa | 48 | 31.96 |
| Spinomannoside 2 | HeLa | 48 | 38.89 |
| 5-deoxypulchelloside I 2 | HeLa | 48 | 42.47 |
| Geniposidic acid 4 | HeLa | 48 | 43.60 |
| 10-O-(E)-p-coumaroylgeniposidic acid 3 | HeLa | 48 | 48.10 |
| Genipin | HeLa | 48 | 419.00 5 |
| Geniposide | HeLa | 48 | NA 5 |
| Ligand | IC50 (µM) | GAPHOMO-LUMO (kcal) | Dipole moment (debye) | Area (Å2) | Volume (Å3) | PSA (Å2) | Ovality | LogP | Polarizability | HBD Count | HBA Count |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Euphrasin | 0.20 | −123.01 | 4.81 | 234.27 | 216.65 | 44.645 | 1.34 | −0.10 | 57.69 | 1 | 4 |
| Campsinol | 1.90 | −122.30 | 5.78 | 236.86 | 216.59 | 46.922 | 1.36 | −0.10 | 57.69 | 1 | 4 |
| Artselaenin B | 12.3 | −125.40 | 4.67 | 227.13 | 208.64 | 26.808 | 1.34 | 1.11 | 57.01 | 0 | 3 |
| Artselaenin A | 13.8 | −121.90 | 4.20 | 228.14 | 209.46 | 27.236 | 1.34 | 1.11 | 57.11 | 0 | 3 |
| Genipin | 419.00 | −126.18 | 1.26 | 239.61 | 219.74 | 64.589 | 1.36 | −0.26 | 57.91 | 2 | 5 |
| Ligand | IC50 (µM) | GAPHOMO-LUMO (kcal) | Dipole Moment (debye) | Area (Å2) | Volume (Å3) | PSA (Å2) | Ovality | LogP | Polarizability | HBD Count | HBA Count |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genipin | 419.00 | −126.18 | 1.26 | 239.61 | 219.74 | 64.589 | 1.36 | −0.26 | 57.91 | 2 | 5 |
| Geniposide | NA | −128.37 | 3.41 | 371.47 | 355.38 | 119.843 | 1.53 | −2.00 | 68.89 | 5 | 9 |
| Ligand | IC50 (µM) | GAPHOMO-LUMO (kcal) | Dipole Moment (debye) | Area (Å2) | Volume (Å3) | PSA (Å2) | Ovality | LogP | Polarizability | HBD Count | HBA Count |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pulchelloside I | 25.22 | −121.84 | 5.95 | 383.69 | 372.30 | 148.544 | 1.53 | −3.30 | 70.33 | 7 | 11 |
| Catalpol | 28.20 | −144.80 | 2.50 | 324.51 | 320.55 | 126.832 | 1.43 | −3.24 | 65.89 | 6 | 10 |
| Laamide | 31.96 | −116.96 | 7.99 | 392.42 | 373.46 | 158.057 | 1.56 | −3.48 | 70.47 | 7 | 11 |
| Spinomannoside | 38.89 | −124.51 | 4.28 | 381.33 | 367.08 | 140.269 | 1.54 | −2.51 | 69.87 | 5 | 10 |
| 5-deoxypulchelloside I | 42.47 | −125.49 | 3.32 | 374.98 | 365.07 | 134.805 | 1.52 | −2.51 | 69.70 | 6 | 10 |
| Geniposidic acid * | 43.60 | −85.84 | 12.02 | 353.50 | 335.17 | 139.852 | 1.51 | -- | 67.68 | 5 | 10 |
| 10-O-(E)-p-coumaroylgeniposidic acid * | 48.10 | −61.97 | 16.64 | 481.01 | 476.36 | 147.201 | 1.63 | -- | 79.38 | 5 | 11 |
| Geniposide | NA | −128.37 | 3.41 | 371.47 | 355.38 | 119.843 | 1.53 | −2.00 | 68.89 | 5 | 9 |
| Ligand | (debye) | θ | φ | IC50 (µM) |
|---|---|---|---|---|
| Euphrasin | 4.81 | 0.8097 | 1.6749 | 0.20 |
| Campsinol | 5.78 | −1.0080 | 1.3527 | 1.90 |
| Artselaenin B | 4.67 | 0.9716 | 1.3933 | 12.3 |
| Artselaenin A | 4.20 | −1.3515 | 1.1341 | 13.8 |
| Genipin | 1.26 | 1.5078 | 2.2265 | 419.00 |
| Ligand | (debye) | ||||
|---|---|---|---|---|---|
| Euphrasin | 4.81 | −19.944 | −1.21 | 0.048 | 0.20 |
| Campsinol | 5.78 | −17.667 | −1.21 | 0.048 | 1.90 |
| Artselaenin B | 4.67 | −37.781 | −2.23 | 0.810 | 12.30 |
| Artselaenin A | 4.20 | −37.353 | −2.23 | 0.640 | 13.80 |
| Pulchelloside I | 5.95 | 83.955 | −0.50 | 154.256 | 25.22 |
| Catalpol | 2.50 | 62.243 | −0.19 | 63.680 | 28.20 |
| Lamiide | 7.99 | 93.468 | −0.69 | 157.754 | 31.96 |
| Spinomannoside | 4.28 | 75.680 | −0.73 | 143.042 | 38.89 |
| 5-deoxypulchelloside I | 3.32 | 70.216 | −0.73 | 139.004 | 42.47 |
| Geniposidic acid 1 | 12.02 | 75.260 | −1.22 | 95.453 | 43.60 |
| 10-O-(E)-p-coumaroylgeniposidic acid 1 | 16.64 | 82.610 | −2.86 | 460.961 | 48.10 |
| Genipin | 1.26 | 0 | −1.50 | 0 | 419.00 |
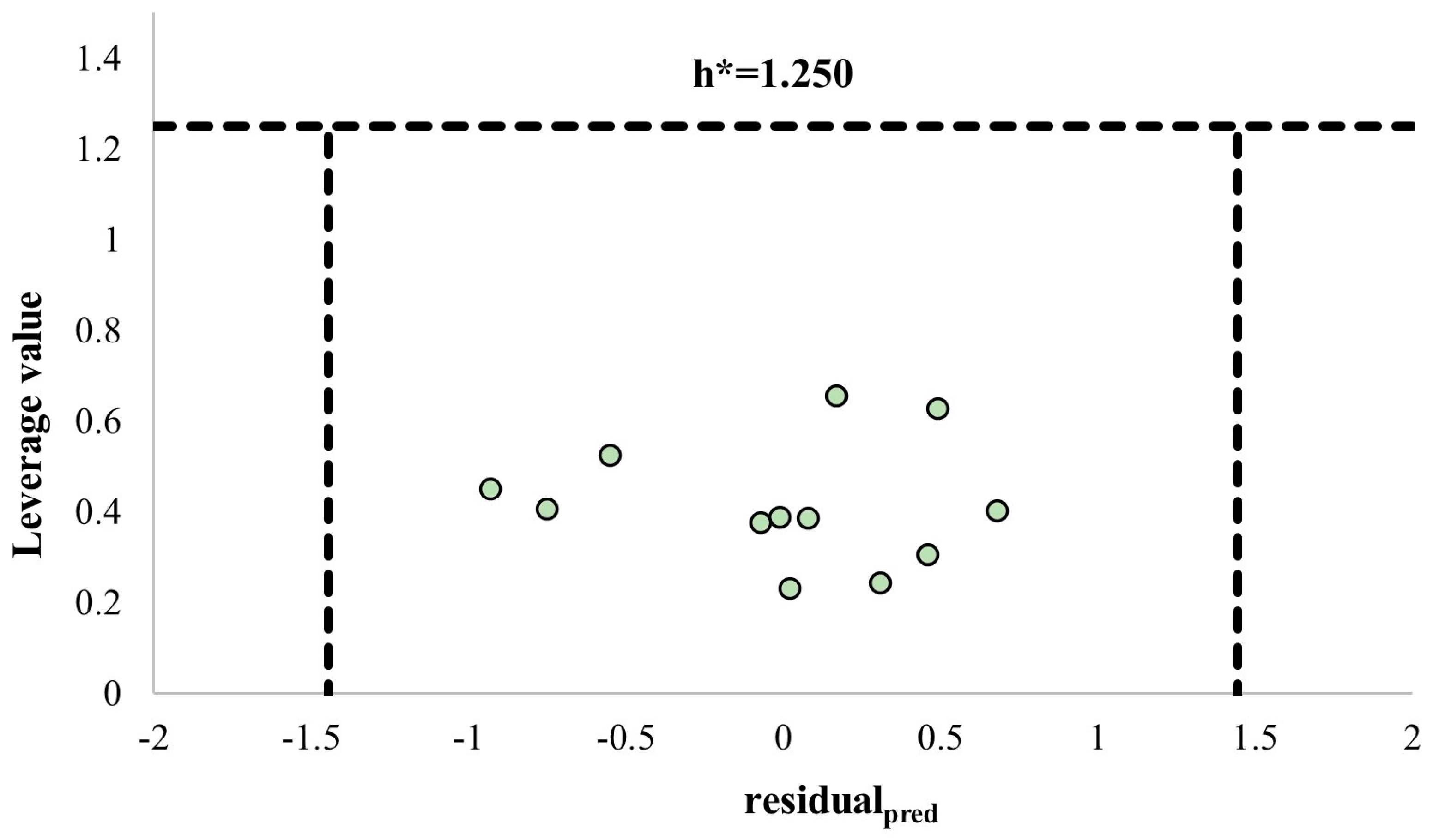
| Ligand | Yexp | Ycalc | Ypred | Hat | residualcalc | residualpred |
|---|---|---|---|---|---|---|
| Euphrasin | 0.70 | 0.19 | −0.24 | 0.451 | −0.51 | −0.93 |
| Campsinol | −0.28 | 0.12 | 0.40 | 0.403 | 0.40 | 0.68 |
| Artselaenin B | −1.09 | −1.09 | −1.10 | 0.389 | 0 | −0.01 |
| Artselaenin A | −1.14 | −1.18 | −1.21 | 0.377 | −0.04 | −0.07 |
| Pulchelloside I | −1.40 | −1.09 | −0.95 | 0.305 | 0.32 | 0.46 |
| Catalpol | −1.45 | −1.71 | −2.00 | 0.524 | −0.26 | −0.55 |
| Lamiide | −1.50 | −1.95 | −2.25 | 0.407 | −0.44 | −0.75 |
| Spinomannoside | −1.59 | −1.57 | −1.57 | 0.231 | 0.02 | 0.02 |
| 5-deoxypulchelloside I | −1.63 | −1.39 | −1.32 | 0.244 | 0.24 | 0.31 |
| Geniposidic acid 1 | −1.64 | −1.59 | −1.56 | 0.387 | 0.05 | 0.08 |
| 10-O-(E)-p-coumaroylgeniposidic acid 1 | −1.68 | −1.62 | −1.51 | 0.656 | 0.06 | 0.17 |
| Genipin | −2.62 | −2.44 | −2.13 | 0.627 | 0.18 | 0.49 |
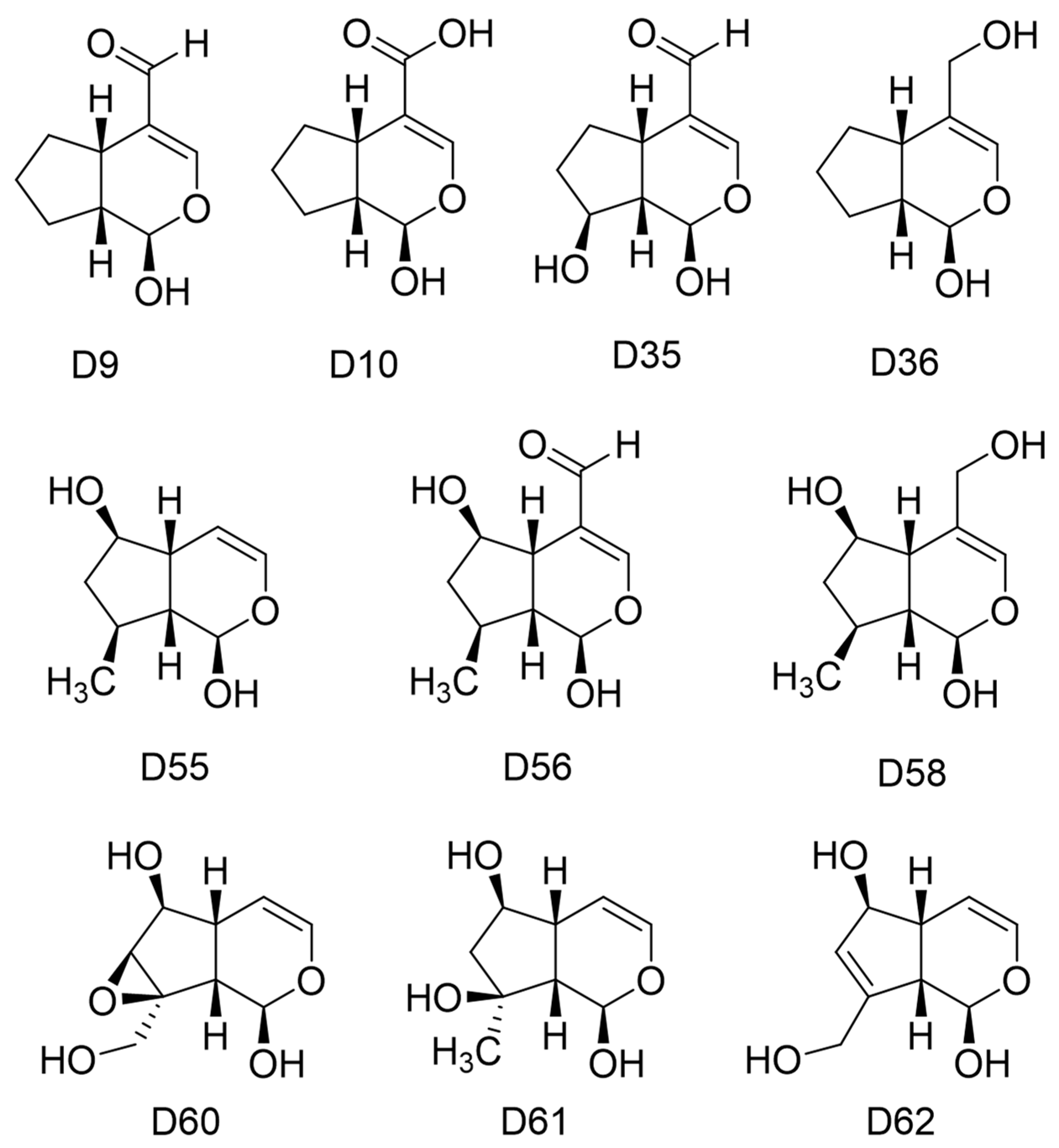
| Ligand | GAPHOMO-LUMO (kcal) | Dipole Moment (debye) | Area (Å2) | Volume (Å3) | PSA (Å2) | Ovality | LogP | Polarizability | HBD Count | HBA Count |
|---|---|---|---|---|---|---|---|---|---|---|
| D9 | −123.61 | 4.80 | 187.58 | 170.55 | 42.052 | 1.26 | 0.42 | 53.94 | 1 | 3 |
| D10 1 | −599.13 | 12.32 | 190.51 | 175.42 | 54.222 | 1.26 | -- | 54.14 | 1 | 4 |
| D35 | −123.99 | 3.36 | 197.34 | 178.62 | 60.872 | 1.29 | −0.67 | 54.59 | 2 | 4 |
| D36 | −138.78 | 0.69 | 196.25 | 176.32 | 47.478 | 1.29 | 0.55 | 54.25 | 2 | 3 |
| D55 | −139.52 | 1.70 | 196.35 | 176.4 | 47.351 | 1.29 | 0.61 | 54.25 | 2 | 3 |
| D56 | −118.90 | 4.81 | 213.74 | 196.37 | 57.202 | 1.31 | −0.41 | 56.08 | 2 | 4 |
| D58 | −140.01 | 1.99 | 219.39 | 201.1 | 61.709 | 1.32 | −0.28 | 56.25 | 3 | 4 |
| D60 | −144.58 | 2.30 | 201.54 | 184.95 | 73.770 | 1.28 | −1.50 | 54.89 | 3 | 5 |
| D61 | −145.63 | 3.19 | 207.72 | 190.41 | 80.817 | 1.30 | −1.25 | 55.33 | 4 | 5 |
| D62 | −135.25 | 0.86 | 196.11 | 178.97 | 61.665 | 1.28 | −0.79 | 54.50 | 3 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-López, D.; Razo-Hernández, R.S.; Millán-Pacheco, C.; Leyva-Peralta, M.A.; Peña-Morán, O.A.; Sánchez-Carranza, J.N.; Rodríguez-López, V. Ligand-Based Drug Design of Genipin Derivatives with Cytotoxic Activity against HeLa Cell Line: A Structural and Theoretical Study. Pharmaceuticals 2023, 16, 1647. https://doi.org/10.3390/ph16121647
López-López D, Razo-Hernández RS, Millán-Pacheco C, Leyva-Peralta MA, Peña-Morán OA, Sánchez-Carranza JN, Rodríguez-López V. Ligand-Based Drug Design of Genipin Derivatives with Cytotoxic Activity against HeLa Cell Line: A Structural and Theoretical Study. Pharmaceuticals. 2023; 16(12):1647. https://doi.org/10.3390/ph16121647
Chicago/Turabian StyleLópez-López, Diana, Rodrigo Said Razo-Hernández, César Millán-Pacheco, Mario Alberto Leyva-Peralta, Omar Aristeo Peña-Morán, Jessica Nayelli Sánchez-Carranza, and Verónica Rodríguez-López. 2023. "Ligand-Based Drug Design of Genipin Derivatives with Cytotoxic Activity against HeLa Cell Line: A Structural and Theoretical Study" Pharmaceuticals 16, no. 12: 1647. https://doi.org/10.3390/ph16121647
APA StyleLópez-López, D., Razo-Hernández, R. S., Millán-Pacheco, C., Leyva-Peralta, M. A., Peña-Morán, O. A., Sánchez-Carranza, J. N., & Rodríguez-López, V. (2023). Ligand-Based Drug Design of Genipin Derivatives with Cytotoxic Activity against HeLa Cell Line: A Structural and Theoretical Study. Pharmaceuticals, 16(12), 1647. https://doi.org/10.3390/ph16121647