

Daphnetin Improves Neuropathic Pain by Inhibiting the Expression of Chemokines and Inflammatory Factors in the Spinal Cord and Interfering with Glial Cell Polarization

 ,
,
Abstract
:1. Introduction
2. Results
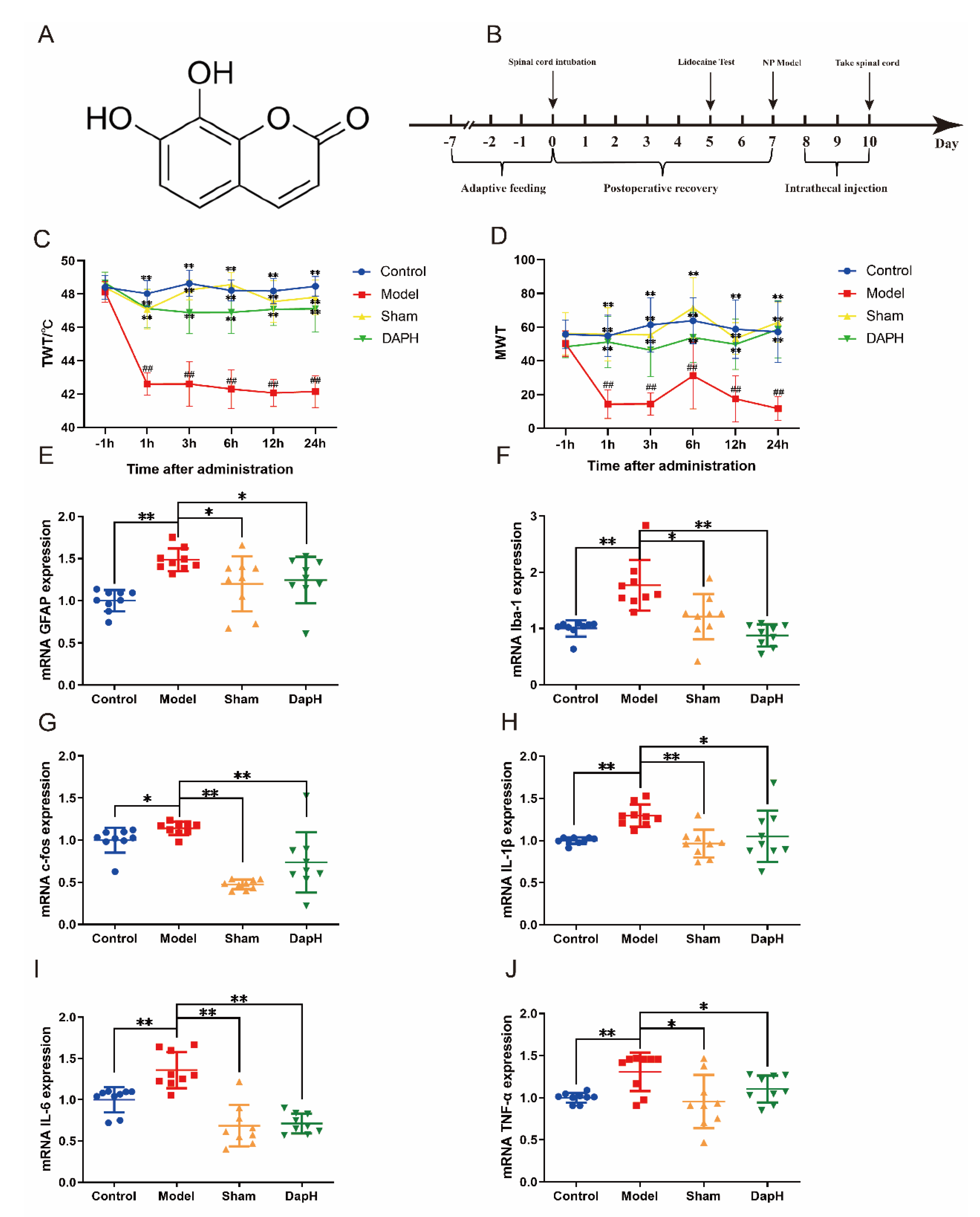
2.1. Daphnetin Improves Hyperalgesia in NP Rats
2.2. Daphnetin Inhibits the Activation of Glial Cells and Neurons in the Spinal Cord of NP Rats
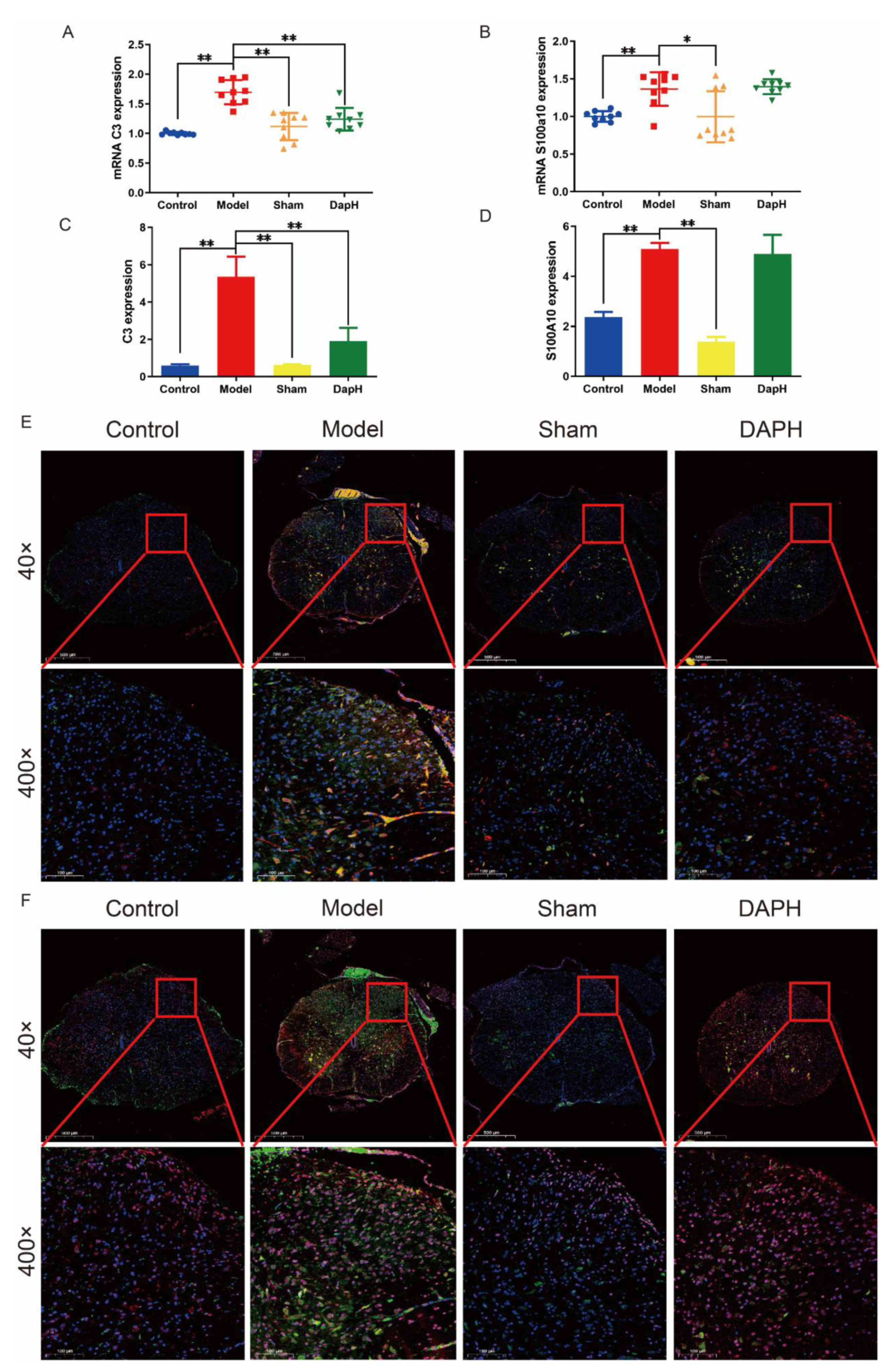
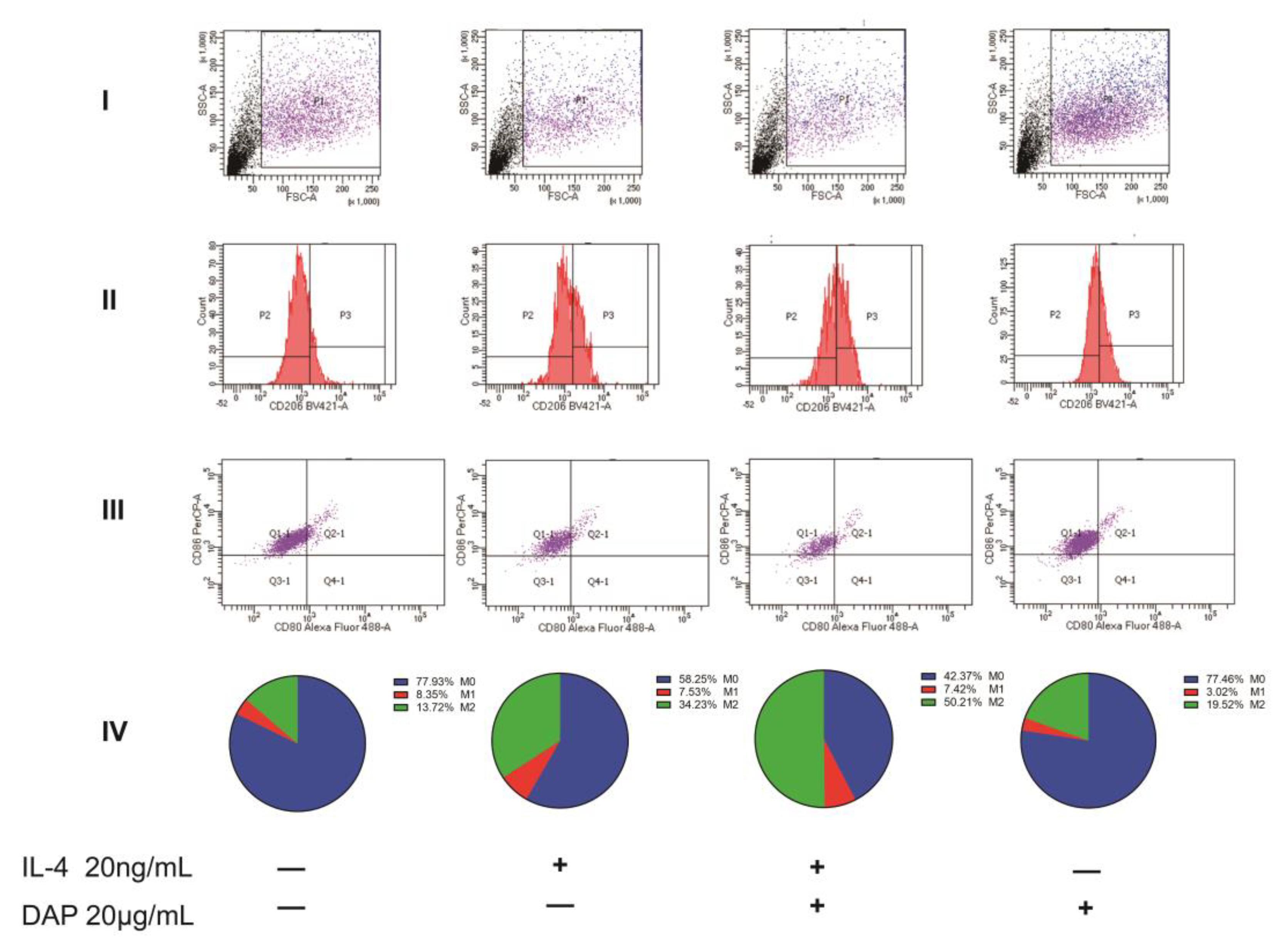
2.3. Daphnetin Inhibits the Polarization of A1 Astrocytes in the Spinal Cord of NP Rats
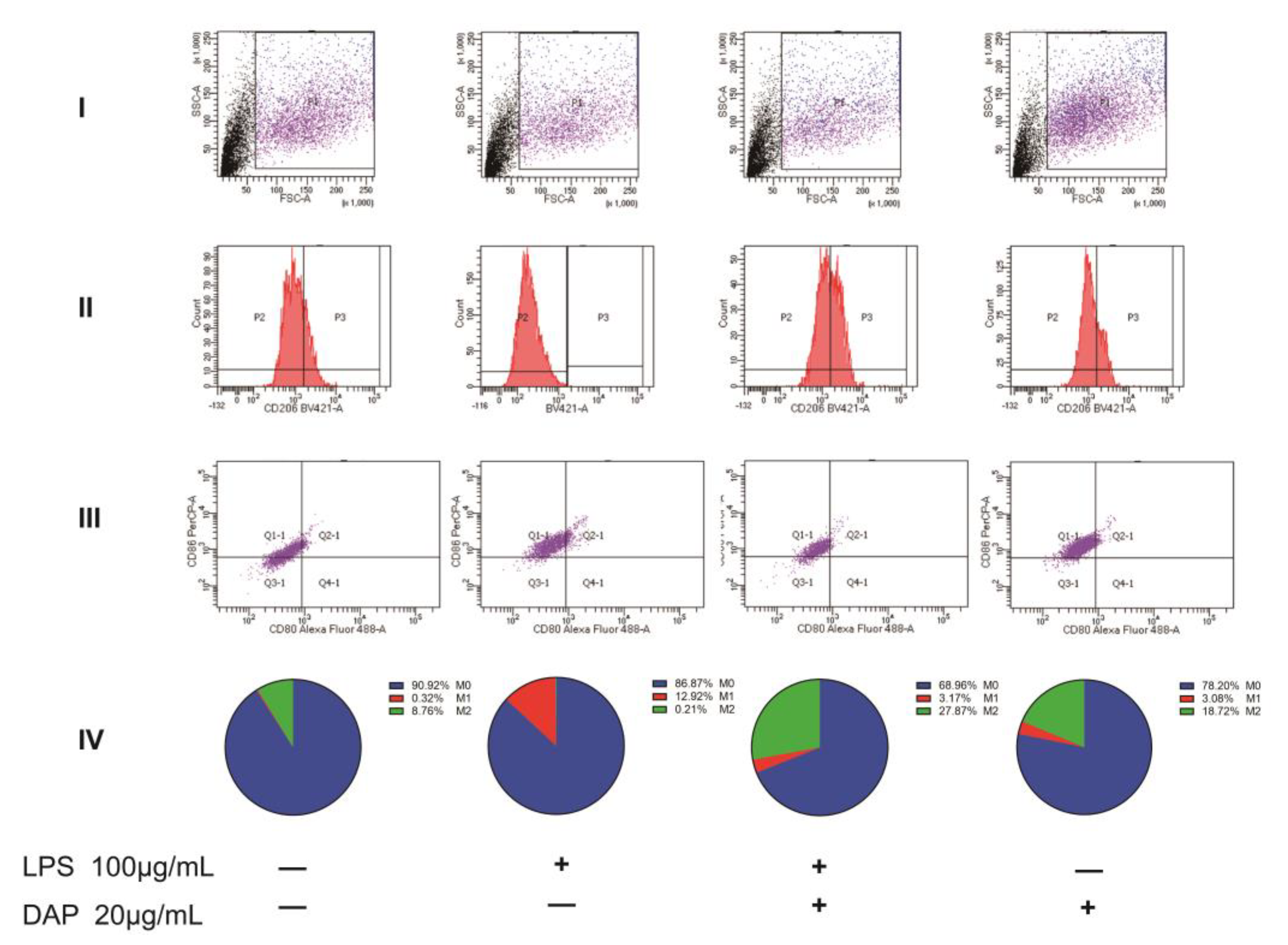
2.4. Daphnetin Inhibits the Polarization of M1 Microglia in the Spinal Cord of NP Rats
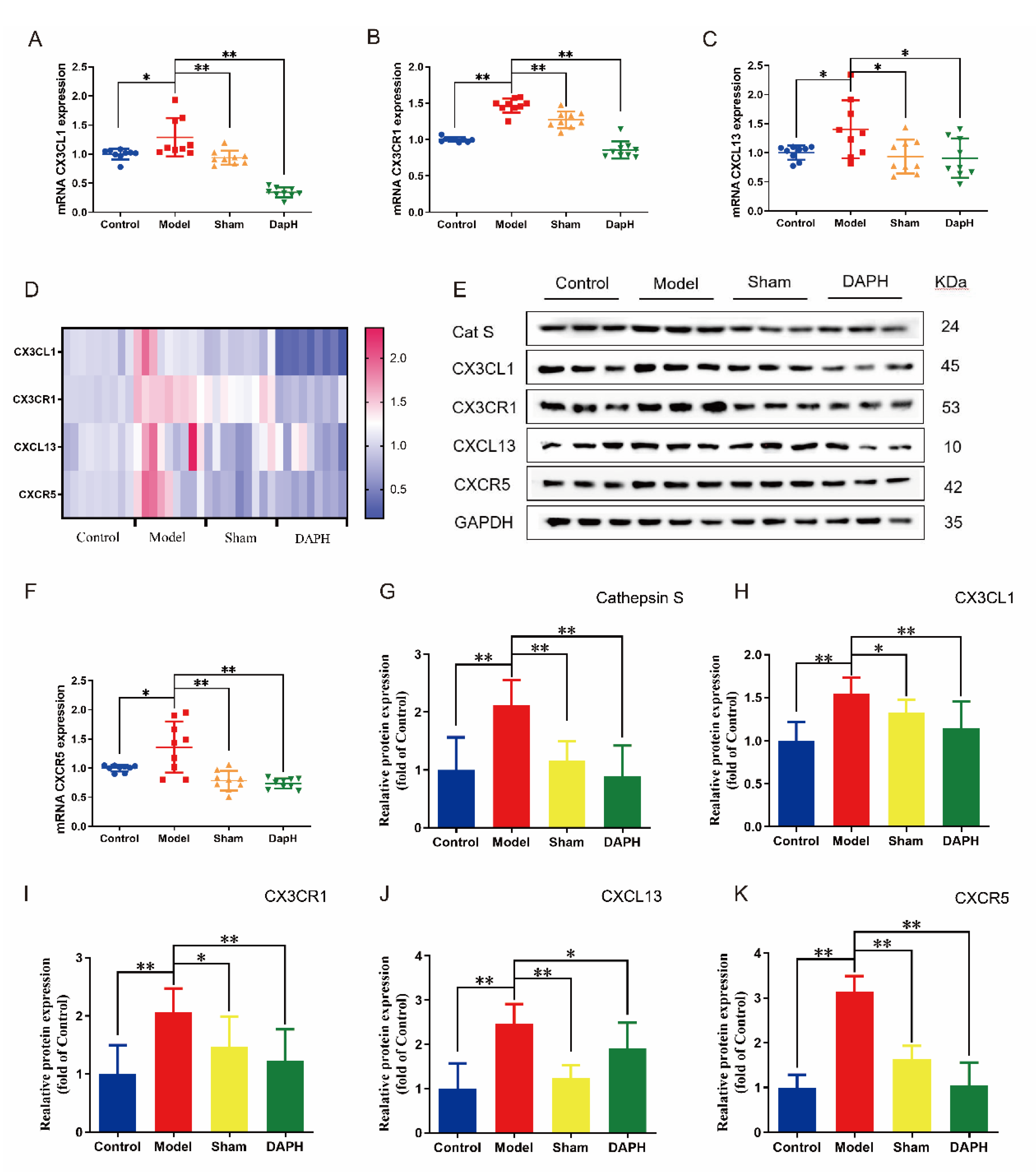
2.5. Daphnetin Inhibits the Expression of CX3CL1/CX3CR1 and CXCL13/CXCR5 Chemokine Pairs in the Spinal Cord of NP Rats
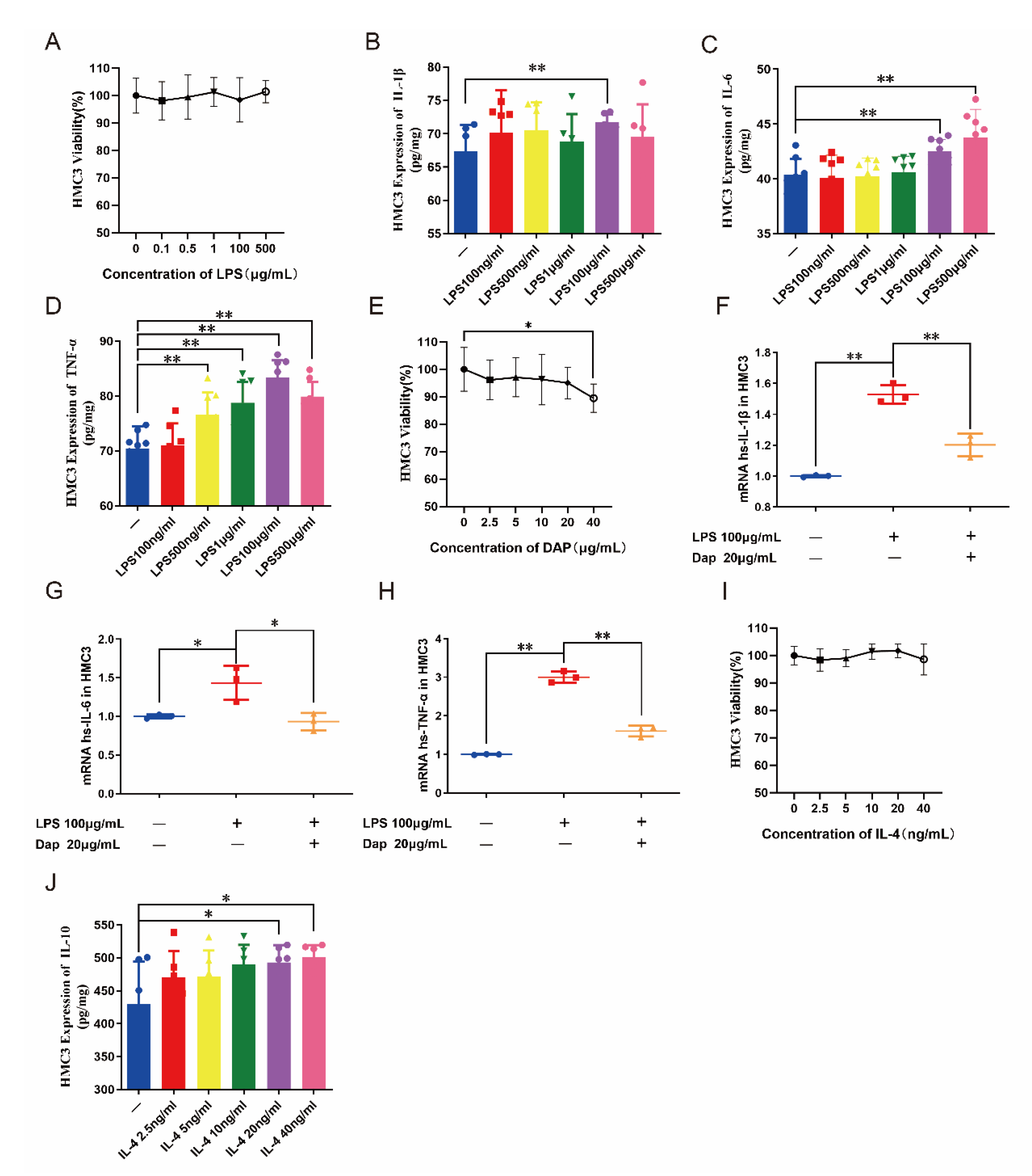
2.6. Intervention of Daphnetin on the Polarization of HMC3 Cells
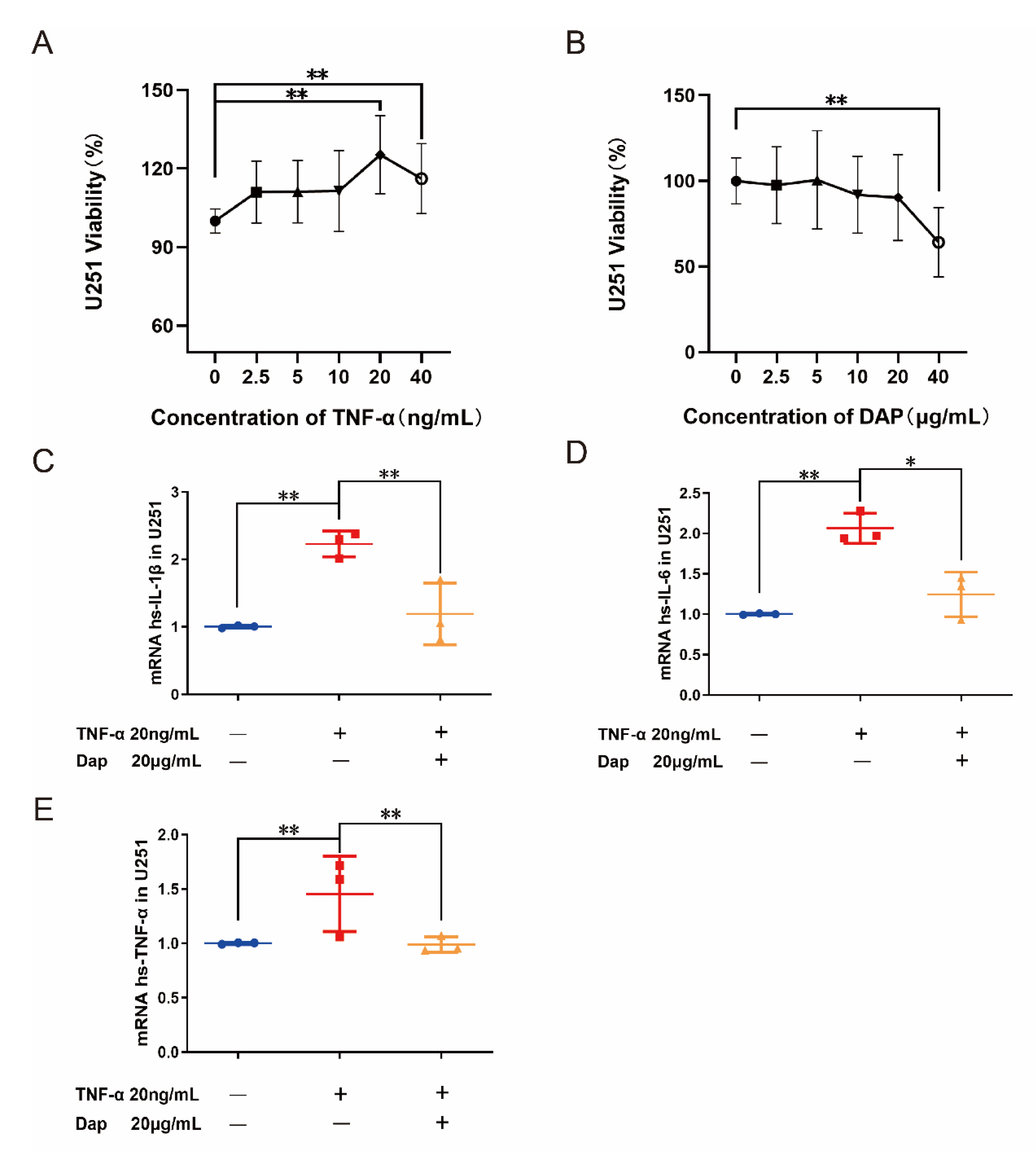
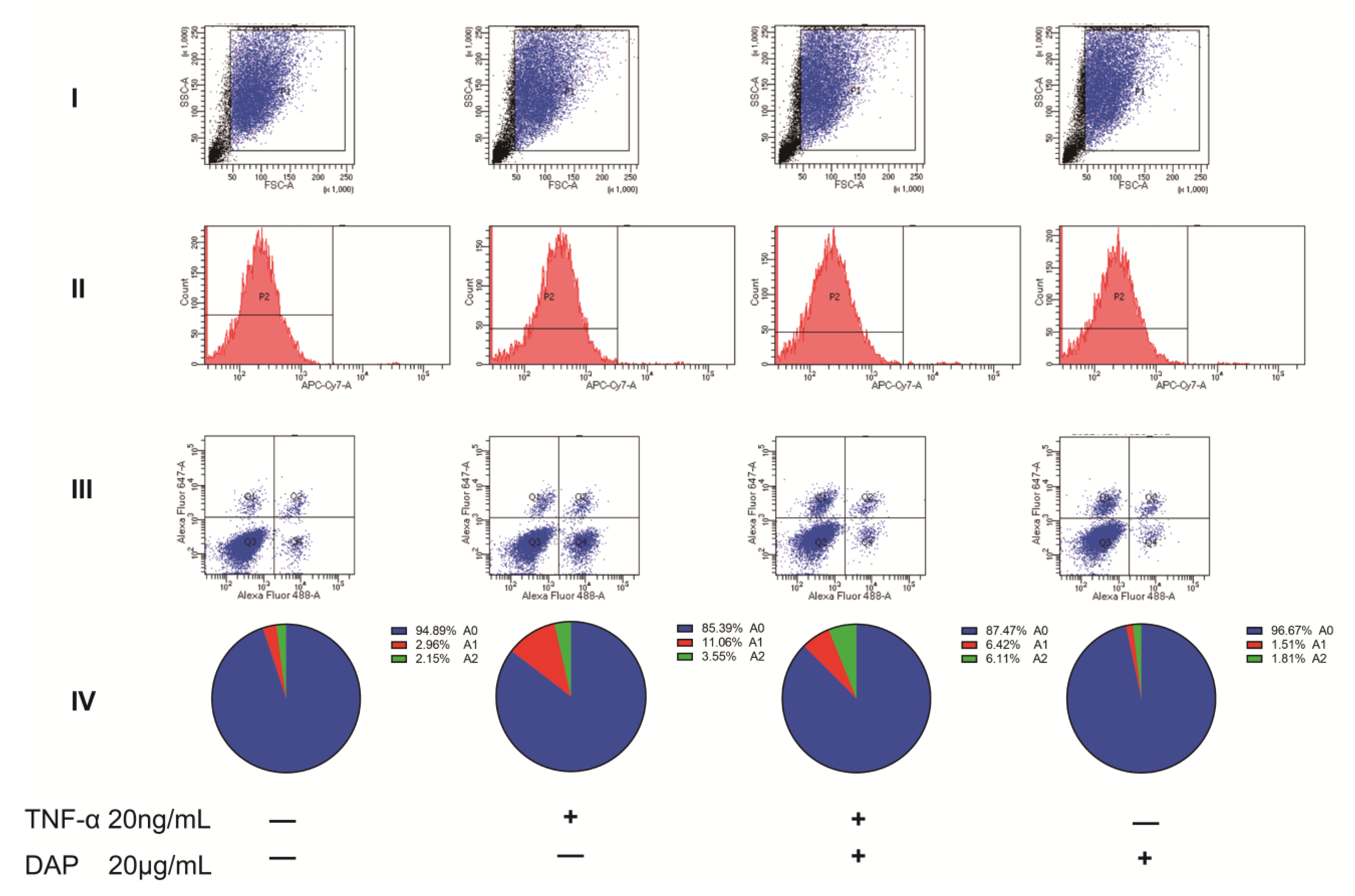
2.7. Intervention of Daphnetin on Polarization of U251 Cells
3. Discussion
4. Materials and Methods
4.1. NP Modeling
4.2. Grouping and Drug Administration
4.3. Behavioral Testing
4.4. Immunofluorescence Staining
4.5. Real-Time Fluorescence Quantitative PCR Detection of Relevant mRNA Content
4.6. Western Blot to Determine Protein Expression
4.7. Cell Culture
4.8. Cell Viability Testing
4.9. Biochemical Testing
4.10. Flow Cytometry
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alles, S.R.A.; Smith, P.A. Etiology and Pharmacology of Neuropathic Pain. Pharmacol. Rev. 2018, 70, 315–347. [Google Scholar] [CrossRef] [PubMed]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Maier, C.; Attal, N.; Binder, A.; Bouhassira, D.; Cruccu, G.; Finnerup, N.; Haanpää, M.; Hansson, P.; Hüllemann, P.; et al. Peripheral neuropathic pain: A mechanism-related organizing principle based on sensory profiles. Pain 2017, 158, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Carlton, S.M.; Du, J.; Tan, H.Y.; Nesic, O.; Hargett, G.L.; Bopp, A.C.; Yamani, A.; Lin, Q.; Willis, W.D.; Hulsebosch, C.E. Peripheral and central sensitization in remote spinal cord regions contribute to. Pain 2009, 147, 265–276. [Google Scholar] [CrossRef]
- Israel, F.C.; Richard, M.R. The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef]
- Zhi-Jun, Z.; Bao-Chun, J.; Yong-Jing, G.; Id, O. Chemokines in neuron-glial cell interaction and pathogenesis of neuropathic pain. Cell. Mol. Life Sci. 2017, 74, 3275–3291. [Google Scholar] [CrossRef]
- Karli, M.; Marzia, M. The therapeutic potential of targeting chemokine signalling in the treatment of chronic pain. J. Neurochem. 2017, 141, 520–531. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Ji, R.-R. Chemokines, neuronal-glial interactions, and central processing of neuropathic pain. Pharmacol. Ther. 2010, 126, 56–68. [Google Scholar] [CrossRef]
- Jenny, S.H.; David, R.B.; Weihua, Z.; Stanley, H.A. Microglia in ALS: The good, the bad, and the resting. J. Neuroimmune Pharmacol. 2009, 4, 389–398. [Google Scholar] [CrossRef]
- Franco, R.; Fernández-Suárez, D. Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 2015, 131, 65–86. [Google Scholar] [CrossRef]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Katarzyna, P.-B.; Natalia, K.; Anna, P.; Wioletta, M.; Ewelina, R.; Agnieszka, M.J.; Dominika, P.; Joanna, M. Parthenolide Relieves Pain and Promotes M2 Microglia/Macrophage Polarization in Rat Model of Neuropathy. Neural Plast. 2015, 676473, 1–18. [Google Scholar] [CrossRef]
- Shane, A.L.; Ben, A.B. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.U.; Minhas, P.S.; Liddelow, S.A.; Haileselassie, B.; Andreasson, K.I.; Dorn, G.W., II; Mochly-Rosen, D. Fragmented mitochondria released from microglia trigger A1 astrocytic response and propagate inflammatory neurodegeneration. Nat. Neurosci. 2019, 22, 1635–1648. [Google Scholar] [CrossRef]
- Javed, M.; Saleem, A.; Xaveria, A.; Akhtar, M.F. Daphnetin: A bioactive natural coumarin with diverse therapeutic potentials. Front. Pharmacol. 2022, 13, 1–18. [Google Scholar] [CrossRef]
- Qin, G.; Jinjun, S.; Liuqing, D.; Lijun, J.; Huiqin, X. Therapeutic effects of daphnetin on adjuvant-induced arthritic rats. J. Ethnopharmacol. 2008, 120, 259–263. [Google Scholar] [CrossRef]
- Lina, T.; Sha, L.; Yingyuan, F.; Rongfeng, Y.; Zhiqin, Z.; Shulong, Y.; Xiaoping, Z.; Nanzhen, K. The therapeutic effects of daphnetin in collagen-induced arthritis involve its regulation of Th17 cells. Int. Immunopharmacol. 2012, 13, 417–423. [Google Scholar] [CrossRef]
- Hung, A.L.; Lim, M.; Doshi, T.L. Targeting cytokines for treatment of neuropathic pain. Scand. J. Pain 2017, 17, 287–293. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, B.C.; Cao, D.L.; Zhang, Z.J.; Zhang, X.; Ji, R.R.; Gao, Y.J. TRAF6 upregulation in spinal astrocytes maintains neuropathic pain by integrating TNF-alpha and IL-1beta signaling. Pain 2014, 155, 2618–2629. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.J.; Zhang, L.; Samad, O.A.; Suter, M.R.; Yasuhiko, K.; Xu, Z.Z.; Park, J.Y.; Lind, A.L.; Ma, Q.; Ji, R.R. JNK-induced MCP-1 production in spinal cord astrocytes contributes to central sensitization and neuropathic pain. J. Neurosci. 2009, 29, 4096–4108. [Google Scholar] [CrossRef]
- Okada, S.; Hara, M.; Kobayakawa, K.; Matsumoto, Y.; Nakashima, Y. Astrocyte reactivity and astrogliosis after spinal cord injury. Neurosci. Res. 2018, 126, 39–43. [Google Scholar] [CrossRef]
- Fan, Y.Y.; Huo, J. A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem. Int. 2021, 148, 1–11. [Google Scholar] [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q.; Nai, Y.; Cao, C. Suppression of miR-155 attenuates neuropathic pain by inducing an M1 to M2 switch in microglia. Folia Neuropathol. 2020, 58, 70–82. [Google Scholar] [CrossRef]
- Gong, X.; Chen, Y.; Fu, B.; Jiang, J.; Zhang, M. Infant nerve injury induces delayed microglial polarization to the M1 phenotype, and exercise reduces delayed neuropathic pain by modulating microglial activity. Neuroscience 2017, 349, 76–86. [Google Scholar] [CrossRef]
- Bazan, J.F.; Bacon, K.B.; Hardiman, G.; Wang, W.; Soo, K.; Rossi, D.; Greaves, D.R.; Zlotnik, A.; Schall, T.J. A new class of membrane-bound chemokine with a CX3C motif. Nature 1997, 385, 640–644. [Google Scholar] [CrossRef]
- Pawelec, P.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Impact of the CX3CL1/CX3CR1 Axis in Neurological Disorders. Cells 2020, 9, 2277. [Google Scholar] [CrossRef]
- Clark, A.K.; Yip, P.K.; Grist, J.; Gentry, C.; Staniland, A.A.; Marchand, F.; Dehvari, M.; Wotherspoon, G.; Winter, J.; Ullah, J.; et al. Inhibition of spinal microglial cathepsin S for the reversal of neuropathic pain. Proc. Natl. Acad. Sci. USA 2007, 104, 10655–10660. [Google Scholar] [CrossRef]
- Gu, N.; Peng, J.; Murugan, M.; Wang, X.; Eyo, U.B.; Sun, D.; Ren, Y.; DiCicco-Bloom, E.; Young, W.; Dong, H.; et al. Spinal Microgliosis Due to Resident Microglial Proliferation Is Required for Pain Hypersensitivity after Peripheral Nerve Injury. Cell Rep. 2016, 16, 605–614. [Google Scholar] [CrossRef] [Green Version]
- Ansel, K.M.; Ngo, V.N.; Hyman, P.L.; Luther, S.A.; Forster, R.; Sedgwick, J.D.; Browning, J.L.; Lipp, M.; Cyster, J.G. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 2000, 406, 309–314. [Google Scholar] [CrossRef]
- Jiang, B.C.; Cao, D.L.; Zhang, X.; Zhang, Z.J.; He, L.N.; Li, C.H.; Zhang, W.W.; Wu, X.B.; Berta, T.; Ji, R.R.; et al. CXCL13 drives spinal astrocyte activation and neuropathic pain via CXCR5. J. Clin. Invest. 2016, 126, 745–761. [Google Scholar] [CrossRef]
- Krumbholz, M.; Theil, D.; Cepok, S.; Hemmer, B.; Kivisakk, P.; Ransohoff, R.M.; Hofbauer, M.; Farina, C.; Derfuss, T.; Hartle, C.; et al. Chemokines in multiple sclerosis: CXCL12 and CXCL13 up-regulation is differentially linked to CNS immune cell recruitment. Brain 2006, 129 Pt 1, 200–211. [Google Scholar] [CrossRef]
- Li, L.; Wu, Y.; Wang, Y.; Wu, J.; Song, L.; Xian, W.; Yuan, S.; Pei, L.; Shang, Y. Resolvin D1 promotes the interleukin-4-induced alternative activation in BV-2 microglial cells. J. Neuroinflamm. 2014, 11, 1–11. [Google Scholar] [CrossRef]
- Zhao, W.; Xu, D.; Hong, W.; Zhang, L.; Wu, Q.; Gao, M.; Wang, J. Grossamide attenuates inflammation by balancing macrophage polarization through metabolic reprogramming of macrophages in mice. Int. Immunopharmacol. 2022, 112, 1–10. [Google Scholar] [CrossRef]
- Lu, H.; Ai, L.; Zhang, B. TNF-alpha induces AQP4 overexpression in astrocytes through the NF-kappaB pathway causing cellular edema and apoptosis. Biosci. Rep. 2022, 42, 1–13. [Google Scholar] [CrossRef]
- Lin, A.; Liu, J.; Gong, P.; Chen, Y.; Zhang, H.; Zhang, Y.; Yu, Y. Serum amyloid A inhibits astrocyte migration via activating p38 MAPK. J. Neuroinflamm. 2020, 17, 1–14. [Google Scholar] [CrossRef]
- Gao, W.; Ning, Y.; Peng, Y.; Tang, X.; Zhong, S.; Zeng, H. LncRNA NKILA relieves astrocyte inflammation and neuronal oxidative stress after cerebral ischemia/reperfusion by inhibiting the NF-kappaB pathway. Mol. Immunol. 2021, 139, 32–41. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, H.; Zhou, Q.; Li, Q.; Liu, N.; Li, Z.; Chen, C.; Deng, Y. Melatonin Ameliorates Axonal Hypomyelination of Periventricular White Matter by Transforming A1 to A2 Astrocyte via JAK2/STAT3 Pathway in Septic Neonatal Rats. J. Inflamm. Res. 2021, 14, 5919–5937. [Google Scholar] [CrossRef]
- Mika, J.; Zychowska, M.; Popiolek-Barczyk, K.; Rojewska, E.; Przewlocka, B. Importance of glial activation in neuropathic pain. Eur. J. Pharmacol. 2013, 716, 106–119. [Google Scholar] [CrossRef]
- Li, Y.Y.; Wei, X.H.; Lu, Z.H.; Chen, J.S.; Huang, Q.D.; Gong, Q.J. Src/p38 MAPK pathway in spinal microglia is involved in mechanical allodynia induced by peri-sciatic administration of recombinant rat TNF-alpha. Brain Res. Bull. 2013, 96, 54–61. [Google Scholar] [CrossRef]
- Wei, X.H.; Zang, Y.; Wu, C.Y.; Xu, J.T.; Xin, W.J.; Liu, X.G. Peri-sciatic administration of recombinant rat TNF-alpha induces mechanical allodynia via upregulation of TNF-alpha in dorsal root ganglia and in spinal dorsal horn: The role of NF-kappa B pathway. Exp. Neurol. 2007, 205, 471–484. [Google Scholar] [CrossRef]
- Yang, Y.; Sheng, Q.; Nie, Z.; Liu, L.; Zhang, W.; Chen, G.; Ye, F.; Shi, L.; Lv, Z.; Xie, J.; et al. Daphnetin inhibits spinal glial activation via Nrf2/HO-1/NF-kappaB signaling pathway and attenuates CFA-induced inflammatory pain. Int. Immunopharmacol. 2021, 98, 1–8. [Google Scholar] [CrossRef]
- Watkins, L.R.; Maier, S.F. Beyond neurons: Evidence that immune and glial cells contribute to pathological pain states. Physiol. Rev. 2002, 82, 981–1011. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Kim, D.S.; Li, K.W.; Boroujerdi, A.; Peter Yu, Y.; Zhou, C.Y.; Deng, P.; Park, J.; Zhang, X.; Lee, J.; Corpe, M.; et al. Thrombospondin-4 contributes to spinal sensitization and neuropathic pain states. J. Neurosci. 2012, 32, 8977–8987. [Google Scholar] [CrossRef]
- Zhang, Q.; Cao, D.L.; Zhang, Z.J.; Jiang, B.C.; Gao, Y.J. Chemokine CXCL13 mediates orofacial neuropathic pain via CXCR5/ERK pathway in the trigeminal ganglion of mice. J. Neuroinflamm. 2016, 13, 1–13. [Google Scholar] [CrossRef]
- Subbarayan, M.S.; Joly-Amado, A.; Bickford, P.C.; Nash, K.R. CX3CL1/CX3CR1 signaling targets for the treatment of neurodegenerative diseases. Pharmacol. Ther. 2022, 231, 1–13. [Google Scholar] [CrossRef]
- Milligan, E.D.; Zapata, V.; Chacur, M.; Schoeniger, D.; Biedenkapp, J.; O’Connor, K.A.; Verge, G.M.; Chapman, G.; Green, P.; Foster, A.C.; et al. Evidence that exogenous and endogenous fractalkine can induce spinal nociceptive facilitation in rats. Eur. J. Neurosci. 2004, 20, 2294–2302. [Google Scholar] [CrossRef]
- Zhuang, Z.Y.; Kawasaki, Y.; Tan, P.H.; Wen, Y.R.; Huang, J.; Ji, R.R. Role of the CX3CR1/p38 MAPK pathway in spinal microglia for the development of neuropathic pain following nerve injury-induced cleavage of fractalkine. Brain Behav. Immun. 2007, 21, 642–651. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.K.; Malcangio, M. Microglial signalling mechanisms: Cathepsin S and Fractalkine. Exp. Neurol. 2012, 234, 283–292. [Google Scholar] [CrossRef]
- Wu, J.; Ding, D.H.; Li, Q.Q.; Wang, X.Y.; Sun, Y.Y.; Li, L.J. Lipoxin A4 Regulates Lipopolysaccharide-Induced BV2 Microglial Activation and Differentiation via the Notch Signaling Pathway. Front. Cell. Neurosci. 2019, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.; Yoo, E.; Choi, H.I.; An, G.Y.; Chai, J.C.; Lee, Y.S.; Jung, K.H.; Chai, Y.G. The BET inhibitor attenuates the inflammatory response and cell migration in human microglial HMC3 cell line. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Kaur, A.; Singh, A.P.; Bhatti, R. Daphnetin, a natural coumarin averts reserpine-induced fibromyalgia in mice: Modulation of MAO-A. Exp. Brain Res. 2021, 239, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Wang, Y.; An, K.; Liu, Q.; Xu, L.; Zhu, C.; Deng, H.; He, Q.; Wang, T.; Xu, M.; et al. Crosstalk between NFkappaB-dependent astrocytic CXCL1 and neuron CXCR2 plays a role in descending pain facilitation. J. Neuroinflamm. 2019, 16, 1–16. [Google Scholar] [CrossRef]
- Reeve, A.J.; Patel, S.; Fox, A.; Walker, K.; Urban, L. Intrathecally administered endotoxin or cytokines produce allodynia, hyperalgesia and changes in spinal cord neuronal responses to nociceptive stimuli in the rat. Eur. J. Pain 2000, 4, 247–257. [Google Scholar] [CrossRef]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef]
- Wang, L.; Yin, C.; Liu, T.; Abdul, M.; Zhou, Y.; Cao, J.L.; Lu, C. Pellino1 regulates neuropathic pain as well as microglial activation through the regulation of MAPK/NF-kappaB signaling in the spinal cord. J. Neuroinflamm. 2020, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward | Reverse |
|---|---|---|
| Gfap | GAGTGGTATCGGTCCAAGTT | CTCAAGGTCGCAGGTCAA |
| Iba-1 | ATGAGCCAGAGCAAGGATT | GCATTCGCTTCAAGGACA |
| C-fos | TGTGACCTCCCTGGACTTG | CACTGGGCCTAGATGATGC |
| Il-1β | GGCAACTGTCCCTGAACT | TCCACAGCCACAATGAGT |
| Il-6 | TCTCTCCGCAAGAGACTTCC | CCGGACTTGTGAAGTAGGGA |
| Tnf-α | ACTCCCAGAAAAGCAAGCAA | CGAGCAGGAATGAGAAGAGG |
| Cd86 | ATAGCACTGCATACCTGCCG | AGTCTTTCTGCTGGGTCTGCC |
| Cd80 | AGGAGGATCGTACGTGGTGA | GGAGGGTCTTCTGGGGGTTT |
| Cd11b | CTGCCATTACCTCCAACGGT | CTCCCAGGCACCGAAATTCT |
| Cd206 | AGTCTGCCTTAACCTGGCAC | AGGCACATCACTTTCCGAGG |
| C3 | TGTGGCAGACCCCTATGAGA | TCATTCCTTCTGGCACGACC |
| S100a10 | GAAAGGGAGTTCCCTGGGTT | CCCACTTTTCCATCTCGGCA |
| Cx3cl1 | TGCACAGCCCAGATCATTCA | CTGCGCTCTCAGATGTAGGAAA |
| Cx3cr1 | GGAGCAGGCAGGACAGCAT | CCCTCTCCCTCGCTTGTGTA |
| Cxcl13 | CCTTGCAAAAATCAGGCTTCC | CACCTTAGGCTGGTAATGCGTC |
| Cxcr5 | CTATTTGCCTTGCCAGAACTCC | CACGAGCATCGGTAGCAGGA |
| β-actin | CTTCCTGGGCATGGAGTCCT | GGA GCA ATGATC TTG ATC TT |
| Hs-il-1β | GGGATAACGAGGCTTATGTGC | AGGTGGAGAGCTTTCAGTTCA |
| Hs-il-6 | GACCCAACCACAAATGCCAG | GAGTTGTCATGTCCTGCAGC |
| Hs-tnf-α | TGAGCACTGAAAGCATGATCC | GGAGAAGAGGCTGAGGAACA |
| Hs-β-actin | CTCTTCCAGCCTTCCTTCCT | AGCACTGTGTTGGCGTACAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Liang, W.; Zhang, M.; Cui, S.; Huang, X.; Ou, W.; Huang, R.; Gao, J.; Jia, Z.; Zhang, S. Daphnetin Improves Neuropathic Pain by Inhibiting the Expression of Chemokines and Inflammatory Factors in the Spinal Cord and Interfering with Glial Cell Polarization. Pharmaceuticals 2023, 16, 243. https://doi.org/10.3390/ph16020243
Zhang T, Liang W, Zhang M, Cui S, Huang X, Ou W, Huang R, Gao J, Jia Z, Zhang S. Daphnetin Improves Neuropathic Pain by Inhibiting the Expression of Chemokines and Inflammatory Factors in the Spinal Cord and Interfering with Glial Cell Polarization. Pharmaceuticals. 2023; 16(2):243. https://doi.org/10.3390/ph16020243
Chicago/Turabian StyleZhang, Tianrui, Wulin Liang, Mingqian Zhang, Shuang Cui, Xiyan Huang, Wenjing Ou, Rikang Huang, Jiahui Gao, Zhanhong Jia, and Shuofeng Zhang. 2023. "Daphnetin Improves Neuropathic Pain by Inhibiting the Expression of Chemokines and Inflammatory Factors in the Spinal Cord and Interfering with Glial Cell Polarization" Pharmaceuticals 16, no. 2: 243. https://doi.org/10.3390/ph16020243
APA StyleZhang, T., Liang, W., Zhang, M., Cui, S., Huang, X., Ou, W., Huang, R., Gao, J., Jia, Z., & Zhang, S. (2023). Daphnetin Improves Neuropathic Pain by Inhibiting the Expression of Chemokines and Inflammatory Factors in the Spinal Cord and Interfering with Glial Cell Polarization. Pharmaceuticals, 16(2), 243. https://doi.org/10.3390/ph16020243