



Underlying Mechanisms of Bergenia spp. to Treat Hepatocellular Carcinoma Using an Integrated Network Pharmacology and Molecular Docking Approach




Abstract
:1. Introduction
2. Results
2.1. Active Compounds of Bergenia spp.
2.2. Target Prediction for Bergenia spp.
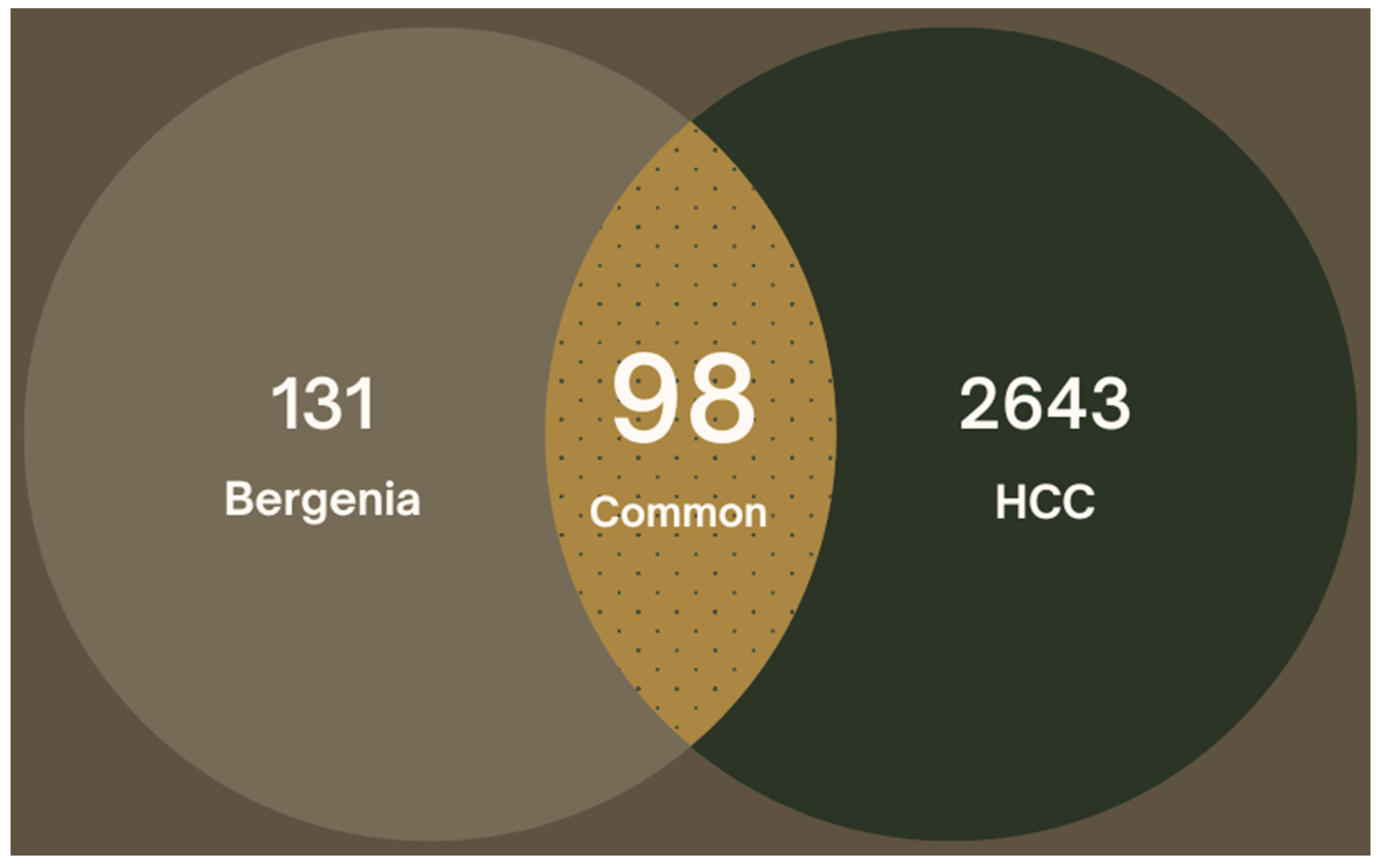
2.3. Target Prediction for Hepatocellular Carcinoma
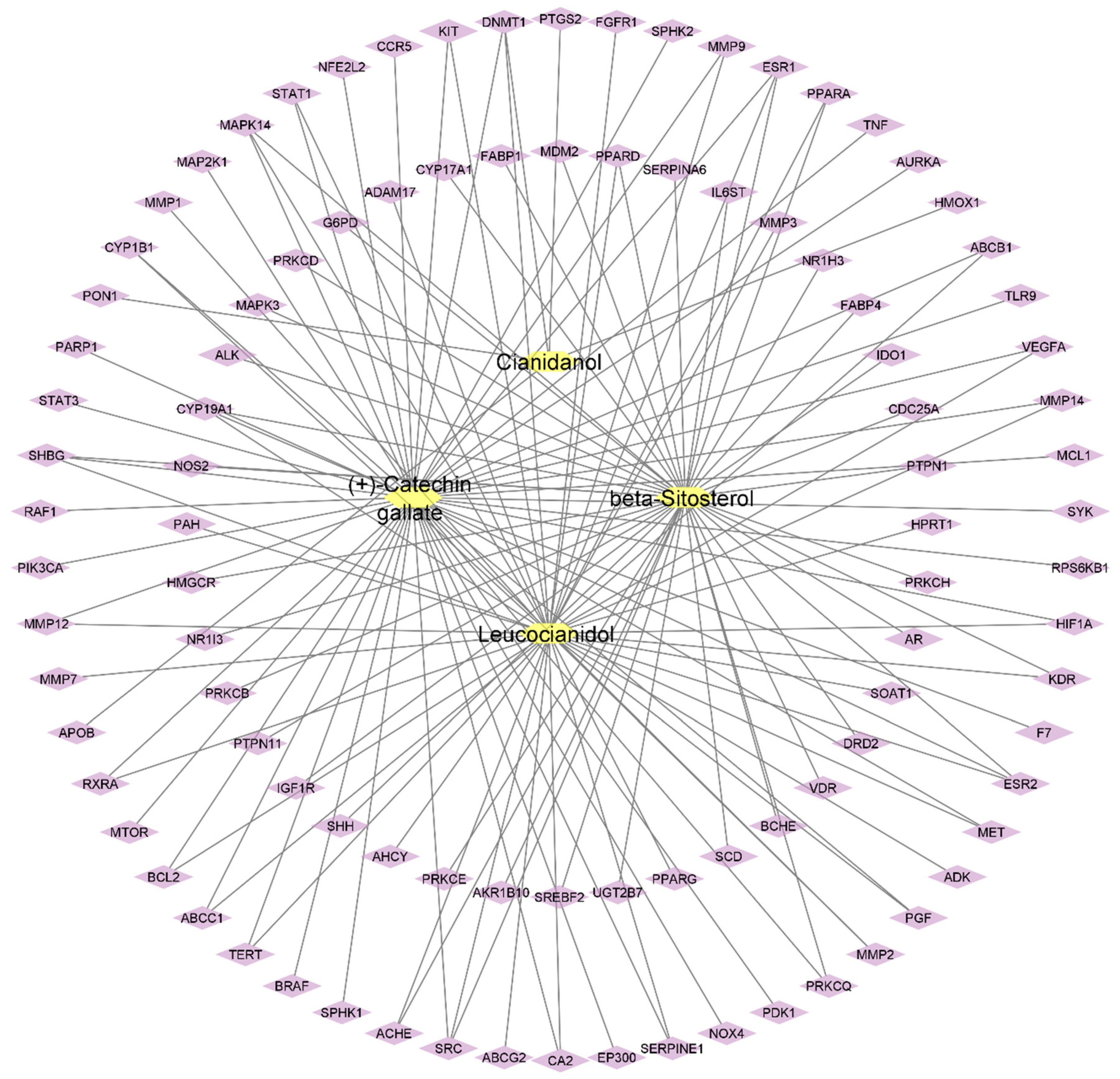
2.4. Compound-Target Network
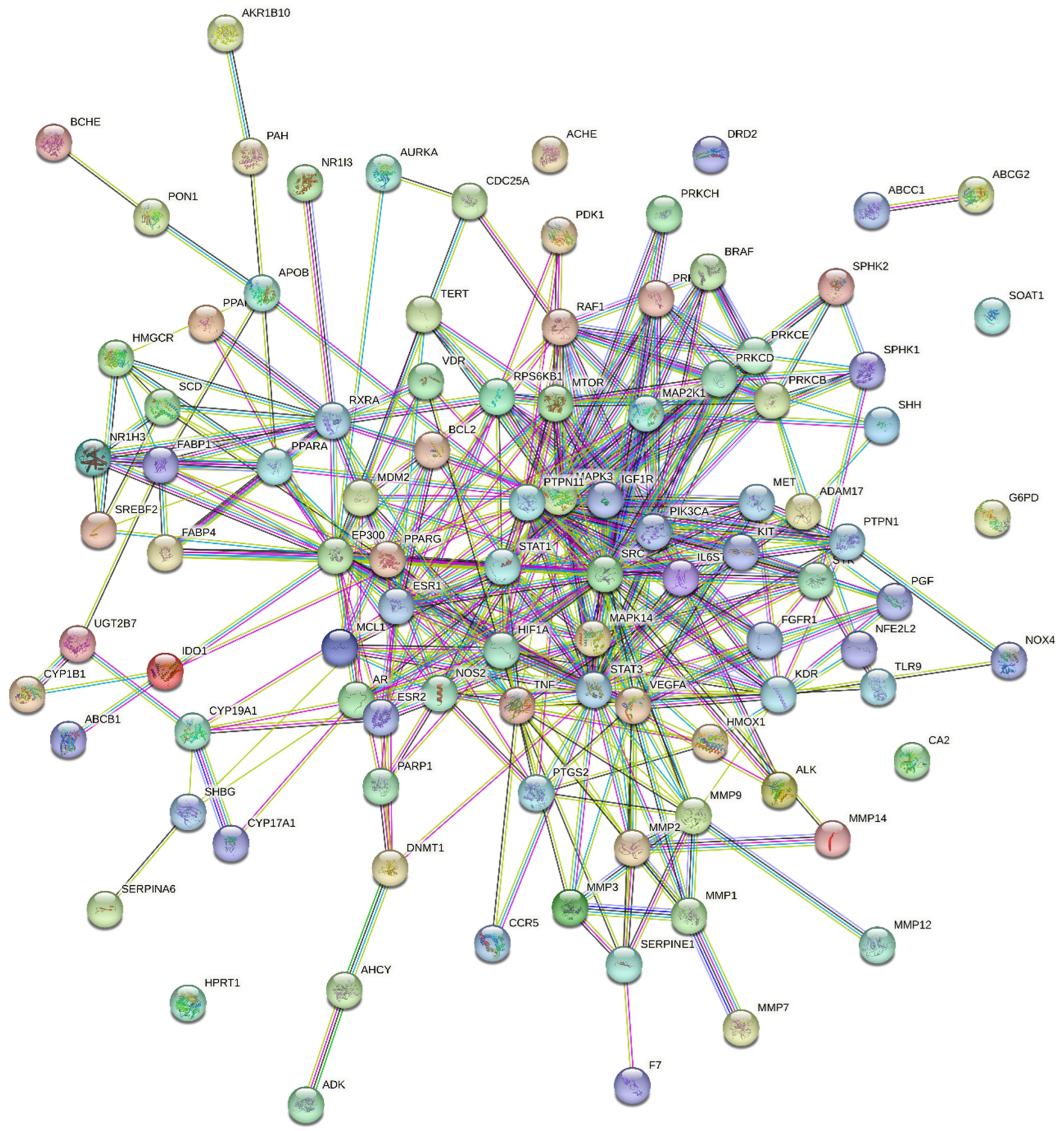
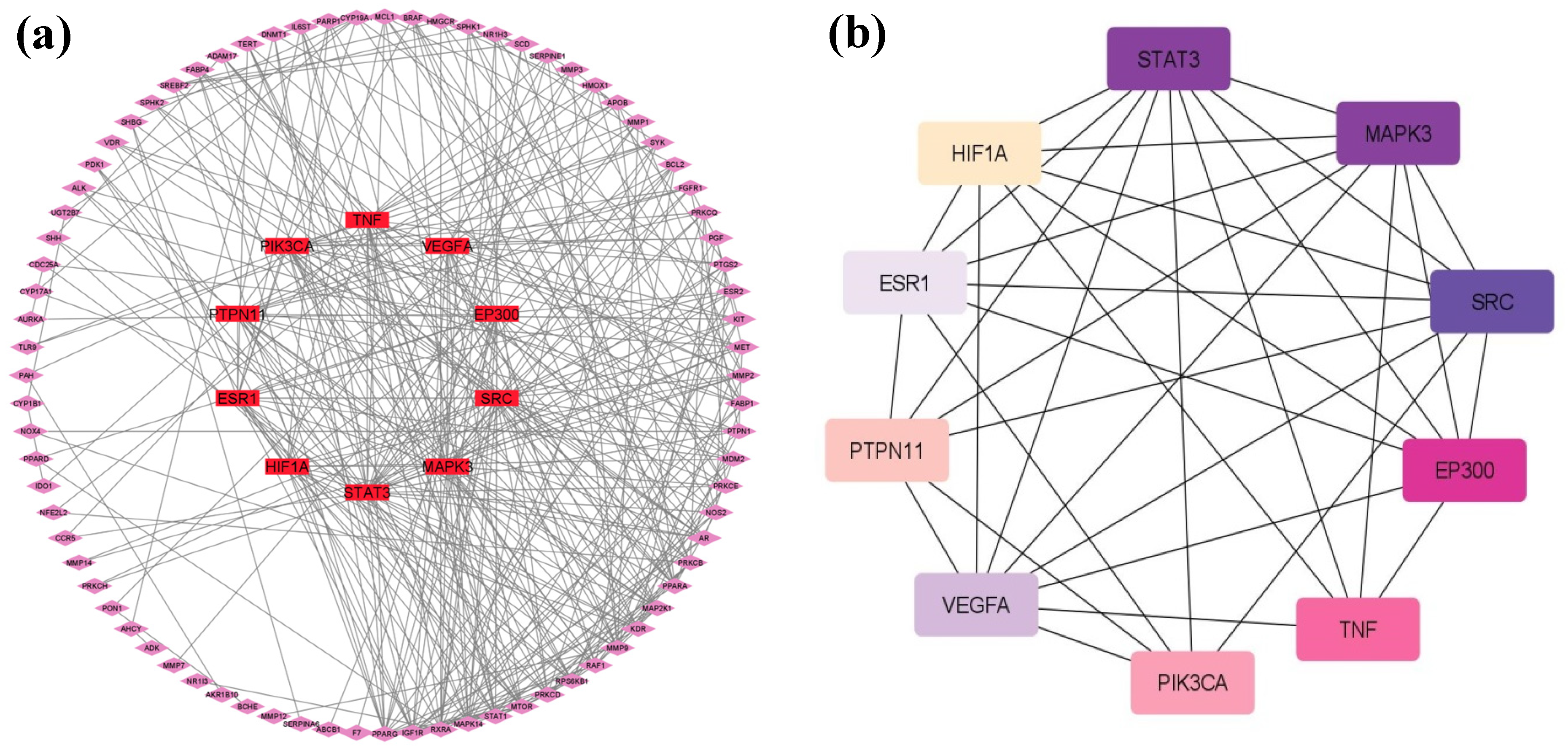
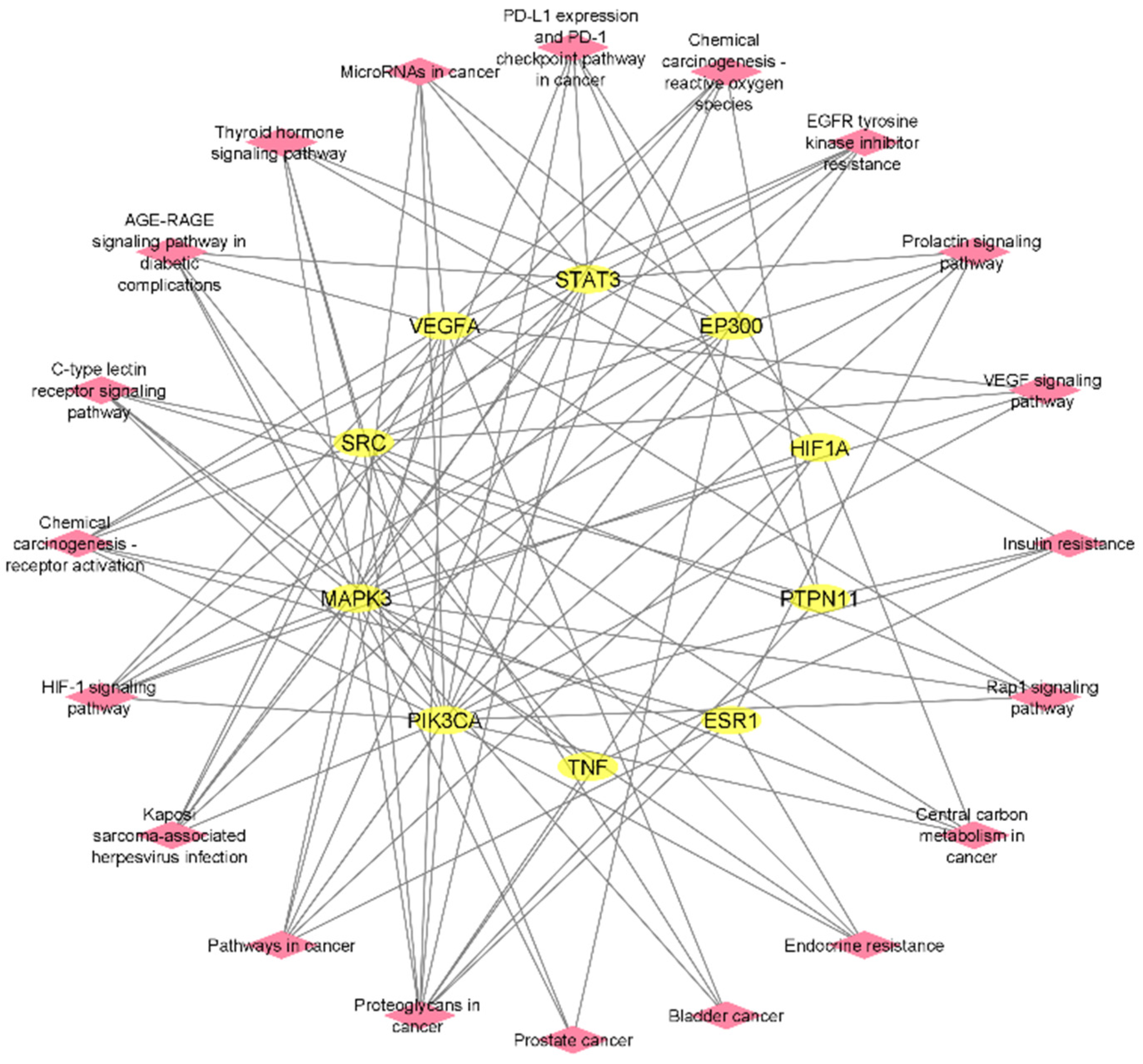
2.5. Protein–Protein Interactions (PPIs)
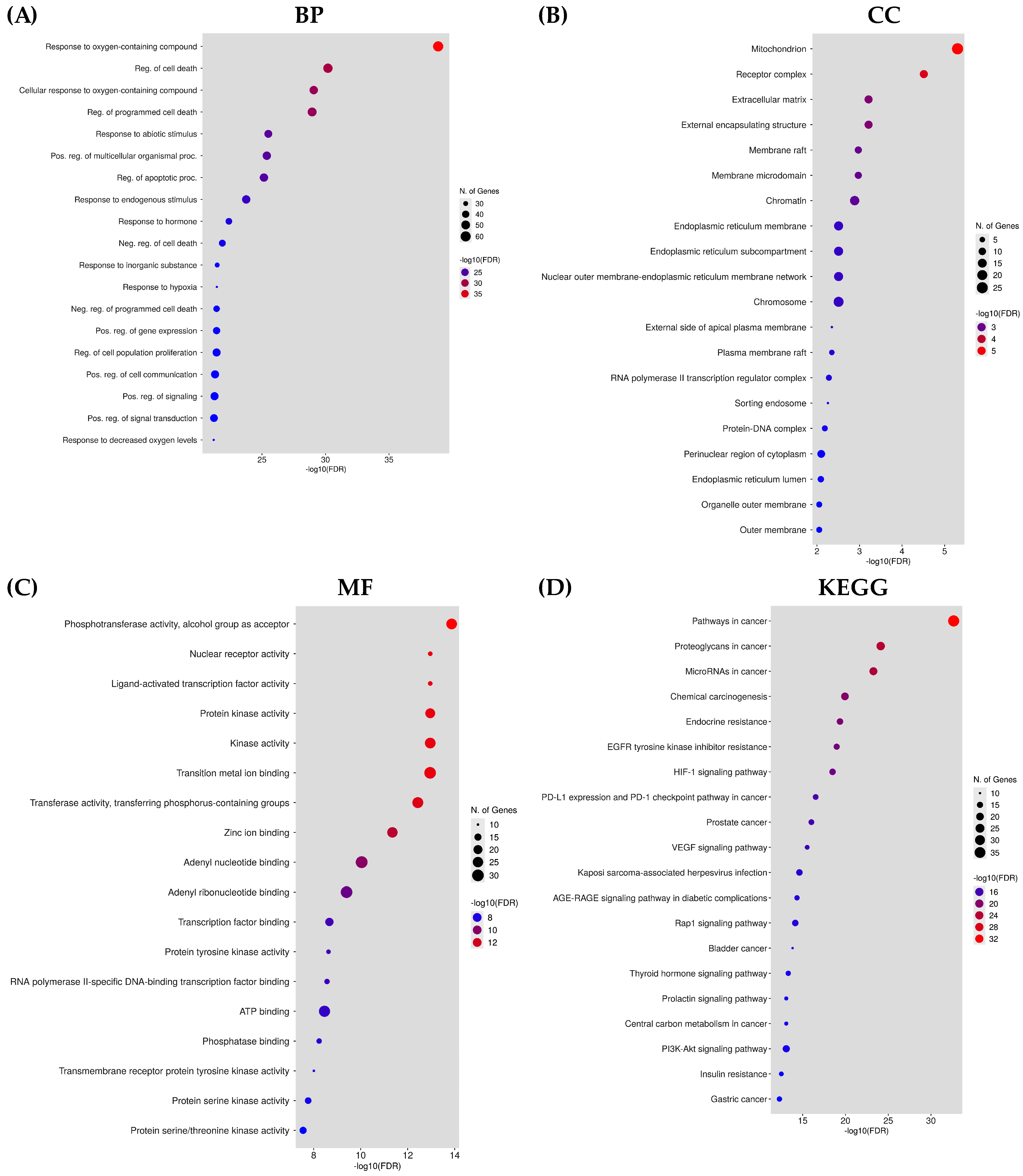
2.6. Analysis of Gene Enrichment
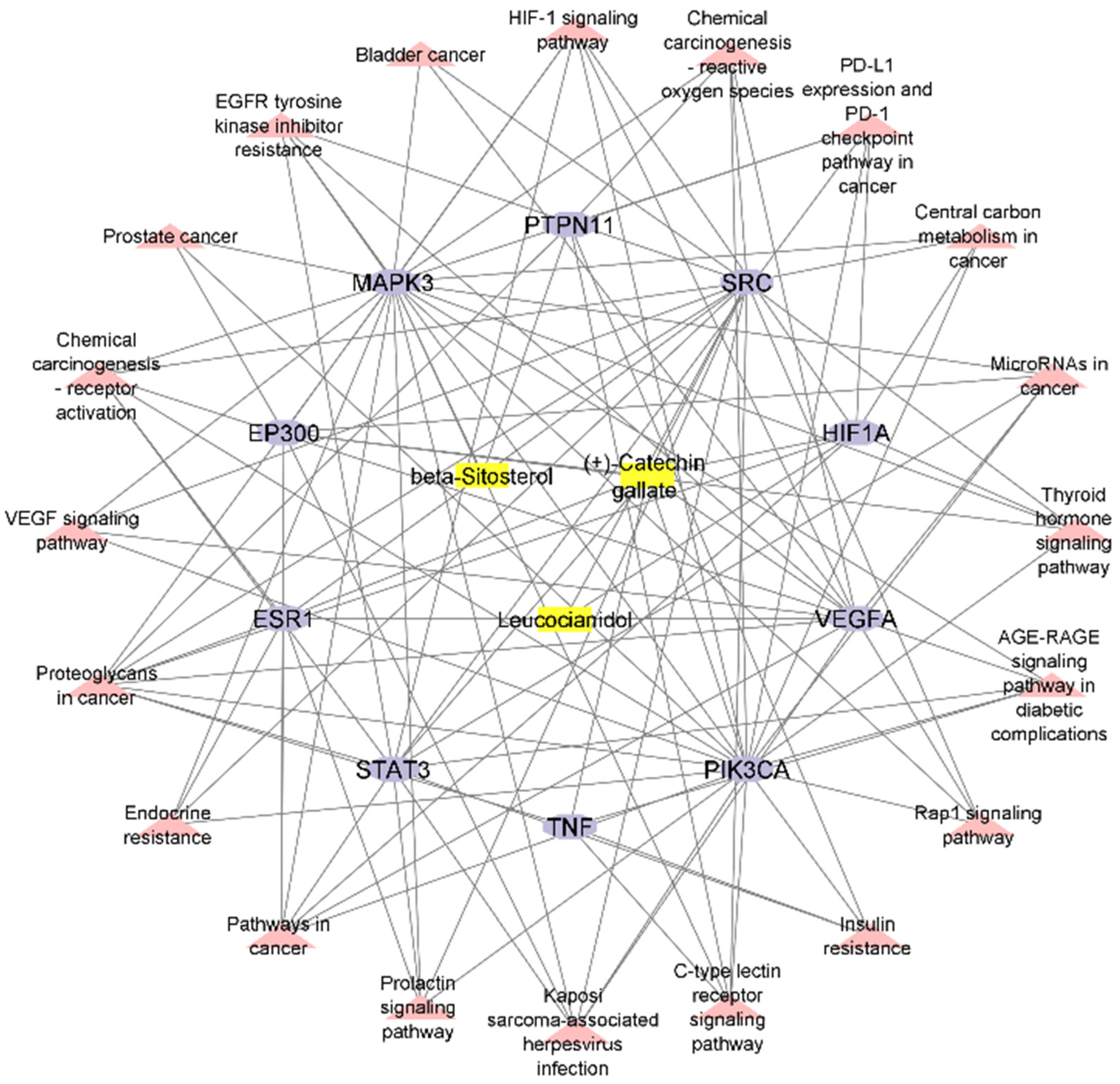
2.7. Construction of Compound–Target–Pathway Network
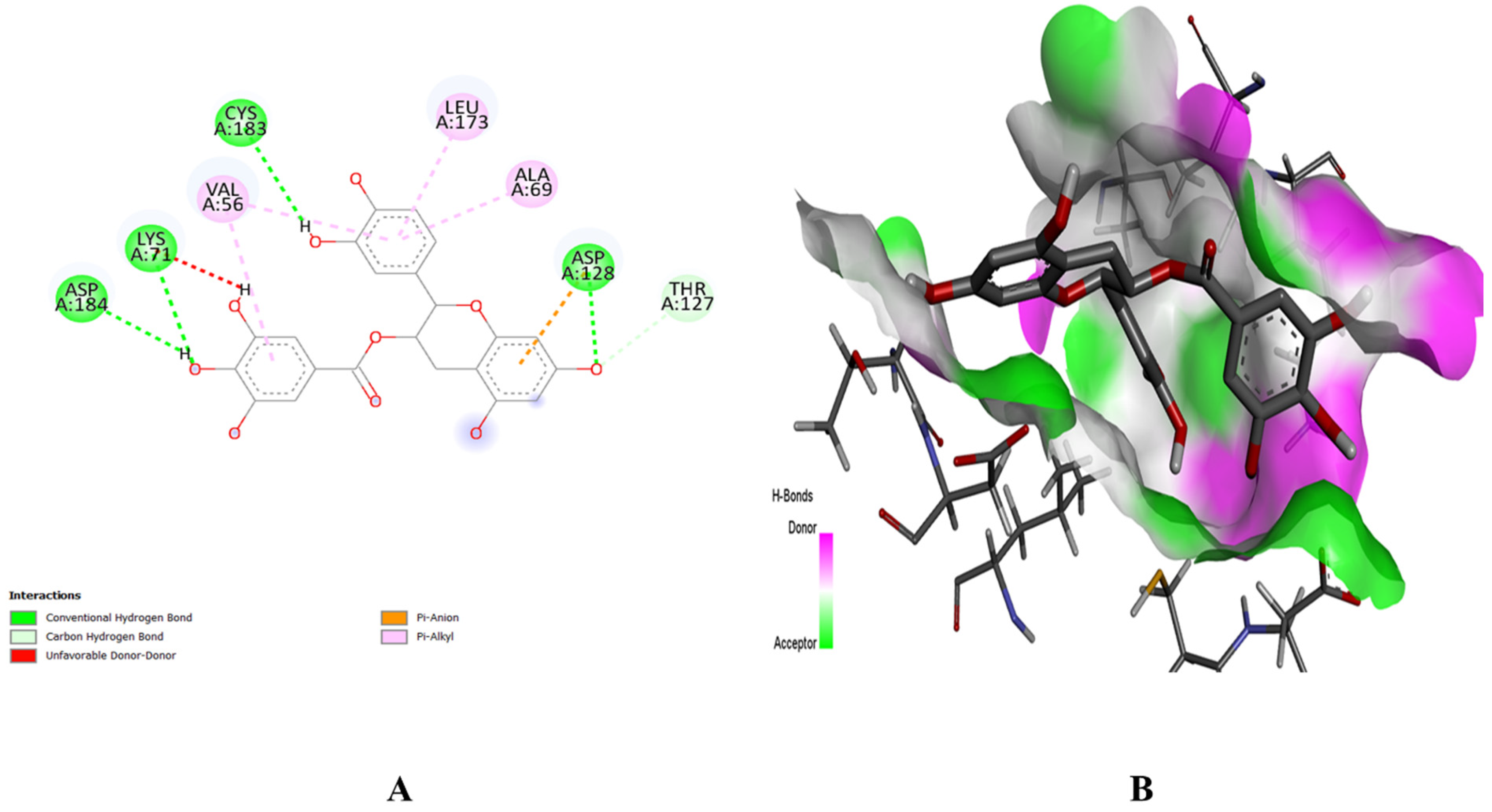
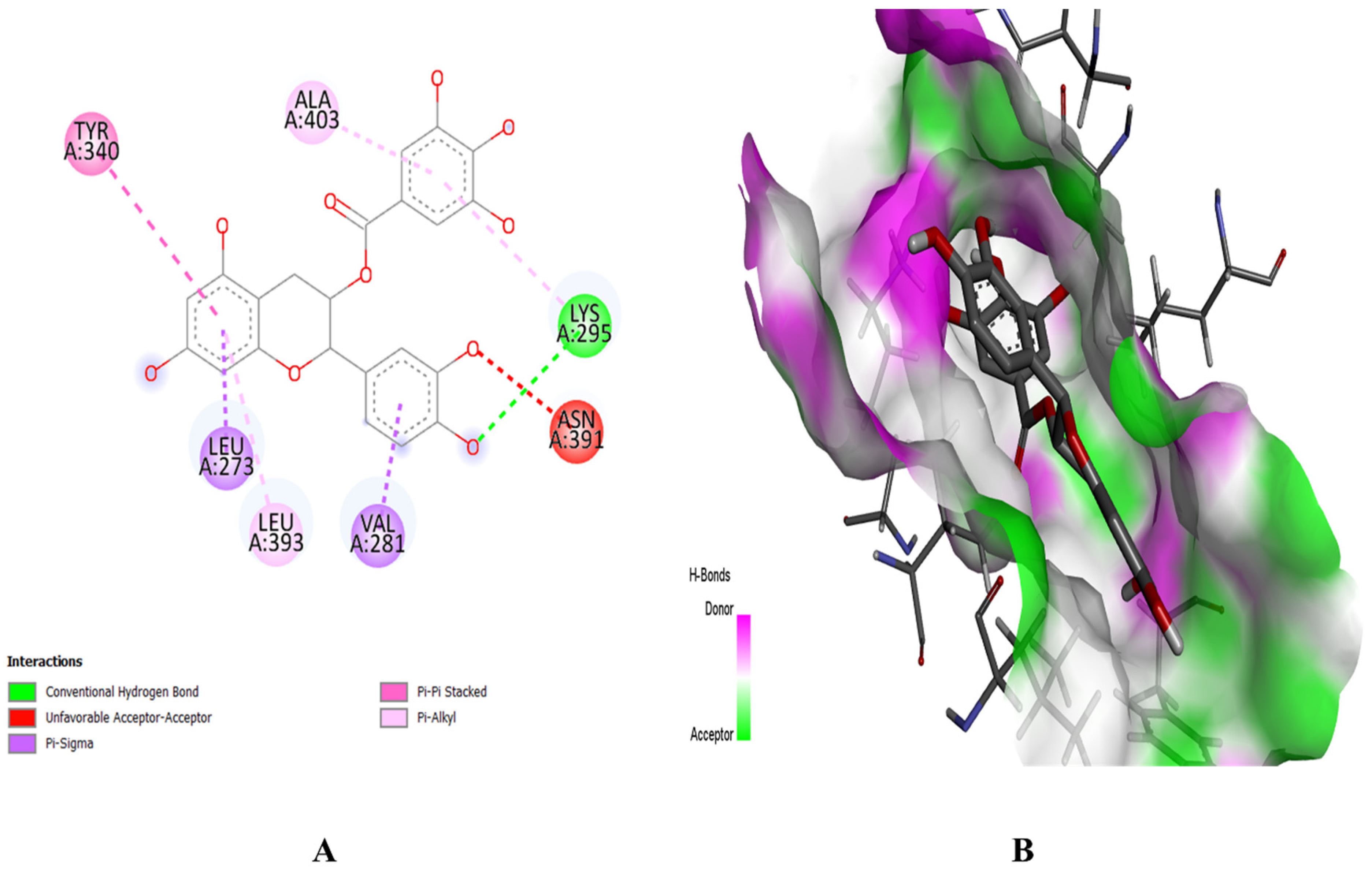
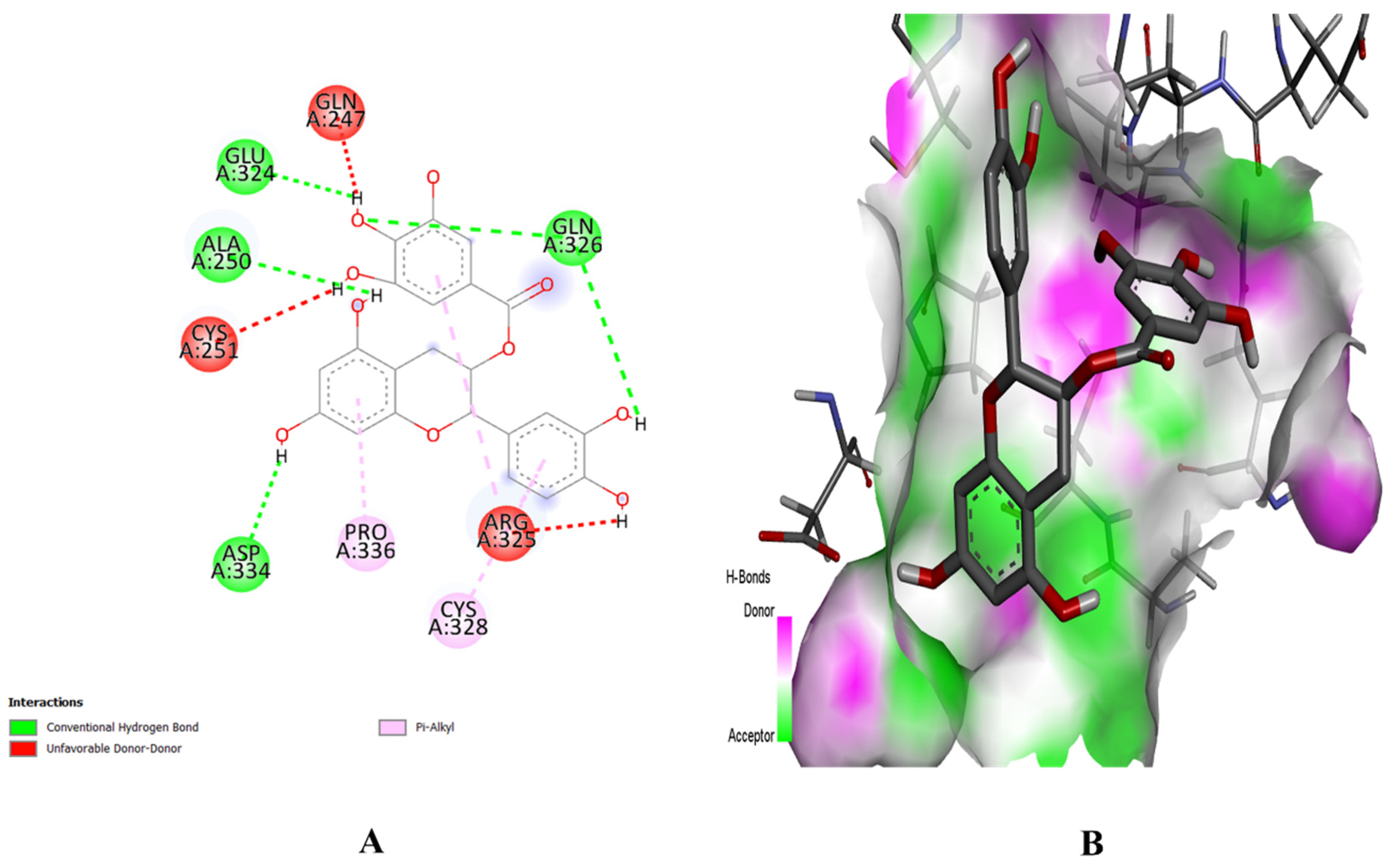
2.8. Molecular Docking Study
2.9. ADMET Profiling
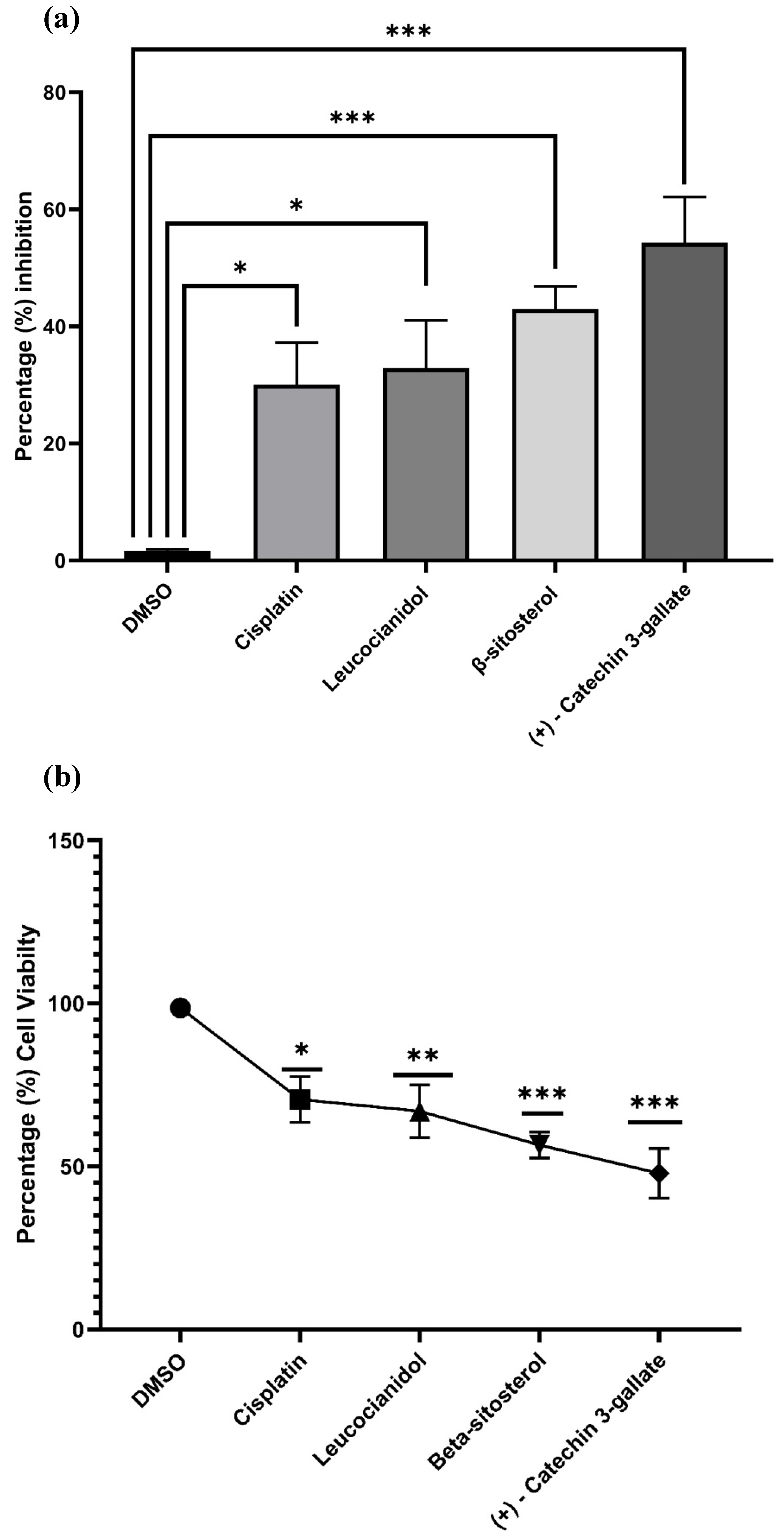
2.10. Cytotoxic Potential of the Best Selected Phytochemicals in HepG2 Cells
3. Discussion
4. Materials and Methods
4.1. Active Compounds and Targets Prediction
4.2. Drug Target Profile for Bergenia spp.
4.3. HCC-Related Target Screening
4.4. Compound–Target Network
4.5. Protein–Protein Interaction Network
4.6. Gene Ontology and KEGG Enrichment Analysis
4.7. Compound–Target–Pathway Network
4.8. Molecular Docking
4.9. ADMET Profiling
4.10. Experimental Study
4.10.1. Hep-G2 Cell Culture
4.10.2. MTT Cytotoxicity Assay
4.10.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Singal, A.G.; Lampertico, P.; Nahon, P. Epidemiology and surveillance for hepatocellular carcinoma: New trends. J. Hepatol. 2020, 72, 250–261. [Google Scholar]
- Sakurai, T.; Kudo, M. Molecular link between liver fibrosis and hepatocellular carcinoma. Liver Cancer 2013, 2, 365. [Google Scholar] [PubMed]
- Sangro, B.; Sarobe, P.; Hervás-Stubbs, S.; Melero, I. Advances in immunotherapy for hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 525–543. [Google Scholar] [PubMed]
- Sayiner, M.; Golabi, P.; Younossi, Z.M. Disease burden of hepatocellular carcinoma: A global perspective. Dig. Dis. Sci. 2019, 64, 910–917. [Google Scholar]
- Zhang, H.; Zhang, W.; Jiang, L.; Chen, Y. Recent advances in systemic therapy for hepatocellular carcinoma. Biomark. Res. 2022, 10, 1–21. [Google Scholar]
- Mustafa, G.; Younas, S.; Mahrosh, H.S.; Albeshr, M.F.; Bhat, E.A. Molecular Docking and Simulation-Binding Analysis of Plant Phytochemicals with the Hepatocellular Carcinoma Targets Epidermal Growth Factor Receptor and Caspase-9. Molecules 2023, 28, 3583. [Google Scholar]
- Khalaf, A.M.; Fuentes, D.; Morshid, A.I.; Burke, M.R.; Kaseb, A.O.; Hassan, M.; Hazle, J.D.; Elsayes, K.M. Role of Wnt/β-catenin signaling in hepatocellular carcinoma, pathogenesis, and clinical significance. J. Hepatocell. Carcinoma 2018, 5, 61–73. [Google Scholar]
- Llovet, J.M.; Castet, F.; Heikenwalder, M.; Maini, M.K.; Mazzaferro, V.; Pinato, D.J.; Pikarsky, E.; Zhu, A.X.; Finn, R.S. Immunotherapies for hepatocellular carcinoma. Nat. Rev. Clin. Oncol. 2022, 19, 151–172. [Google Scholar]
- Dutt, R.; Garg, V.; Khatri, N.; Madan, A.K. Phytochemicals in anticancer drug development. Anti-Cancer Agents Med. Chem. 2019, 19, 172–183. [Google Scholar]
- Koul, B.; Kumar, A.; Yadav, D.; Jin, J.-O. Bergenia genus: Traditional uses, phytochemistry and pharmacology. Molecules 2020, 25, 5555. [Google Scholar]
- Ali, I.; Bibi, S.; Hussain, H.; Bano, F.; Ali, S.; Khan, S.W.; Ahmad, V.U.; Al-Harrasi, A. Biological activities of Suaeda heterophylla and Bergenia stracheyi. Asian Pac. J. Trop. Dis. 2014, 4, S885–S889. [Google Scholar]
- Wang, J.; Shi, J.; Jia, N.; Sun, Q. Network pharmacology analysis reveals neuroprotection of Gynostemma pentaphyllum (Thunb.) Makino in Alzheimer’disease. BMC Complement. Med. Ther. 2022, 22, 57. [Google Scholar] [CrossRef] [PubMed]
- Sharif, S.; Atta, A.; Huma, T.; Shah, A.A.; Afzal, G.; Rashid, S.; Shahid, M.; Mustafa, G. Anticancer, antithrombotic, antityrosinase, and anti-α-glucosidase activities of selected wild and commercial mushrooms from Pakistan. Food Sci. Nutr. 2018, 6, 2170–2176. [Google Scholar]
- Mustafa, G.; Ahmed, S.; Ahmed, N.; Jamil, A. Phytochemical and antibacterial activity of some unexplored medicinal plants of Cholistan desert. Pak. J. Bot. 2016, 48, 2057–2062. [Google Scholar]
- Mustafa, G.; Arif, R.; Atta, A.; Sharif, S.; Jamil, A. Bioactive compounds from medicinal plants and their importance in drug discovery in Pakistan. Matrix Sci. Pharma 2017, 1, 17–26. [Google Scholar]
- Ali, M.; Iqbal, R.; Safdar, M.; Murtaza, S.; Mustafa, G.; Sajjad, M.; Bukhari, S.A.; Huma, T. Antioxidant and antibacterial activities of Artemisia absinthium and Citrus paradisi extracts repress viability of aggressive liver cancer cell line. Mol. Biol. Rep. 2021, 48, 7703–7710. [Google Scholar] [CrossRef]
- Spriha, S.E.; Rahman, S. In silico evaluation of selected compounds from Bergenia ciliata (haw.) sternb against molecular targets of breast cancer. Indian J. Pharm. Educ. Res 2022, 56, S105–S114. [Google Scholar] [CrossRef]
- Faheem, M.M.; Bhagat, M.; Sharma, P.; Anand, R. Induction of p53 mediated mitochondrial apoptosis and cell cycle arrest in human breast cancer cells by plant mediated synthesis of silver nanoparticles from Bergenia ligulata (Whole plant). Int. J. Pharm. 2022, 619, 121710. [Google Scholar] [CrossRef]
- Dulta, K.; Koşarsoy Ağçeli, G.; Chauhan, P.; Jasrotia, R.; Chauhan, P. A novel approach of synthesis zinc oxide nanoparticles by bergenia ciliata rhizome extract: Antibacterial and anticancer potential. J. Inorg. Organomet. Polym. Mater. 2021, 31, 180–190. [Google Scholar]
- Noor, F.; Tahir ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.; Aljasir, M.A. Network pharmacology approach for medicinal plants: Review and assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar]
- Lee, C.; Cheung, S.T. STAT3: An emerging therapeutic target for hepatocellular carcinoma. Cancers 2019, 11, 1646. [Google Scholar] [PubMed]
- Xu, J.; Lin, H.; Wu, G.; Zhu, M.; Li, M. IL-6/STAT3 is a promising therapeutic target for hepatocellular carcinoma. Front. Oncol. 2021, 11, 760971. [Google Scholar] [PubMed]
- Moon, H.; Ro, S.W. MAPK/ERK signaling pathway in hepatocellular carcinoma. Cancers 2021, 13, 3026. [Google Scholar] [PubMed]
- Hunter, C.A.; Koc, H.; Koc, E.C. c-Src kinase impairs the expression of mitochondrial OXPHOS complexes in liver cancer. Cell. Signal. 2020, 72, 109651. [Google Scholar]
- Ren, H.; Fang, J.; Ding, X.; Chen, Q. Role and inhibition of Src signaling in the progression of liver cancer. Open Life Sci. 2016, 11, 513–518. [Google Scholar] [CrossRef]
- Pal, D.; Sur, S.; Roy, R.; Mandal, S.; Kumar Panda, C. Epigallocatechin gallate in combination with eugenol or amarogentin shows synergistic chemotherapeutic potential in cervical cancer cell line. J. Cell. Physiol. 2019, 234, 825–836. [Google Scholar]
- Sabry, D.; Abdelaleem, O.O.; El Amin Ali, A.M.; Mohammed, R.A.; Abdel-Hameed, N.D.; Hassouna, A.; Khalifa, W.A. Anti-proliferative and anti-apoptotic potential effects of epigallocatechin-3-gallate and/or metformin on hepatocellular carcinoma cells: In vitro study. Mol. Biol. Rep. 2019, 46, 2039–2047. [Google Scholar]
- Ditty, M.J.; Ezhilarasan, D. β-sitosterol induces reactive oxygen species-mediated apoptosis in human hepatocellular carcinoma cell line. Avicenna J. Phytomedicine 2021, 11, 541. [Google Scholar]
- Raj, R.K. β-Sitosterol-assisted silver nanoparticles activates Nrf2 and triggers mitochondrial apoptosis via oxidative stress in human hepatocellular cancer cell line. J. Biomed. Mater. Res. Part A 2020, 108, 1899–1908. [Google Scholar]
- Maiyoa, F.; Moodley, R.; Singh, M. Phytochemistry, cytotoxicity and apoptosis studies of β-sitosterol-3-oglucoside and β-amyrin from Prunus africana. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 105–112. [Google Scholar]
- Vivek-Ananth, R.; Mohanraj, K.; Sahoo, A.K.; Samal, A. IMPPAT 2.0: An enhanced and expanded phytochemical atlas of Indian medicinal plants. ACS Omega 2023, 8, 8827–8845. [Google Scholar] [PubMed]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar]
- Guo, W.; Huang, J.; Wang, N.; Tan, H.-Y.; Cheung, F.; Chen, F.; Feng, Y. Integrating network pharmacology and pharmacological evaluation for deciphering the action mechanism of herbal formula zuojin pill in suppressing hepatocellular carcinoma. Front. Pharmacol. 2019, 10, 1185. [Google Scholar] [PubMed]
- Tao, W.; Xu, X.; Wang, X.; Li, B.; Wang, Y.; Li, Y.; Yang, L. Network pharmacology-based prediction of the active ingredients and potential targets of Chinese herbal Radix Curcumae formula for application to cardiovascular disease. J. Ethnopharmacol. 2013, 145, 1–10. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; Von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting protein–chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar]
- Safran, M.; Rosen, N.; Twik, M.; BarShir, R.; Stein, T.I.; Dahary, D.; Fishilevich, S.; Lancet, D. The genecards suite. In Practical Guide to Life Science Databases; Springer: Singapore, 2021; pp. 27–56. [Google Scholar]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Arif, R.; Zia, M.A.; Mustafa, G. Structural and functional annotation of napin-like protein from momordica charantia to explore its medicinal importance. Biochem. Genet. 2021, 60, 415–432. [Google Scholar]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [PubMed]
- Mustafa, G.; Mahrosh, H.S.; Attique, S.A.; Arif, R.; Farah, M.A.; Al-Anazi, K.M.; Ali, S. Identification of Plant Peptides as Novel Inhibitors of Orthohepevirus A (HEV) Capsid Protein by Virtual Screening. Molecules 2023, 28, 2675. [Google Scholar] [PubMed]
- Bittrich, S.; Rose, Y.; Segura, J.; Lowe, R.; Westbrook, J.D.; Duarte, J.M.; Burley, S.K. RCSB Protein Data Bank: Improved annotation, search and visualization of membrane protein structures archived in the PDB. Bioinformatics 2022, 38, 1452–1454. [Google Scholar]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. In Chemical Biology. Methods in Molecular Biology; Hempel, J., Williams, C., Hong, C., Eds.; Humana Press: New York, NY, USA, 2015; Volume 1263, pp. 243–250. [Google Scholar]
- Sharma, S.; Sharma, A.; Gupta, U. Molecular Docking studies on the Anti-fungal activity of Allium sativum (Garlic) against Mucormycosis (black fungus) by BIOVIA discovery studio visualizer 21.1. 0.0. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar]
- Pires, D.E.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar]
- Azeem, M.; Mustafa, G.; Mahrosh, H.S. Virtual screening of phytochemicals by targeting multiple proteins of severe acute respiratory syndrome coronavirus 2: Molecular docking and molecular dynamics simulation studies. Int. J. Immunopathol. Pharmacol. 2022, 36, 03946320221142793. [Google Scholar]
- Rasul, A.; Riaz, A.; Wei, W.; Sarfraz, I.; Hassan, M.; Li, J.; Asif, F.; Adem, Ş.; Bukhari, S.A.; Asrar, M. Mangifera indica extracts as novel PKM2 inhibitors for treatment of triple negative breast cancer. BioMed Res. Int. 2021, 2021, 5514669. [Google Scholar] [PubMed]
- Zara, R.; Rasul, A.; Sultana, T.; Jabeen, F.; Selamoglu, Z. Identification of Macrolepiota procera extract as a novel G6PD inhibitor for the treatment of lung cancer. Saudi J. Biol. Sci. 2022, 29, 3372–3379. [Google Scholar] [PubMed]
- Abdel-Hamid, N.M.; EL-Gharieb, M.S.; El-Senduny, F.F.; Alnakib, N.A.-A. Effect of Doxorubicin and Cisplatin on Alpha-fetoprotein levels in Hepatocellular Carcinoma Cell lines. Alfarama J. Basic Appl. Sci. 2022, 3, 35–44. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Compound | Molecular Formula | Oral Bioavailability (>30%) | Drug Likeness (>0.18) | MW (g/mol) | PubChem ID |
|---|---|---|---|---|---|---|
| 1 | β-Sitosterol | C29H50O | 36.91 | 0.75 | 414.79 | 222284 |
| 2 | Cianidanol | C15H14O6 | 54.83 | 0.24 | 290.29 | 9064 |
| 3 | (+)-catechin gallate | C22H18O10 | 53.57 | 0.75 | 442.4 | 5276454 |
| 4 | Leucocianidol | C15H14O7 | 30.84 | 0.27 | 306.29 | 440833 |
| Sr. No. | Hub Genes | Degrees |
|---|---|---|
| 1 | STAT3 | 37 |
| 2 | MAPK3 | 34 |
| 3 | SRC | 33 |
| 4 | EP300 | 25 |
| 5 | VEGFA | 22 |
| 6 | PIK3CA | 22 |
| 7 | TNF | 22 |
| 8 | PTPN11 | 21 |
| 9 | ESR1 | 19 |
| 10 | HIF1A | 19 |
| Sr. No. | Target Protein | PDB ID | UniProt ID | Phytochemical | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|
| 1 | STAT3 | 6TLC | P40763 | (+)-Catechin 3-gallate | −8.0 |
| β-sitosterol | −7.4 | ||||
| Leucocianidol | −7.1 | ||||
| 2 | MAPK3 | 6GES | P27361 | (+)-Catechin 3-Gallate | −10.2 |
| β-sitosterol | −9.2 | ||||
| Leucocianidol | −7.4 | ||||
| 3 | SRC | 2H8H | P12931 | (+)-Catechin 3-Gallate | −8.9 |
| β-sitosterol | −8.4 | ||||
| Leucocianidol | −7.2 |
| ADMET Parameters | Phytochemicals | ||
|---|---|---|---|
| (+)-Catechin 3-Gallate | β-Sitosterol | Leucocianidol | |
| Absorption and distribution | |||
| BBB | No | No | No |
| Intestinal absorption (human) | 62.096% | 94.464% | 56.712% |
| PGS | Yes | No | Yes |
| PGI | No | No | No |
| Metabolism | |||
| CYP3A4 substrate | No | Yes | No |
| CYP2D6 substrate | No | No | No |
| CYP3A4 inhibition | No | No | No |
| CYP2C9 inhibition | No | No | No |
| CYP2C19 inhibition | No | No | No |
| CYP2D6 inhibition | No | No | No |
| CYP1A2 inhibition | No | No | No |
| Excretion | |||
| Total Clearance | −0.169 log mL/min/kg | 0.628 log mL/min/kg | −0.072 log mL/min/kg |
| Toxicity | |||
| AMES Toxicity | No | No | No |
| Hepatotoxicity | No | No | No |
| Skin Sensitization | No | No | No |
| Concentration (μg/mL) | DMSO | Catechin | β-Sitosterol | (+)-Catechin 3-Gallate |
|---|---|---|---|---|
| 1.5625 | 0 | 7 NS | 19.42 *** | 16.21 ** |
| 3.125 | 1.3 | 12 NS | 39.3 **** | 34.23 **** |
| 6.25 | 1.7 | 15 NS | 40.2 **** | 42.65 **** |
| 12.50 | 2 | 29.24 ** | 41.65 ** | 55.92 *** |
| 25 | 2 | 36.95 *** | 49.14 **** | 60.05 **** |
| 50 | 2 | 41.56 ** | 64.86 **** | 69.6 **** |
| 100 | 2 | 43.1 *** | 56.29 **** | 75.63 **** |
| 200 | 2 | 78.4 **** | 46.8 **** | 80.24 **** |
| Concentration (μg/mL) | Cisplatin | Catechin | β-Sitosterol | (+)-Catechin 3-Gallate |
| 1.5625 | 5.6 | 7 NS | 19.42 ** | 16.21 * |
| 3.125 | 10.5 | 12 NS | 39.3 **** | 34.23 **** |
| 6.25 | 13.4 | 15 NS | 40.2 *** | 42.65 *** |
| 12.50 | 27.5 | 29.24 NS | 41.65 NS | 55.92 ** |
| 25 | 32.4 | 36.95 NS | 49.14 NS | 60.05 ** |
| 50 | 39.5 | 41.56 NS | 64.86 ** | 69.6 ** |
| 100 | 46.4 | 43.1 NS | 56.29 ** | 75.63 *** |
| 200 | 65.4 | 78.4 NS | 46.8 NS | 80.24 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, S.; Mustafa, G.; Ahmed, S.; Albeshr, M.F. Underlying Mechanisms of Bergenia spp. to Treat Hepatocellular Carcinoma Using an Integrated Network Pharmacology and Molecular Docking Approach. Pharmaceuticals 2023, 16, 1239. https://doi.org/10.3390/ph16091239
Hussain S, Mustafa G, Ahmed S, Albeshr MF. Underlying Mechanisms of Bergenia spp. to Treat Hepatocellular Carcinoma Using an Integrated Network Pharmacology and Molecular Docking Approach. Pharmaceuticals. 2023; 16(9):1239. https://doi.org/10.3390/ph16091239
Chicago/Turabian StyleHussain, Shoukat, Ghulam Mustafa, Sibtain Ahmed, and Mohammed Fahad Albeshr. 2023. "Underlying Mechanisms of Bergenia spp. to Treat Hepatocellular Carcinoma Using an Integrated Network Pharmacology and Molecular Docking Approach" Pharmaceuticals 16, no. 9: 1239. https://doi.org/10.3390/ph16091239
APA StyleHussain, S., Mustafa, G., Ahmed, S., & Albeshr, M. F. (2023). Underlying Mechanisms of Bergenia spp. to Treat Hepatocellular Carcinoma Using an Integrated Network Pharmacology and Molecular Docking Approach. Pharmaceuticals, 16(9), 1239. https://doi.org/10.3390/ph16091239