Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-κB Signaling of Immortalized and Cancer Cell Lines


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Iridoids | Phenylpropanoid | Phytosterol | Organic acids |
|---|---|---|---|
| Harpagide 12.5 mg/kg | Sibirioside A 8 mg/kg | β-Sitosterol 12 mg/kg | Cinnamic acid 14 mg/kg |
| Harpagoside 20 mg/kg | Cistanoside F 7 mg/kg | Daucosterol | Ferulic acid |
| Galactopyranosylharpagoside | Angoroside C 2 mg/kg | Butane diacid | |
| Feruloyharpagide 1 mg/kg | Cistanoside D 6 mg/kg | ||
| Hydroxycinnamoylharpagide | Acetoside 12 mg/kg | ||
| Acetylharpagoside | Decaffeoylacetoside 4 mg/kg | ||
| Aucubin 3 mg/kg | Ningposide A 5 mg/kg | ||
| Geniposide | Ningposide B 1.5 mg/kg | ||
| Catapol | Ningposide C 3 mg/kg | ||
| Methylcatapol 33 mg/kg | Ningposide D 6 mg/kg | ||
| Scropolioside A | |||
| Scrophuloside B4 6 mg/kg | Triterpenoid saponins | Sugars | Terpenes |
| Iridoidlacton | Ursolic acid 10 mg/kg | Fructose | Cryptomeriol |
| Ningpogenin | Asparagines | Sucrose | |
| Ningpogoside A | Triterpenoid saponins | Glucose | |
| Ningpogoside B |
2. Experimental Section
2.1. Chemicals
2.2. Aqueous Herbal Extract
2.3. Cell Culture

2.4. XTT Proliferation Assay

2.5. Immunofluorescence
2.6. Western Blot
2.7. Detection of Apoptosis
2.8. Statistical Analyses
3. Results and Discussion
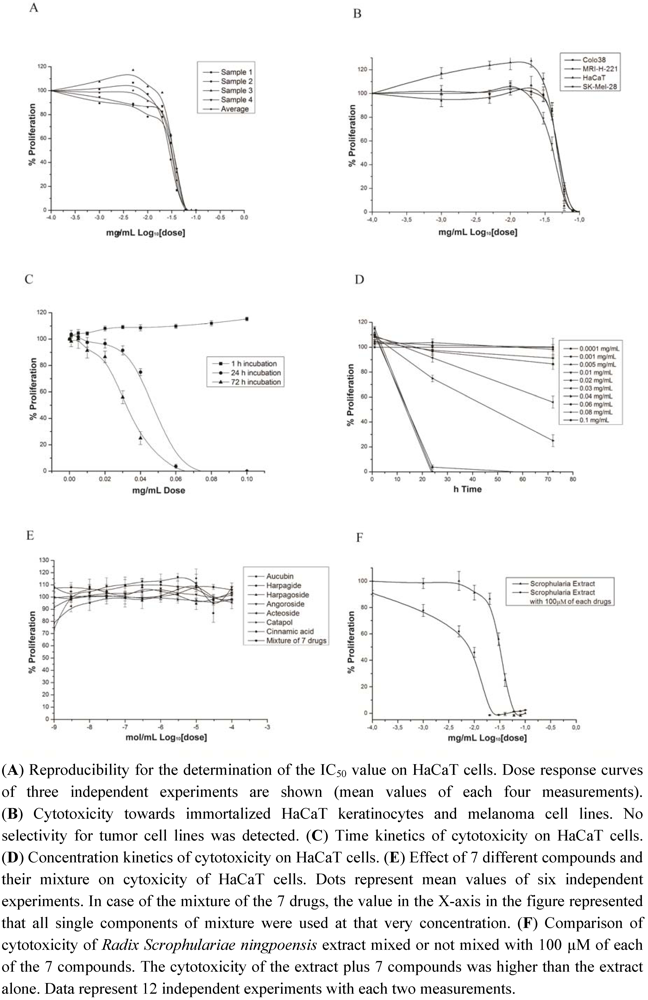
3.1. Cytotoxicity of Radix Scrophulariae ningpoensis
3.2. Enhanced Cytotoxicity of Radix Scrophularia ningpoensis


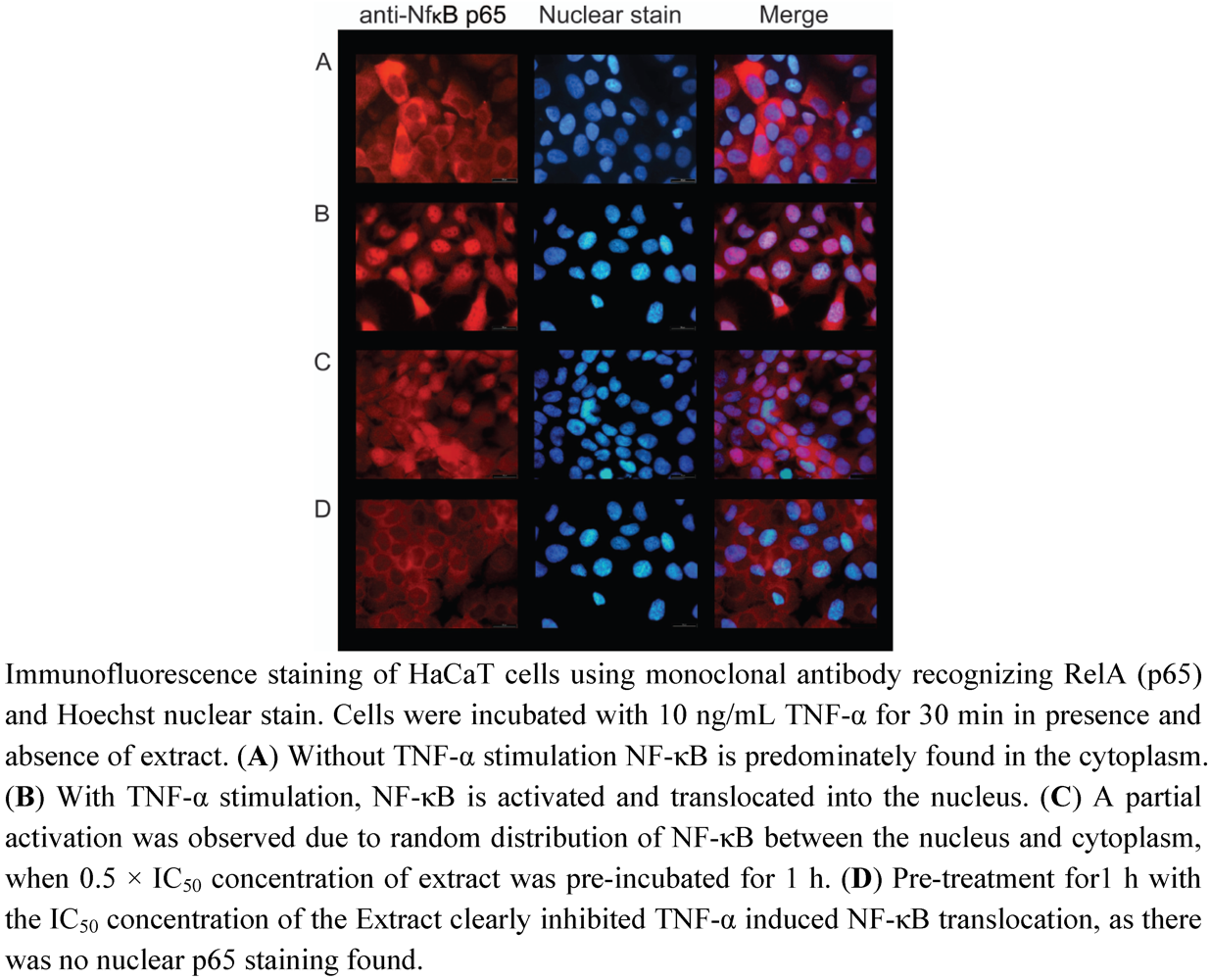
3.3. Inhibition of TNF-α Induced NF-κB Activation by Radix Scrophularia ningpoensis
3.4. Regulating effect on MAPK pathway
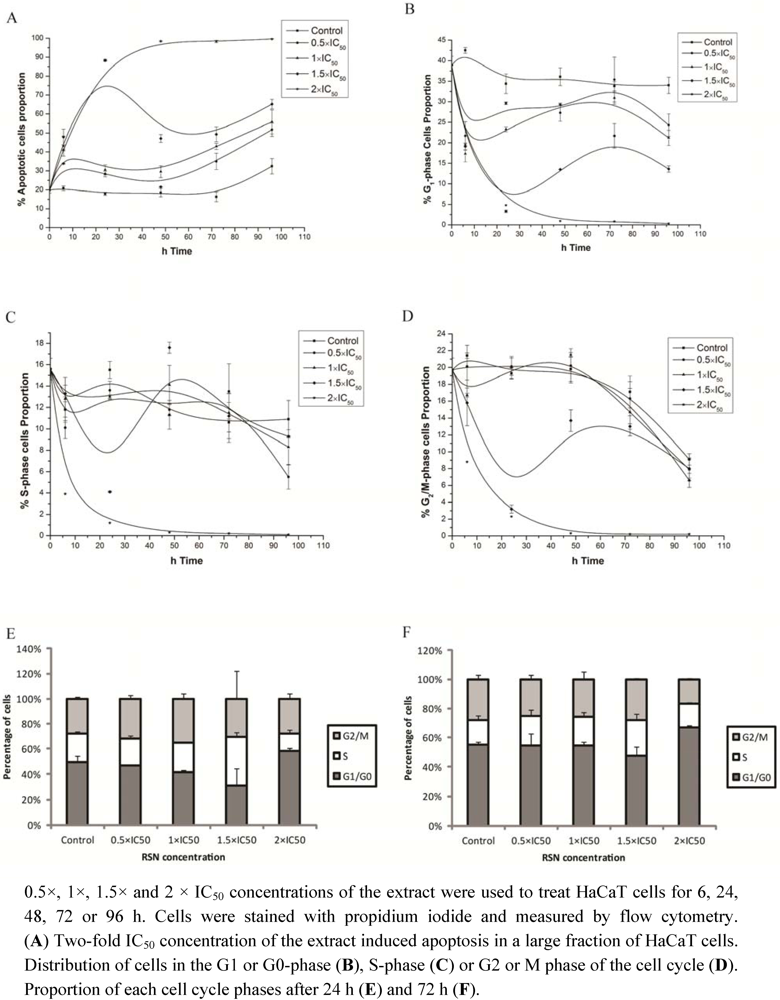
3.5. Apoptosis and Cell Growth Inhibition Induced by Radix Scrophulariae ningpoensis


3.6. Cross-Resistance of the NCI Cell Line Panel between Acetoside and Standard Drugs
| COMPARE Coefficient | Drug | Mode of action |
|---|---|---|
| 0.654 | cyclodisone | DNA alkylator |
| 0.638 | didemnin B | protein synthesis inhibitor |
| 0.614 | chlorozotocin | DNA alkylator |
| 0.604 | irinotecan | DNA topoisomerase I inhibitor |
| 0.562 | actinomycin D | RNA synthesis inhibitor |
| 0.558 | pipobroman | DNA alkylator |
| 0.554 | hepsulfam | DNA alkylator |
| 0.554 | melphalan | DNA alkylator |
| 0.552 | BCNU | DNA alkylator |
| 0.552 | chlorambucil | DNA alkylator |
| 0.545 | Yoshi-864 | DNA alkylator |
| 0.542 | methyl-CCNU | DNA alkylator |
| 0.532 | teroxirone | DNA alkylator |
| 0.508 | aclacinomycin A | DNA intercalator |
3.7. COMPARE and Cluster Analyses of Microarray-Based mRNA Hybridization
| Symbol | COMPARE Coefficient | Pattern ID | Genbank | Name | Function |
|---|---|---|---|---|---|
| Standard COMPARE: | |||||
| RAC2 | 0.723 | GC35722 | W68830 | Ras-related C3 botulinum toxin substrate 2 | Signal transduction, regulates phagocytosis of apoptotic cells |
| (Rho family, small GTP binding protein Rac2) | |||||
| PTPRC | 0.706 | GC30950 | Y00638 | Protein tyrosine phosphatase, receptor type, C | Signal transduction, T-cell activation |
| CDK6 | 0.697 | GC168940 | AW194766 | Cyclin-dependent kinase 6 | Cell cycle control |
| GPR68 | 0.695 | GC74908 | AI805006 | G protein-coupled receptor 68 | Signal transducer, metastasis suppressor in prostate cancer |
| KIRREL2 | 0.694 | GC85039 | AW025274 | Kin of IRRE like 2 (Drosophila) | Cell adhesion |
| FMNL1 | 0.688 | GC27500 | AJ008112 | Formin-like 1 | Control of cell motility and survival of macrophages |
| SOCS1 | 0.682 | GC53793 | AB000734 | Suppressor of cytokine signaling 1 | Signal transduction, regulator of IL6- and LIF-signaling |
| RGS14 | 0.679 | GC152154 | AF037194 | Regulator of G-protein signaling 14 | Signal transduction |
| PTPN22 | 0.678 | GC27207 | AF001846 | Protein tyrosine phosphatase, non-receptor | Signal transduction; negative regulator of T cell receptor signaling |
| type 22 (lymphoid) | |||||
| CD53 | 0.672 | GC28789 | M37033 | CD53 molecule | Growth regulation in hematopoietic cells |
| Reverse COMPARE: | |||||
| MSI1 | −0.609 | GC12913 | H42504 | Musashi homolog 1 (Drosophila) RNA | Regulates expression of target mRNAs at the translation level; |
| regulates proliferation and maintenance of CNS stem cells | |||||
| MAPK8 | −0.595 | GC89100 | L26318 | Mitogen-activated protein kinase 8 | Signal transduction; activated by stress and inflammatory signals |
| TSPAN6 | −0.578 | GC15930 | W68001 | Tetraspanin 6 | Signal transduction |
| ALDH7A1 | −0.552 | GC16889 | AA024918 | Aldehyde dehydrogenase 7 family, member A1 | Oxidoreductase; protects cells from oxidative stress |
| PTK2 | −0.551 | GC18530 | AA031671 | PTK2 protein tyrosine kinase 2 | Signal transduction: involved in cell motility, proliferation and apoptosis |
| DLG2 | −0.549 | GC10718 | R41930 | Discs, large homolog 2 (Drosophila) | Regulates surface expression of NMDA receptors |
| MPEG1 | −0.534 | GC18080 | AA004905 | Macrophage expressed 1 | unknown |
| KIF1B | −0.534 | GC17932 | AA002163 | Kinesin family member 1B | Motor for anterograde transport of mitochondria |
| ATPAF1 | −0.534 | GC15393 | N79086 | ATP synthase mitochondrial F1 complex | Role for the assembly of the mitochondrial F1-F0 complex |
| assembly factor 1 | |||||
| ALDH1L2 | −0.53 | GC92376 | N72255 | Aldehyde dehydrogenase 1 family, member L2 | Oxidoreductase |
4. Conclusion
Acknowledgments
Conflict of Interest
References
- Pharmacopoeia Commission of the People’s Republic of China. In Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2005.
- Miyazawa, M.; Okuno, Y.; Nakamura, S.I.; Kameokam, H. Suppression of SOS-inducing activity of chemical mutagens by cinnamic acid derivatives from Scrophularia ningpoensis in the Salmonella typhymurium TA1535/pSK1002 umu test. J. Agric. Food Chem. 1998, 46, 904–910. [Google Scholar] [CrossRef]
- Greten, J. Kursbuch Traditionelle Chinesische Medizin. Thieme Verlag: Stuttgart, New York, NY, USA, 2003. [Google Scholar]
- Yu, J.; Zhang, Z.; Yang, C. Study on the fingerprints of Radix Scrophulariae by HPLC/UV. Prog. Pharm. Sci. 2005, 29, 4. [Google Scholar]
- Hu, Y.; Huang, Z. Advances in research of chemical constituents and pharmacological effects of Scrophularia ningpoensis Hemsl. J. Zhejiang Univ. Trad. Chin. Med. 2008, 32, 268–270. [Google Scholar]
- Sun, K.; Jiang, H. Research about the hepatocytes protective function of phenylpropanoid from Scrophularia ningpoensis Hemsl. J. Pharm. Pract. 2002, 4, 234–236. [Google Scholar]
- Niu, Z.R.; Wang, R.F.; Shang, M.Y.; Cai, S.Q. A new iridoid glycoside from Scrophularia ningpoensis. Nat. Prod. Res. 2009, 23, 1181–1188. [Google Scholar] [CrossRef]
- Li, M.Y.; Jiang, S.H.; Gao, W.Y.; Zhu, D.Y. Phenylpropanoid glycosides from Scrophularia ningpoensis. Phytochemistry 2000, 54, 923–925. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Fontaine, J.; Malonne, H.; Claeys, M.; Luhmer, M.; Duez, P. A sugar ester and an iridoid glycoside from Scrophularia ningpoensis. Phytochemistry 2005, 66, 1186–1191. [Google Scholar]
- Li, Y.M.; Han, Z.H.; Jiang, S.H.; Jiang, Y.; Yao, S.D.; Zhu, D.Y. Fast repairing of oxidized OH radical adducts of dAMP and dGMP by phenylpropanoid glycosides from Scrophularia ningpoensis Hemsl. Acta Pharmacol. Sin. 2000, 21, 1125–1128. [Google Scholar]
- Xie, L.; Liu, H.; Qian, R.; Zhang, C.; Wang, J.; Cai, S. The effect of harpagid and harpagoside on immune function and cycle-nucleotides of yin weak mice. [Article in Chinese]. J. Peking Univ. 2001, 33, 283–284. [Google Scholar]
- Li, J.; Zheng, Y.; Zhou, H.; Su, B.; Zheng, R. Iridoid and phenylpropanoid glycosides of Scrophularia ningpoensis inhibit the formation of LTB4 and platelet aggregation. Acad. J. Second Mil. Med. Univ. 1999, 20, 301–303. [Google Scholar]
- Wang, S.; Zheng, Z.; Weng, Y.; Yu, Y.; Zhang, D.; Fan, W.; Dai, R.; Hu, Z. Angiogenesis and anti-angiogenesis activity of Chinese medicinal herbal extracts. Life Sci. 2004, 74, 2467–2478. [Google Scholar] [CrossRef]
- Kim, S.R.; Koo, K.A.; Sung, S.H.; Ma, C.J.; Yoon, J.S.; Kim, Y.C. Iridoids from Scrophularia buergeriana attenuate glutamate-induced neurotoxicity in rat cortical cultures. J. Neurosci. Res. 2003, 74, 948–955. [Google Scholar] [CrossRef]
- Xu, C.; Luo, L.; Tan, R.X. Antidepressant effect of three traditional Chinese medicines in the learned helplessness model. J Ethnopharmacol. 2004, 91, 345–349. [Google Scholar] [CrossRef]
- Kim, S.R.; Kang, S.Y.; Lee, K.Y.; Kim, S.H.; Markelonis, G.J.; Oh, T.H.; Kim, Y.C. Anti-amnestic activity of E-p-methoxycinnamic acid from Scrophularia buergeriana. Brain Res. Cogn. Brain Res. 2003, 17, 454–461. [Google Scholar] [CrossRef]
- Xia, C.; Shi, S.; Ge, W.; Li, C. Advance in pharmacological activities of Scrophularia ningpoensis. [Article in Chinese]. China Pharm. 2008, 11, 121–125. [Google Scholar]
- Huang, C.; Li, Y.; He, X.; Wie, S.; Jiao, B. Apoptotic effect of phenylpropanoid glycosides derived from Scrophularia ningpoensis on liver injured rats. [Article in Chinese]. Chin. J. Integr. Trad. West. Med. Liver Dis. 2004, 14, 160–161. [Google Scholar]
- Li, J.; Zheng, Y.; Zheng, R.; Liu, Z. Antitumour effects of phenylpropanoid glycosides. Chin. Pharm. J. 1995, 30, 269–271. [Google Scholar]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef]
- Cattel, L.; Delprino, L.; Brusa, P.; Dosio, F.; Comoglio, P.M.; Prat, M. Comparison of blocked and non-blocked ricin-antibody immunotoxins against human gastric carcinoma and colorectal adenocarcinoma cell lines. Cancer Immunol. Immunother. 1988, 27, 233–240. [Google Scholar]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar]
- Latz, D.; Dewey, W.C.; Flentje, M.; Lohr, F.; Wenz, F.; Weber, K.J. Migration patterns in pulsed-field electrophoresis of DNA restriction fragments from log-phase mammalian cells after irradiation and incubation for repair. Int. J. Radiat. Biol. 1996, 70, 637–646. [Google Scholar] [CrossRef]
- Konkimalla, V.B.; Blunder, M.; Korn, B.; Soomro, S.A.; Jansen, H.; Chang, W.; Posner, G.H.; Bauer, R.; Efferth, T. Effect of artemisinins and other endoperoxides on nitric oxide-related signaling pathway in RAW 264. 7 mouse macrophage cells. Nitric Oxide 2008, 19, 184–191. [Google Scholar] [CrossRef]
- Developmentel Therapeutics Program NCI/NIH. Available online: http://dtp.nci.nih.gov. (Accessed on 2 February 2012).
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T.; et al. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef]
- Staunton, J.E.; Slonim, D.K.; Coller, H.A.; Tamayo, P.; Angelo, M.J.; Park, J.; Scherf, U.; Lee, J.K.; Reinhold, W.O.; Weinstein, J.N.; et al. Chemosensitivity prediction by transcriptional profiling. Proc. Natl. Acad. Sci. USA 2001, 98, 10787–10792. [Google Scholar]
- Wosikowski, K.; Schuurhuis, D.; Johnso, K.; Paull, K.D.; Myers, T.G.; Weinstein, J.N.; Bates, S.E. Identification of epidermal growth factor receptor and c-erbB2 pathway inhibitors by correlation with gene expression patterns. J. Natl. Cancer Inst. 1997, 89, 1505–1515. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Y. Progress in studying Chinese crude drug figwort root. Guiding J. TCM 2007, 13, 103–105. [Google Scholar]
- Karin, M.; Delhase, M. The IκB kinase (IKK) and NF-κB: Key elements of proinflammatory signalling. Semin. Immunol. 2000, 12, 85–98. [Google Scholar] [CrossRef]
- Delhalle, S.; Blasius, R.; Dicato, M.; Diederich, M. A beginner’s guide to NF-kappaB signaling pathways. Ann. N. Y. Acad. Sci. 2004, 1030, 1–13. [Google Scholar]
- Dvorák, Z.; Vrzal, R.; Maurel, P.; Ulrichová, J. Differential effects of selected natural compounds with anti-inflammatory activity on the glucocorticoid receptor and NF-κB in HeLa cells. Chem. Biol. Interact. 2006, 159, 117–128. [Google Scholar] [CrossRef]
- Enk, R.; Ehehalt, R.; Graham, J.E.; Bierhaus, A.; Remppis, A.; Greten, H.J. Differential effect of Rhizoma coptidis and its main alkaloid compound berberine on TNF-α induced NF-κB translocation in human keratinocytes. J. Ethnopharmacol. 2007, 109, 170–175. [Google Scholar] [CrossRef]
- Ahmed, B.; Al-Rehaily, A.J.; Al-Howiriny, T.A.; El-Sayed, K.A.; Ahmad, M.S. Scropholiosid-D2 and Harpagoside-B: Two new iridoid glycosides from Scrophularia deserti and their antidiabetic and antiinflammatory activity. Biol. Pharm. Bull. 2003, 26, 462–467. [Google Scholar] [CrossRef]
- OMIM® – Online Mendelian Inheritance in Man. Available online: http://www.ncbi.nlm.nih.gov/Omim (Accessed on 13 February 2012).
- Weizmann Institute of Science. Gene Cards®. The human gene compendium. Available online: http://bioinfo.weizmann.ac.il/cards/index.html (Accessed on 13 February 2012).
- Song, J. Effects of polysaccharide from roots of Scrophularia ningpoensis Hemsl on exhaustive swimming time and glycogen stockpile in mice. Food Sci. 2009, 30, 217–219. [Google Scholar]
- Wang, H.; Chen, P.; Zhang, L.P.; Qian, L.N.; Xiao, Y.Y.; Yu, X.Q. Study on the antifatigue function of polysaccharides from Scrophularia ningpoensis Hemsl. J. Wuhan Bot. Res. 2009, 27, 118–120. [Google Scholar]
- Wang, H.; Chen, P.; Zang, L.P.; Xiao, Y.Y.; Jia, H.-Q.; Ma, S.-J. Research about the anti-inflammatory and analgetic activity of the total pigment extract from Scrophularia ningpoensis. Chin. J. Hosp. Pharm. 2008, 28, 1456–1458. [Google Scholar]
- Huang, C.; Li, Y.; He, X.; Wie, S.; Jiang, H. Effect of phenylpropanoid glycosides XS-8 derived from Scrophularia ningpoensis on cAMP in rabbit platelet and PGI2 /TXA2 in rabbit serum. Acad. J. Second Mil. Med. Univ. 2004, 25, 920–921. [Google Scholar]
- He, J.; Dou, H. Yanyan tea to treat 237 cases of chronic pharyngitis. Chin. Nurs. Res. 2001, 15, 54. [Google Scholar]
- Huang, X. 50 Clinical case reports about treatments on acute inflammation of periodontal tissue using combined Chinese and Western medicine. [Article in Chinese]. Chin. J. Trad. West. Med. 2004, 5, 5. [Google Scholar]
- Ni, J. Using Scrophularia ningpoensis as main compound to treat drug induced Atopic dermatitis. [Article in Chinese]. J. Trad. Chin. Med. 2010, 51, 61. [Google Scholar]
- Guo, L. Current state about using combined Chinese and Western medicine for primary bronchogenic lung cancer treatments. [Article in Chinese]. Chin. J. Clin. Oncol. Rehabil. 2001, 8, 103–104. [Google Scholar]
- Huang, G.; Zhao, C.; Han, F.; Zhang, B.; Qiu, H.; Xu, B.; Chen, X.; Hu, P. Clinical study in prophylactic use of Chinese medicine to prevent chemoradiotherapy induced mucositis in nasopharynageal carcinoma. Chin. J. Cancer 2003, 22, 1084–1087. [Google Scholar]
- He, Z.; Jiang, J. Clinical Observation of treating constipation of advanced cancer patients using Yiqizengye decocts [Article in Chinese]. J. Pract. Trad. Chin. Med. 2007, 23, 213. [Google Scholar]
- Leteurtre, F.; Kohlhagen, G.; Paull, K.D.; Pommier, Y. Topoisomerase II inhibition and cytotoxicity of the anthrapyrazoles DuP 937 and DuP 941 (Losoxantrone) in the National Cancer Institute preclinical antitumor drug discovery screen. J. Natl. Cancer Inst. 1994, 86, 1239–1244. [Google Scholar] [CrossRef]
- Efferth, T.; Gebhart, E.; Ross, D.D.; Sauerbrey, A. Identification of gene expression profiles predicting tumor cell response to L-alanosine. Biochem. Pharmacol. 2003, 66, 613–621. [Google Scholar] [CrossRef]
- Efferth, T.; Olbrich, A.; Bauer, R. mRNA expression profiles for the response of human tumor cell lines to the antimalarial drugs artesunate, arteether, and artemether. Biochem. Pharmacol. 2002, 64, 617–623. [Google Scholar] [CrossRef]
- Sohn, S.H.; Ko, E.; Jeon, S.-B.; Lee, B.-J.; Kim, S.-H.; Dong, M.-S.; Lee, D.-U.; Kwak, J.-H.; Kim, Y.; Shin, M. The genome-wide expression profile of Scrophularia ningpoensis-treated thapsigargin-stimulated U-87MG cells. Neurotoxicology 2009, 30, 368–376. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shen, X.; Eichhorn, T.; Greten, H.J.; Efferth, T. Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-κB Signaling of Immortalized and Cancer Cell Lines. Pharmaceuticals 2012, 5, 189-208. https://doi.org/10.3390/ph5020189
Shen X, Eichhorn T, Greten HJ, Efferth T. Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-κB Signaling of Immortalized and Cancer Cell Lines. Pharmaceuticals. 2012; 5(2):189-208. https://doi.org/10.3390/ph5020189
Chicago/Turabian StyleShen, Xiao, Tolga Eichhorn, Henry Johannes Greten, and Thomas Efferth. 2012. "Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-κB Signaling of Immortalized and Cancer Cell Lines" Pharmaceuticals 5, no. 2: 189-208. https://doi.org/10.3390/ph5020189
APA StyleShen, X., Eichhorn, T., Greten, H. J., & Efferth, T. (2012). Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-κB Signaling of Immortalized and Cancer Cell Lines. Pharmaceuticals, 5(2), 189-208. https://doi.org/10.3390/ph5020189