
Association of Bitter Taste Receptor T2R38 Polymorphisms, Oral Microbiota, and Rheumatoid Arthritis

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population, Design, and Sample Collection
2.2. TAS2R38 Sequencing
2.3. 16S rRNA Amplicon Sequencing and Data Analysis
2.4. Statistical Analysis
3. Results
3.1. Participants
3.2. Taste Genetics and RA
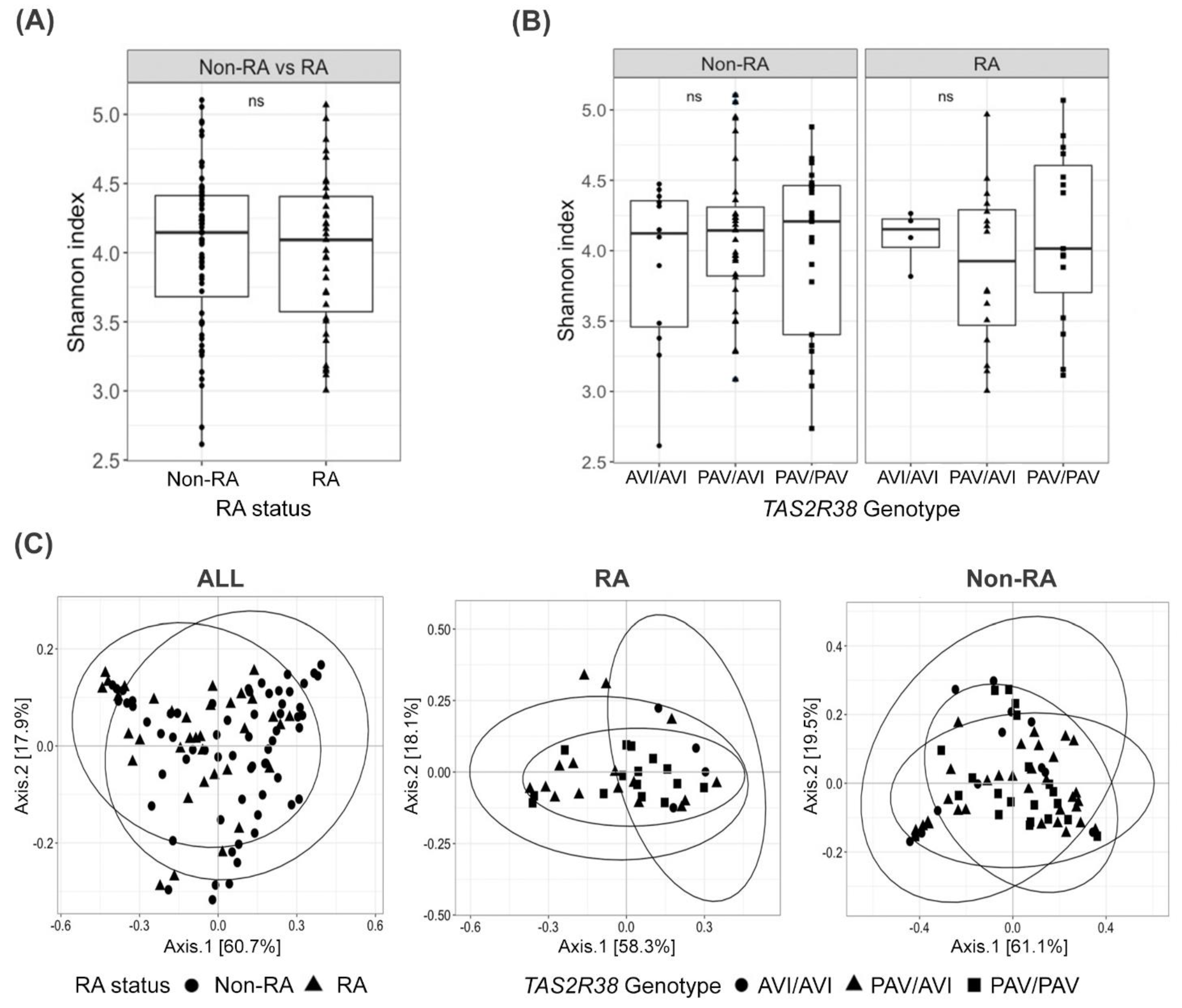
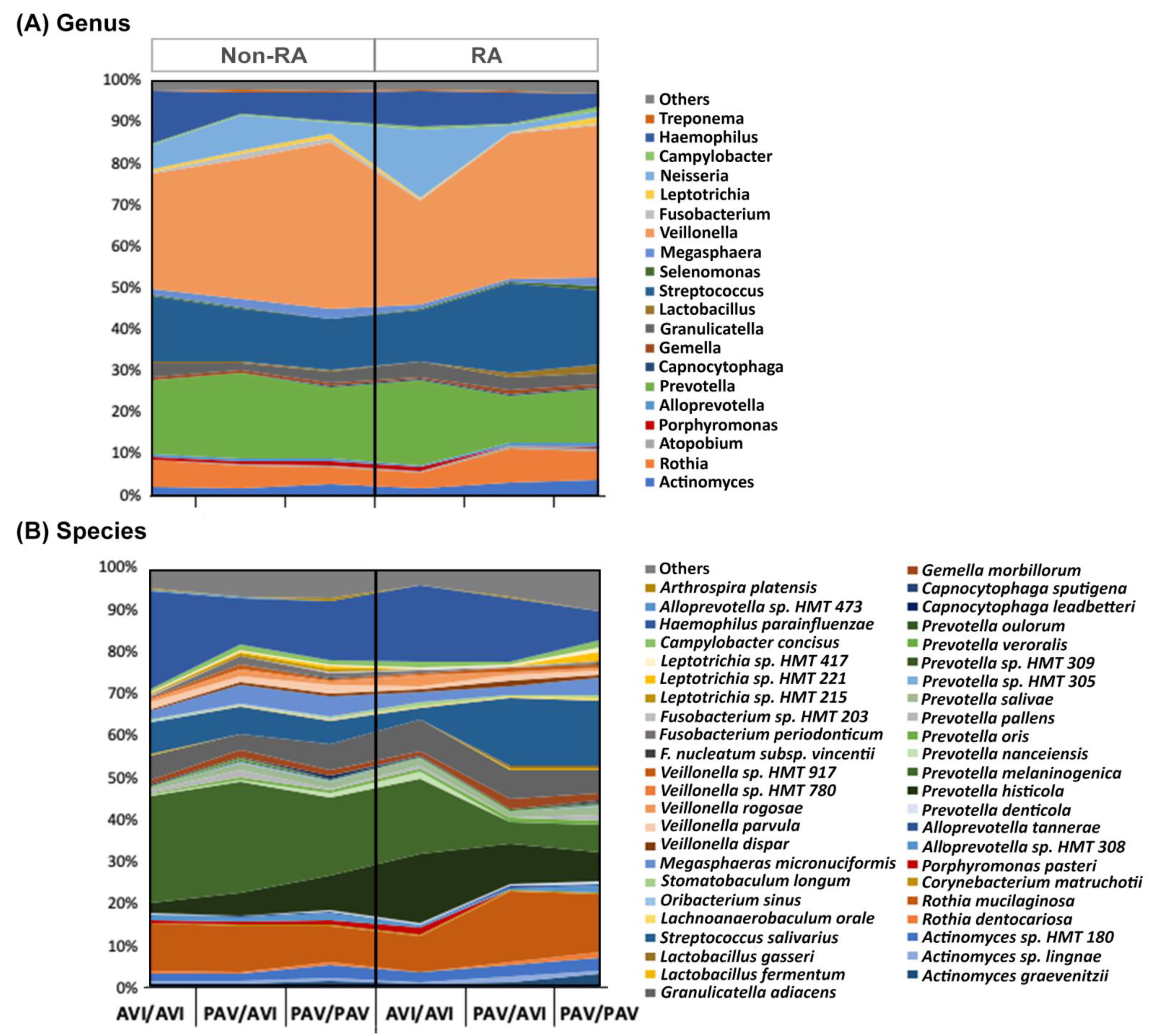
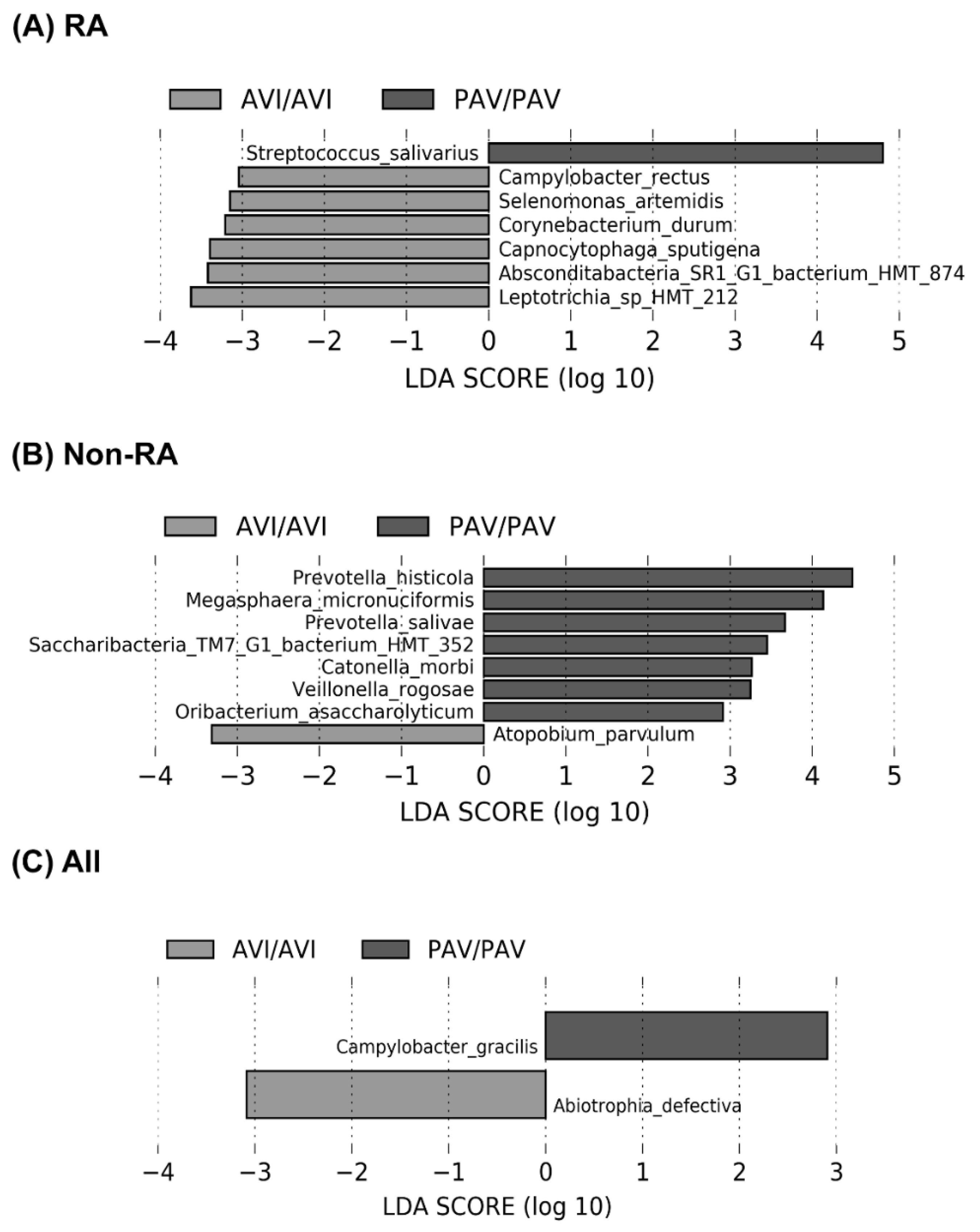
3.3. The Oral Microbiome, TAS2R38 Genotypes, and RA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, P.S. From focal sepsis to periodontal medicine: A century of exploring the role of the oral microbiome in systemic disease. J. Physiol. 2017, 595, 465–476. [Google Scholar] [CrossRef]
- Fuggle, N.R.; Smith, T.O.; Kaul, A.; Sofat, N. Hand to Mouth: A Systematic Review and Meta-Analysis of the Association between Rheumatoid Arthritis and Periodontitis. Front. Immunol. 2016, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lozano, B.; Gonzalez-Febles, J.; Garnier-Rodriguez, J.L.; Dadlani, S.; Bustabad-Reyes, S.; Sanz, M.; Sanchez-Alonso, F.; Sanchez-Piedra, C.; Gonzalez-Davila, E.; Diaz-Gonzalez, F. Association between severity of periodontitis and clinical activity in rheumatoid arthritis patients: A case-control study. Arthritis Res. 2019, 21, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef]
- Potempa, J.; Mydel, P.; Koziel, J. The case for periodontitis in the pathogenesis of rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, J.D.; Fernandes, G.R.; Calderaro, D.C.; Mendonca, S.M.S.; Silva, J.M.; Albiero, M.L.; Cunha, F.Q.; Xiao, E.; Ferreira, G.A.; Teixeira, A.L.; et al. Oral microbial dysbiosis linked to worsened periodontal condition in rheumatoid arthritis patients. Sci. Rep. 2019, 9, 8379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workman, A.D.; Palmer, J.N.; Adappa, N.D.; Cohen, N.A. The Role of Bitter and Sweet Taste Receptors in Upper Airway Immunity. Curr. Allergy Asthma Rep. 2015, 15, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggupilli, A.; Singh, N.; Upadhyaya, J.; Sikarwar, A.S.; Arakawa, M.; Dakshinamurti, S.; Bhullar, R.P.; Duan, K.; Chelikani, P. Analysis of the expression of human bitter taste receptors in extraoral tissues. Mol. Cell. Biochem. 2017, 426, 137–147. [Google Scholar] [CrossRef]
- Carey, R.M.; Lee, R.J. Taste Receptors in Upper Airway Innate Immunity. Nutrients 2019, 11, 2017. [Google Scholar] [CrossRef] [Green Version]
- Shaik, F.A.; Singh, N.; Arakawa, M.; Duan, K.; Bhullar, R.P.; Chelikani, P. Bitter taste receptors: Extraoral roles in pathophysiology. Int. J. Biochem Cell Biol. 2016, 77, 197–204. [Google Scholar] [CrossRef]
- Jaggupilli, A.; Singh, N.; Jesus, V.C.; Duan, K.; Chelikani, P. Characterization of the Binding Sites for Bacterial Acyl Homoserine Lactones (AHLs) on Human Bitter Taste Receptors (T2Rs). ACS Infect. Dis. 2018, 4, 1146–1156. [Google Scholar] [CrossRef]
- Medapati, M.R.; Singh, N.; Bhagirath, A.Y.; Duan, K.; Triggs-Raine, B.; Batista, E.L., Jr.; Chelikani, P. Bitter taste receptor T2R14 detects quorum sensing molecules from cariogenic Streptococcus mutans and mediates innate immune responses in gingival epithelial cells. FASEB J. 2021, 35, e21375. [Google Scholar] [CrossRef]
- Medapati, M.R.; Bhagirath, A.Y.; Singh, N.; Schroth, R.J.; Bhullar, R.P.; Duan, K.; Chelikani, P. Bitter Taste Receptor T2R14 Modulates Gram-Positive Bacterial Internalization and Survival in Gingival Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 9920. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Tizzano, M.; Redding, K.; He, J.; Peng, X.; Jiang, P.; Xu, X.; Zhou, X.; Margolskee, R.F. Gingival solitary chemosensory cells are immune sentinels for periodontitis. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, U.K.; Jorgenson, E.; Coon, H.; Leppert, M.; Risch, N.; Drayna, D. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science 2003, 299, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Risso, D.S.; Mezzavilla, M.; Pagani, L.; Robino, A.; Morini, G.; Tofanelli, S.; Carrai, M.; Campa, D.; Barale, R.; Caradonna, F.; et al. Global diversity in the TAS2R38 bitter taste receptor: Revisiting a classic evolutionary PROPosal. Sci. Rep. 2016, 6, 25506. [Google Scholar] [CrossRef] [Green Version]
- Bufe, B.; Breslin, P.A.S.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Gil, S.; Coldwell, S.; Drury, J.L.; Arroyo, F.; Phi, T.; Saadat, S.; Kwong, D.; Chung, W.O. Genotype-specific regulation of oral innate immunity by T2R38 taste receptor. Mol. Immunol. 2015, 68, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Adappa, N.D.; Zhang, Z.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Scott, T.; Zhao, N.W.; Owens, D.; et al. The bitter taste receptor T2R38 is an independent risk factor for chronic rhinosinusitis requiring sinus surgery. Int. Forum Allergy Rhinol. 2014, 4, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [Green Version]
- Douglas, J.E.; Cohen, N.A. Taste Receptors Mediate Sinonasal Immunity and Respiratory Disease. Int. J. Mol. Sci. 2017, 18, 437. [Google Scholar] [CrossRef] [Green Version]
- Adappa, N.D.; Workman, A.D.; Hadjiliadis, D.; Dorgan, D.J.; Frame, D.; Brooks, S.; Doghramji, L.; Palmer, J.N.; Mansfield, C.; Reed, D.R.; et al. T2R38 genotype is correlated with sinonasal quality of life in homozygous DeltaF508 cystic fibrosis patients. Int. Forum Allergy Rhinol. 2016, 6, 356–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkonen, H.; Brink, M.; Hansson, M.; Lassen, E.; Mathsson-Alm, L.; Holmdahl, R.; Rönnelid, J.; Klareskog, L.; Rantapää-Dahlqvist, S. Associations of antibodies against citrullinated peptides with human leukocyte antigen-shared epitope and smoking prior to the development of rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitchon, C.A.; Chandad, F.; Ferucci, E.D.; Willemze, A.; Ioan-Facsinay, A.; van der Woude, D.; Markland, J.; Robinson, D.; Elias, B.; Newkirk, M.; et al. Antibodies to porphyromonas gingivalis are associated with anticitrullinated protein antibodies in patients with rheumatoid arthritis and their relatives. J. Rheumatol. 2010, 37, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Na. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ 2018, 6, 852–857. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, V.C.; Khan, M.W.; Mittermuller, B.A.; Duan, K.; Hu, P.; Schroth, R.J.; Chelikani, P. Characterization of Supragingival Plaque and Oral Swab Microbiomes in Children With Severe Early Childhood Caries. Front. Microbiol. 2021, 12, 683685. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2 Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Wendy, S.G.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Prodi, D.A.; Drayna, D.; Forabosco, P.; Palmas, M.A.; Maestrale, G.B.; Piras, D.; Pirastu, M.; Angius, A. Bitter taste study in a sardinian genetic isolate supports the association of phenylthiocarbamide sensitivity to the TAS2R38 bitter receptor gene. Chem. Senses 2004, 29, 697–702. [Google Scholar] [CrossRef] [Green Version]
- Peschken, C.A.; Hitchon, C.A.; Robinson, D.B.; Smolik, I.; Barnabe, C.R.; Prematilake, S.; El-Gabalawy, H.S. Rheumatoid arthritis in a north american native population: Longitudinal followup and comparison with a white population. J. Rheumatol. 2010, 37, 1589–1595. [Google Scholar] [CrossRef] [Green Version]
- Wells, P.M.; Adebayo, A.S.; Bowyer, R.C.E.; Freidin, M.B.; Finckh, A.; Strowig, T.; Lesker, T.R.; Alpizar-Rodriguez, D.; Gilbert, B.; Kirkham, B.; et al. Associations between gut microbiota and genetic risk for rheumatoid arthritis in the absence of disease: A cross-sectional study. Lancet. Rheumatol. 2020, 2, e418–e427. [Google Scholar] [CrossRef]
- Kroese, J.M.; Brandt, B.W.; Buijs, M.J.; Crielaard, W.; Lobbezoo, F.; Loos, B.G.; van Boheemen, L.; van Schaardenburg, D.; Zaura, E.; Volgenant, C.M.C. The oral microbiome in early rheumatoid arthritis patients and individuals at risk differs from healthy controls. Arthritis Rheumatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, K.; Fei, G.; Lundmark, A.; Benchimol, D.; Lee, L.; Hu, Y.O.O.; Kats, A.; Saevarsdottir, S.; Catrina, A.I.; Klinge, B.; et al. Periodontal Health and Oral Microbiota in Patients with Rheumatoid Arthritis. J. Clin. Med. 2019, 8, 630. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Do, T.; Mankia, K.; Meade, J.; Hunt, L.; Clerehugh, V.; Speirs, A.; Tugnait, A.; Emery, P.; Devine, D. Dysbiosis in the oral microbiomes of anti-CCP positive individuals at risk of developing rheumatoid arthritis. Ann. Rheum. Dis. 2021, 80, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Esberg, A.; Johansson, L.; Johansson, I.; Dahlqvist, S.R. Oral Microbiota Identifies Patients in Early Onset Rheumatoid Arthritis. Microorganisms 2021, 9, 1657. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Joshua, V.; Artacho, A.; Abdollahi-Roodsaz, S.; Öckinger, J.; Kullberg, S.; Sköld, M.; Eklund, A.; Grunewald, J.; Clemente, J.C.; et al. The lung microbiota in early rheumatoid arthritis and autoimmunity. Microbiome 2016, 4, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantapää-Dahlqvist, S.; de Jong, B.A.W.; Berglin, E.; Hallmans, G.; Wadell, G.; Stenlund, H.; Sundin, U.; van Venrooij, W.J. Antibodies against cyclic citrullinated peptide and IgA rheumatoid factor predict the development of rheumatoid arthritis. Arthritis Rheum. 2003, 48, 2741–2749. [Google Scholar] [CrossRef]
- Konig, M.F.; Andrade, F. A Critical Reappraisal of Neutrophil Extracellular Traps and NETosis Mimics Based on Differential Requirements for Protein Citrullination. Front. Immunol. 2016, 7, 461. [Google Scholar] [CrossRef] [Green Version]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter actinomycetemcomitans–induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 369ra176. [Google Scholar] [CrossRef] [Green Version]
- Mankia, K.; Cheng, Z.; Do, T.; Hunt, L.; Meade, J.; Kang, J.; Clerehugh, V.; Speirs, A.; Tugnait, A.; Hensor, E.M.A.; et al. Prevalence of Periodontal Disease and Periodontopathic Bacteria in Anti-Cyclic Citrullinated Protein Antibody-Positive At-Risk Adults Without Arthritis. JAMA Netw. Open 2019, 2, e195394. [Google Scholar] [CrossRef] [PubMed]
- Loutan, L.; Alpizar-Rodriguez, D.; Courvoisier, D.S.; Finckh, A.; Mombelli, A.; Giannopoulou, C. Periodontal status correlates with anti-citrullinated protein antibodies in first-degree relatives of individuals with rheumatoid arthritis. J. Clin. Periodontol. 2019, 46, 690–698. [Google Scholar] [CrossRef]
- Lundberg, K.; Wegner, N.; Yucel-Lindberg, T.; Venables, P.J. Periodontitis in RA-the citrullinated enolase connection. Nat. Rev. Rheumatol. 2010, 6, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Mikuls, T.R.; Payne, J.B.; Reinhardt, R.A.; Thiele, G.M.; Maziarz, E.; Cannella, A.C.; Holers, V.M.; Kuhn, K.A.; O’Dell, J.R. Antibody responses to Porphyromonas gingivalis (P. gingivalis) in subjects with rheumatoid arthritis and periodontitis. Int. Immunopharmacol. 2009, 9, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Schwenzer, A.; Quirke, A.M.; Marzeda, A.M.; Wong, A.; Montgomery, A.B.; Sayles, H.R.; Eick, S.; Gawron, K.; Chomyszyn-Gajewska, M.; Lazarz-Bartyzel, K.; et al. Association of Distinct Fine Specificities of Anti-Citrullinated Peptide Antibodies With Elevated Immune Responses to Prevotella intermedia in a Subgroup of Patients With Rheumatoid Arthritis and Periodontitis. Arthritis Rheumatol. 2017, 69, 2303–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Bañuelos, E.; Johansson, L.; Konig, M.F.; Lundquist, A.; Paz, M.; Buhlin, K.; Johansson, A.; Rantapää-Dahlqvist, S.; Andrade, F. Exposure to Aggregatibacter actinomycetemcomitans before Symptom Onset and the Risk of Evolving to Rheumatoid Arthritis. J. Clin. Med. 2020, 9, 1906. [Google Scholar] [CrossRef] [PubMed]
- Moura, M.F.; Cota, L.O.M.; Silva, T.A.; Cortelli, S.C.; Ferreira, G.A.; López, M.M.; Cortelli, J.R.; Costa, F.O. Clinical and microbiological effects of non-surgical periodontal treatment in individuals with rheumatoid arthritis: A controlled clinical trial. Odontology 2021, 109, 484–493. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Non-RA (n = 64) | RA (n = 35) | p-Value a |
|---|---|---|---|
| Demographics | |||
| Age, mean ± SD | 43.3 ± 11.53 | 51.2 ± 13.8 | 0.004 |
| Sex, female | 32 (50.0%) | 29 (82.9%) | 0.001 |
| RA features | |||
| ACPA positive b | 2 (3.4%) | 23 (79.3%) | <0.001 |
| ACPA titer if positive b,c | 155 (19,155) | 300 (32.5, 300) | 0.35 |
| RF positive b | 8 (13.6%) | 22 (76.9%%) | <0.001 |
| RF titer if positive b | 37 (22.3, 50.1) | 382 (172, 1047.8) | <0.001 |
| Current DMARD | NA | 21 (65.6%) | - |
| Swollen joint count | NA | 1.0 (0.0, 3.3) | - |
| CRP (mg/L) | 2.0 (1.0, 4.7) | 6.0 (1.0, 15.0) | 0.02 |
| Overall health | |||
| Current smoking | 47 (79.7%) | 21 (75.0%) | 0.62 |
| Diabetes mellitus | 14 (23.3%) | 5 (15.2%) | 0.35 |
| BMI | 28.6 (25.5, 33.0) | 28.23 (24.3, 32.8) | 0.75 |
| Currently on antibiotics | 1 (1.7%) | 3 (10.7%) | 0.06 |
| Oral Health | |||
| Denture | 14 (23.7%) | 10 (33.3%) | 0.33 |
| Gum bleeding | 0.17 | ||
| Never | 15 (27.8%) | 11 (50.0%) | |
| Rarely | 20 (37.0%) | 5 (22.7%) | |
| Sometimes/often/always | 19 (35.2%) | 6 (27.3%) | |
| Metallic taste | 0.91 | ||
| Never | 37 (68.5%) | 14 (63.6%) | |
| Rarely | 6 (11.1%) | 3 (13.6%) | |
| Sometimes/often/always | 11 (20.4%) | 5 (22.7%) | |
| Tender or painful gum | 0.31 | ||
| Never | 27 (50.0%) | 10 (45.5%) | |
| Rarely | 9 (16.7%) | 7 (31.8%) | |
| Sometimes/often/always | 18 (33.3%) | 5 (22.7%) | |
| Feeling of loose teeth | 0.95 | ||
| Never | 38 (70.4%) | 15 (68.2%) | |
| Rarely | 10 (18.5%) | 4 (18.2%) | |
| Sometimes/often/always | 6 (11.1%) | 3 (13.6%) | |
| Brush/week | 0.44 | ||
| Less than 6 times | 34 (60.7%) | 13 (50.0%) | |
| 7 to 14 times | 18 (32.1%) | 9 (34.6%) | |
| More than 15 times | 4 (7.1%) | 4 (15.4%) | |
| Floss/week | 0.31 | ||
| Less than 6 times | 47 (83.9%) | 18 (69.2%) | |
| 7 to 14 times | 7 (12.5%) | 6 (23.1%) | |
| 15+ times | 2 (3.6%) | 2 (7.7%) | |
| Dentist visit | 0.24 | ||
| Never | 7 (12.3%) | 16 (59.3%) | |
| 1–2 times per year | 44 (77.2%) | 16 (59.3%) | |
| 2+ times per year | 6 (10.5%) | 5 (18.5%) |
| Non-Taster AVI/AVI (n = 16) | Taster PAV/AVI (n = 47) | Super Taster PAV/PAV (n = 36) | p-Value a | |
|---|---|---|---|---|
| Demographics | ||||
| Age, mean ± SD | 44.4 ± 11.1 | 44.7 ± 13.2 | 48.8 ± 13.1 | 0.35 |
| Sex, female | 12 (75.0%) | 24 (54.5%) | 25 (71.4%) | 0.18 |
| Clinical features | ||||
| RA | 4 (25.0%) | 16 (34.0%) | 15 (41.7%) | 0.49 |
| ACPA positive b | 2 (13.3%) | 11 (27.5%) | 12 (35.4%) | 0.26 |
| ACPA titer if positive | 82 (-) | 223 (22.5, 300) | 300 (885, 300.0) | 0.34 |
| RF positive b | 4 (26.7%) | 11 (27.5%) | 15 (45.5%) | 0.21 |
| RF titer if positive | 243 (38, 243) | 208 (56.4,4525) | 109 (26.7, 1010) | 0.9 |
| Current DMARD | 4 (26.7%) | 7 (18.42%) | 10 (32.3%) | 0.41 |
| Overall health | ||||
| Current smoking | 10 (66.7%) | 30 (76.9%) | 28 (84.8%) | 0.36 |
| Diabetes mellitus | 3 (20.0%) | 6 (13.6%) | 10 (29.4%) | 0.23 |
| BMI | 31.8 (27.8, 36.6) | 27.1 (23.3, 32.3) | 28.2 (24.8, 31.3) | 0.11 |
| Currently on antibiotics | 1 (6.7%) | 1 (2.6%) | 2 (6.2%) | 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Jesus, V.C.; Singh, M.; Schroth, R.J.; Chelikani, P.; Hitchon, C.A. Association of Bitter Taste Receptor T2R38 Polymorphisms, Oral Microbiota, and Rheumatoid Arthritis. Curr. Issues Mol. Biol. 2021, 43, 1460-1472. https://doi.org/10.3390/cimb43030103
de Jesus VC, Singh M, Schroth RJ, Chelikani P, Hitchon CA. Association of Bitter Taste Receptor T2R38 Polymorphisms, Oral Microbiota, and Rheumatoid Arthritis. Current Issues in Molecular Biology. 2021; 43(3):1460-1472. https://doi.org/10.3390/cimb43030103
Chicago/Turabian Stylede Jesus, Vivianne Cruz, Manu Singh, Robert J. Schroth, Prashen Chelikani, and Carol A. Hitchon. 2021. "Association of Bitter Taste Receptor T2R38 Polymorphisms, Oral Microbiota, and Rheumatoid Arthritis" Current Issues in Molecular Biology 43, no. 3: 1460-1472. https://doi.org/10.3390/cimb43030103
APA Stylede Jesus, V. C., Singh, M., Schroth, R. J., Chelikani, P., & Hitchon, C. A. (2021). Association of Bitter Taste Receptor T2R38 Polymorphisms, Oral Microbiota, and Rheumatoid Arthritis. Current Issues in Molecular Biology, 43(3), 1460-1472. https://doi.org/10.3390/cimb43030103