
Piperazine Derivative Stabilizes Actin Filaments in Primary Fibroblasts and Binds G-Actin In Silico

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Compounds
2.2. Mice
2.3. Plasmids
2.4. Primary Hippocampal Neuronal Cultures
2.5. Calcium Phosphate Transfection of Primary Hippocampal Cultures
2.6. Dendritic Spine Analysis in Primary Hippocampal Neuronal Cultures
2.7. Calcium Imaging
2.8. Fibroblast Culture
2.9. Analysis of F-Actin Structure
2.10. In Vitro Statistical Analysis
2.11. Molecular Models Selection
2.12. Molecular Docking
2.13. Molecular Dynamics
2.14. Conformation Analysis and Interaction Visualization
2.15. In Silico Clustering and Statistical Analysis
2.16. Constant Binding Calculation
3. Results
3.1. 51164 Does Not Upregulate Decreased SOCE in Postsynaptic Spines of Primary Hippocampal Neurons with CaMKIIβ Knockdown
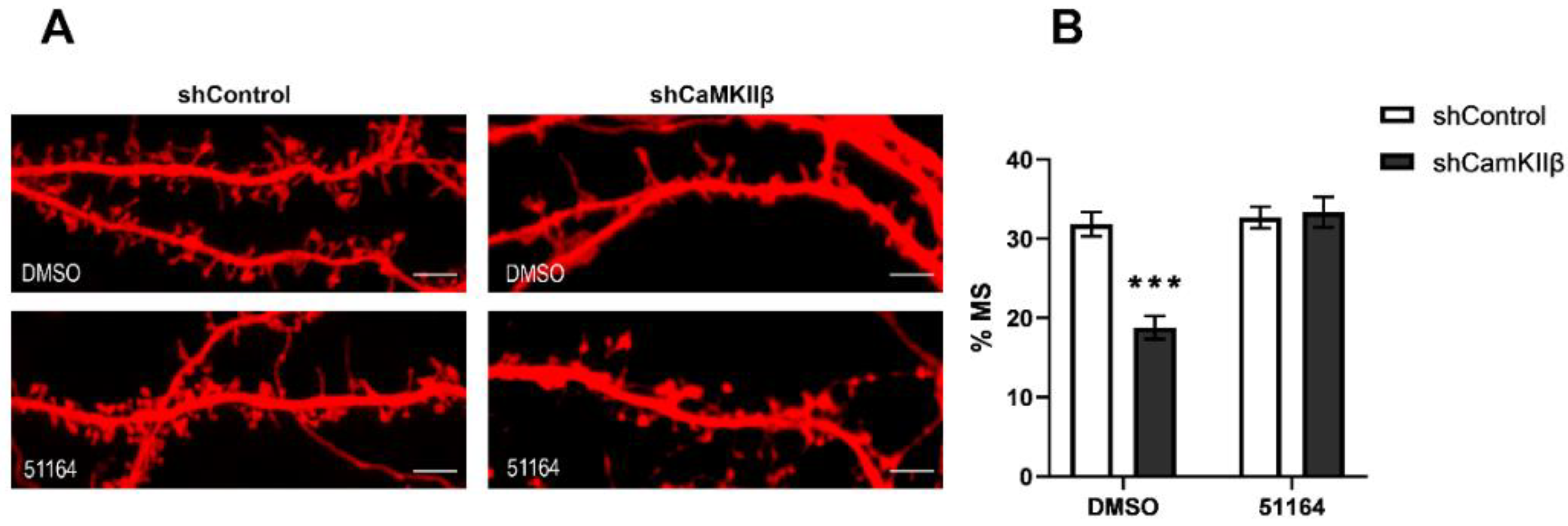
3.2. 51164 Recovers Mushroom Spine Percentage in CaMKIIβ Knockdown Hippocampal Cultures
3.3. 51164 Protects Actin Filaments in Cytochalasin D Treated Fibroblasts
3.4. Molecular Docking Study
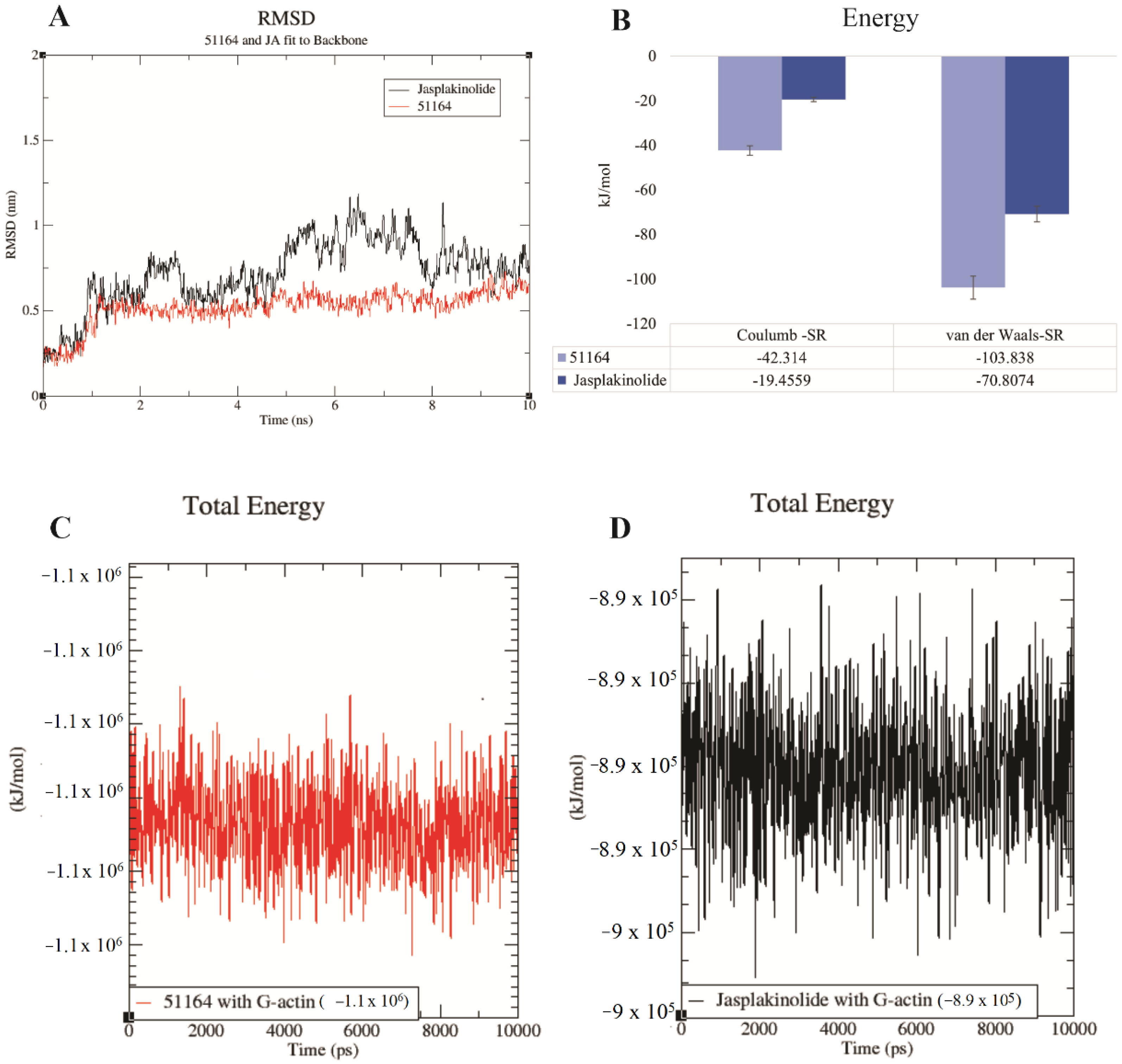
3.5. Molecular Dynamics Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2021 Alzheimer’s Disease Facts and Figures. Alzheimers Dement. 2021, 17, 327–406. [Google Scholar] [CrossRef] [PubMed]
- Hanseeuw, B.J.; Betensky, R.A.; Jacobs, H.I.L.; Schultz, A.P.; Sepulcre, J.; Becker, J.A.; Cosio, D.M.O.; Farrell, M.; Quiroz, Y.T.; Mormino, E.C.; et al. Association of Amyloid and Tau with Cognition in Preclinical Alzheimer Disease: A Longitudinal Study. JAMA Neurol. 2019, 76, 915–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, J.; Harris, K.M. Do Thin Spines Learn to Be Mushroom Spines That Remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef]
- Pchitskaya, E.; Bezprozvanny, I. Dendritic Spines Shape Analysis—Classification or Clusterization? Perspective. Front. Synaptic Neurosci. 2020, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Matsuba, Y.; Mihira, N.; Takano, J.; Nilsson, P.; Itohara, S.; Iwata, N.; Saido, T.C. Single App Knock-in Mouse Models of Alzheimer’s Disease. Nat. Neurosci. 2014, 17, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, H.; Liu, J.; Popugaeva, E.; Xu, N.J.; Feske, S.; White, C.L.; Bezprozvanny, I. Reduced Synaptic STIM2 Expression and Impaired Store-Operated Calcium Entry Cause Destabilization of Mature Spines in Mutant Presenilin Mice. Neuron 2014, 82, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wu, L.; Pchitskaya, E.; Zakharova, O.; Saito, T.; Saido, T.; Bezprozvanny, I. Neuronal Store-Operated Calcium Entry and Mushroom Spine Loss in Amyloid Precursor Protein Knock-In Mouse Model of Alzheimer’s Disease. J. Neurosci. 2015, 35, 13275–13286. [Google Scholar] [CrossRef] [Green Version]
- Popugaeva, E.; Pchitskaya, E.; Speshilova, A.; Alexandrov, S.; Zhang, H.; Vlasova, O.; Bezprozvanny, I. STIM2 Protects Hippocampal Mushroom Spines from Amyloid Synaptotoxicity. Mol. Neurodegener. 2015, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Popugaeva, E.; Supnet, C.; Bezprozvanny, I. Presenilins, Deranged Calcium Homeostasis, Synaptic Loss and Dysfunction in Alzheimer’s Disease. Messenger 2012, 1, 53–62. [Google Scholar] [CrossRef]
- Tackenberg, C.; Ghori, A.; Brandt, R. Thin, Stubby or Mushroom: Spine Pathology in Alzheimers Disease. Curr. Alzheimer Res. 2009, 6, 261–268. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.; Wu, L.; Pchitskaya, E.; Zakharova, O.; Fon Tacer, K.; Bezprozvanny, I. Store-Operated Calcium Channel Complex in Postsynaptic Spines: A New Therapeutic Target for Alzheimer’s Disease Treatment. J. Neurosci. 2016, 36, 11837–11850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernyuk, D.; Zernov, N.; Kabirova, M.; Bezprozvanny, I.; Popugaeva, E. Antagonist of Neuronal Store-Operated Calcium Entry Exerts Beneficial Effects in Neurons Expressing PSEN1ΔE9 Mutant Linked to Familial Alzheimer Disease. Neuroscience 2019, 410, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Popugaeva, E.; Chernyuk, D.; Zhang, H.; Postnikova, T.Y.; Pats, K.; Fedorova, E.; Poroikov, V.; Zaitsev, A.V.; Bezprozvanny, I. Derivatives of Piperazines as Potential Therapeutic Agents for Alzheimer’s Disease. Mol. Pharmacol. 2019, 95, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zernov, N.; Bezprozvanny, I.; Popugaeva, E. CaMKIIβ Knockdown Decreases Store-Operated Calcium Entry in Hippocampal Dendritic Spines. IBRO Neurosci. Rep. 2022, 12, 90. [Google Scholar] [CrossRef] [PubMed]
- Waldner, C.; Roose, M.; Ryffel, G.U. Red Fluorescent Xenopus Laevis: A New Tool for Grafting Analysis. BMC Dev. Biol. 2009, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Thaler, C.; Koushik, S.V.; Puhl, H.L., III; Blank, P.S.; Vogel, S.S. Structural Rearrangement of CaMKIIalpha Catalytic Domains Encodes Activation. Proc. Natl. Acad. Sci. USA 2009, 106, 6369–6374. [Google Scholar] [CrossRef] [Green Version]
- Shen, K.; Teruel, M.N.; Subramanian, K.; Meyer, T. CaMKIIbeta Functions as an F-Actin Targeting Module That Localizes CaMKIIalpha/Beta Heterooligomers to Dendritic Spines. Neuron 1998, 21, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Ehlenberger, D.B.; Dickstein, D.L.; Hof, P.R.; Wearne, S.L. Automated Three-Dimensional Detection and Shape Classification of Dendritic Spines from Fluorescence Microscopy Images. PLoS ONE 2008, 3, e1997. [Google Scholar] [CrossRef] [Green Version]
- Uppal, S.O.; Voronine, D.V.; Wendt, E.; Heckman, C.A. Morphological Fractal Analysis of Shape in Cancer Cells Treated with Combinations of Microtubule-Polymerizing and -Depolymerizing Agents. Microsc. Microanal. 2010, 16, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Rajković, N.; Krstonošić, B.; Milošević, N. Box-Counting Method of 2D Neuronal Image: Method Modification and Quantitative Analysis Demonstrated on Images from the Monkey and Human Brain. Comput. Math. Methods Med. 2017, 2017, 8967902. [Google Scholar] [CrossRef]
- Revittser, A.; Selin, I.; Negulyaev, Y.; Chubinskiy-Nadezhdin, V. The Analysis of F-Actin Structure of Mesenchymal Stem Cells by Quantification of Fractal Dimension. PLoS ONE 2021, 16, e0260727. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Mackerell, A.D. CHARMM Additive and Polarizable Force Fields for Biophysics and Computer-Aided Drug Design. Biochim. Biophys. Acta 2015, 1850, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, K.R. Computer Review of ChemDraw Ultra 12.0. J. Am. Chem. Soc. 2011, 133, 8388. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadava, U. Search Algorithms and Scoring Methods in Protein-Ligand Docking. Endocrinol. Int. J. 2018, 6, 359–367. [Google Scholar] [CrossRef]
- Hassan, N.M.; Alhossary, A.A.; Mu, Y.; Kwoh, C.K. Protein-Ligand Blind Docking Using QuickVina-W With Inter-Process Spatio-Temporal Integration. Sci. Rep. 2017, 7, 15451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Walker, B.; Man, V.H.; Ren, P.; Wang, J. Recent Progress in General Force Fields of Small Molecules. Curr. Opin. Struct. Biol. 2022, 72, 187–193. [Google Scholar] [CrossRef]
- Gil Pineda, L.I.; Milko, L.N.; He, Y. Performance of CHARMM36m with Modified Water Model in Simulating Intrinsically Disordered Proteins: A Case Study. Biophys. Rep. 2020, 6, 80–87. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1998, 103, 8577. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.J. XMGRACE, Version 5.1.19; Center for Coastal and Land-Margin Research, Oregon Graduate Institute of Science and Technology: Beaverton, OR, USA, 2005. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Lamm, G. The Poisson–Boltzmann Equation. Rev. Comput. Chem. 2003, 19, 147–365. [Google Scholar] [CrossRef]
- Landis, D.M.D.; Reese, T.S. Cytoplasmic Organization in Cerebellar Dendritic Spines. J. Cell Biol. 1983, 97, 1169–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuriu, T.; Inoue, A.; Bito, H.; Sobue, K.; Okabe, S. Differential Control of Postsynaptic Density Scaffolds via Actin-Dependent and -Independent Mechanisms. J. Neurosci. 2006, 26, 7693–7706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.; Kaech, S.; Knutti, D.; Matus, A. Rapid Actin-Based Plasticity in Dendritic Spines. Neuron 1998, 20, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, K.I.; Nagai, T.; Miyawaki, A.; Hayashi, Y. Rapid and Persistent Modulation of Actin Dynamics Regulates Postsynaptic Reorganization Underlying Bidirectional Plasticity. Nat. Neurosci. 2004, 7, 1104–1112. [Google Scholar] [CrossRef]
- Star, E.N.; Kwiatkowski, D.J.; Murthy, V.N. Rapid Turnover of Actin in Dendritic Spines and Its Regulation by Activity. Nat. Neurosci. 2002, 5, 239–246. [Google Scholar] [CrossRef]
- Cheng, D.; Hoogenraad, C.C.; Rush, J.; Ramm, E.; Schlager, M.A.; Duong, D.M.; Xu, P.; Wijayawardana, S.R.; Hanfelt, J.; Nakagawa, T.; et al. Relative and Absolute Quantification of Postsynaptic Density Proteome Isolated from Rat Forebrain and Cerebellum. Mol. Cell. Proteom. 2006, 5, 1158–1170. [Google Scholar] [CrossRef] [Green Version]
- Hering, H.; Sheng, M. Activity-Dependent Redistribution and Essential Role of Cortactin in Dendritic Spine Morphogenesis. J. Neurosci. 2003, 23, 11759–11769. [Google Scholar] [CrossRef]
- Ivanov, A.; Esclapez, M.; Pellegrino, C.; Shirao, T.; Ferhat, L. Drebrin A Regulates Dendritic Spine Plasticity and Synaptic Function in Mature Cultured Hippocampal Neurons. J. Cell Sci. 2009, 122, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Terry-Lorenzo, R.T.; Roadcap, D.W.; Otsuka, T.; Blanpied, T.A.; Zamorano, P.L.; Garner, C.C.; Shenolikar, S.; Ehlers, M.D. Neurabin/Protein Phosphatase-1 Complex Regulates Dendritic Spine Morphogenesis and Maturation. Mol. Biol. Cell 2005, 16, 2349–2362. [Google Scholar] [CrossRef]
- Wang, G.; Simon, D.J.; Wu, Z.; Belsky, D.M.; Heller, E.; O’Rourke, M.K.; Hertz, N.T.; Molina, H.; Zhong, G.; Tessier-Lavigne, M.; et al. Structural Plasticity of Actin-Spectrin Membrane Skeleton and Functional Role of Actin and Spectrin in Axon Degeneration. eLife 2019, 8, e38730. [Google Scholar] [CrossRef] [PubMed]
- Zonderland, J.; Wieringa, P.; Moroni, L. A Quantitative Method to Analyse F-Actin Distribution in Cells. MethodsX 2019, 6, 2562–2569. [Google Scholar] [CrossRef] [PubMed]
- Fuseler, J.W.; Millette, C.F.; Davis, J.M.; Carver, W. Fractal and Image Analysis of Morphological Changes in the Actin Cytoskeleton of Neonatal Cardiac Fibroblasts in Response to Mechanical Stretch. Microsc. Microanal. 2007, 13, 133–143. [Google Scholar] [CrossRef]
- Qian, A.R.; Li, D.; Han, J.; Gao, X.; Di, S.M.; Zhang, W.; Hu, L.F.; Shang, P. Fractal Dimension as a Measure of Altered Actin Cytoskeleton in MC3T3-E1 Cells under Simulated Microgravity Using 3-D/2-D Clinostats. IEEE Trans. Biomed. Eng. 2012, 59, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, L.A.; Goda, Y. Actin in Action: The Interplay between the Actin Cytoskeleton and Synaptic Efficacy. Nat. Rev. Neurosci. 2008, 9, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W.; Mannherz, H.G.; Suck, D.; Pai, E.F.; Holmes, K.C. Atomic Structure of the Actin: DNase I Complex. Nature 1990, 347, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Kale, A. Actin Polymerization in Apicomplexan: A Structural, Functional and Evolutionary Analysis; Springer: Singapore, 2019; pp. 93–97. [Google Scholar]
- Wang, H.; Robinson, R.C.; Burtnick, L.D. The Structure of Native G-Actin. Cytoskeleton 2010, 67, 456–465. [Google Scholar] [CrossRef]
- Oztug Durer, Z.A.; Diraviyam, K.; Sept, D.; Kudryashov, D.S.; Reisler, E. F-Actin Structure Destabilization and DNase-I Binding Loop Fluctuations: Mutational Cross-Linking and Electron Microscopy Analysis of the Loop States and Effects on F-Actin. J. Mol. Biol. 2010, 395, 544. [Google Scholar] [CrossRef] [Green Version]
- Graceffa, P.; Dominguez, R. Crystal Structure of Monomeric Actin in the ATP State. Structural Basis of Nucleotide-Dependent Actin Dynamics. J. Biol. Chem. 2003, 278, 34172–34180. [Google Scholar] [CrossRef] [Green Version]
- Oda, T.; Iwasa, M.; Aihara, T.; Maéda, Y.; Narita, A. The Nature of the Globular- to Fibrous-Actin Transition. Nature 2009, 457, 441–445. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin Structure and Function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubb, M.R.; Spector, I.; Beyer, B.B.; Fosen, K.M. Effects of Jasplakinolide on the Kinetics of Actin Polymerization: An Explanation for Certain In Vivo Observations. J. Biol. Chem. 2000, 275, 5163–5170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimaite, R.; Seikowski, J.; Schimpfhauser, J.; Kostiuk, G.; Gilat, T.; D’Este, E.; Schnorrenberg, S.; Lukinavičius, G. Efflux Pump Insensitive Rhodamine–Jasplakinolide Conjugates for G- and F-Actin Imaging in Living Cells. Org. Biomol. Chem. 2020, 18, 2929–2937. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fan, Y.; Dube, D.K.; Sanger, J.M.; Sanger, J.W. Jasplakinolide Reduces Actin and Tropomyosin Dynamics during Myofibrillogenesis. Cytoskeleton 2014, 71, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayot, C.; Debeir, O.; Van Ham, P.; Van Damme, M.; Kiss, R.; Decaestecker, C. Characterization of the Activities of Actin-Affecting Drugs on Tumor Cell Migration. Toxicol. Appl. Pharmacol. 2006, 211, 30–40. [Google Scholar] [CrossRef]
- Stehn, J.; Schevzov, G.; O’Neill, G.; Gunning, P. Specialisation of the Tropomyosin Composition of Actin Filaments Provides New Potential Targets for Chemotherapy. Curr. Cancer Drug Targets 2006, 6, 245–256. [Google Scholar] [CrossRef]
- Schweikart, K.; Guo, L.; Shuler, Z.; Abrams, R.; Chiao, E.T.; Kolaja, K.L.; Davis, M. The Effects of Jaspamide on Human Cardiomyocyte Function and Cardiac Ion Channel Activity. Toxicol. Vitr. 2013, 27, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Trendowski, M.; Wong, V.; Wellington, K.; Hatfield, S.; Fondy, T.P. Tolerated Doses in Zebrafish of Cytochalasins and Jasplakinolide for Comparison with Tolerated Doses in Mice in the Evaluation of Pre-Clinical Activity of Microfilament-Directed Agents in Tumor Model Systems In Vivo. In Vivo 2014, 28, 1021–1032. [Google Scholar]
- Foerster, F.; Braig, S.; Chen, T.; Altmann, K.H.; Vollmar, A.M. Pharmacological Characterization of Actin-Binding (-)-Doliculide. Bioorg. Med. Chem. 2014, 22, 5117–5122. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Parameter | Meaning |
|---|---|---|
| Quantity primary conformers | 20 | |
| 1 | Exhaustiveness | 200 |
| 2 | Repeatability experiment | 5 |
| 3 | Volume virtual boxing | 96314 Å3 |
| 4 | RMS deviation | ≤2 Å |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zernov, N.; Ghamaryan, V.; Makichyan, A.; Melenteva, D.; Hunanyan, L.; Popugaeva, E. Piperazine Derivative Stabilizes Actin Filaments in Primary Fibroblasts and Binds G-Actin In Silico. Curr. Issues Mol. Biol. 2022, 44, 5191-5208. https://doi.org/10.3390/cimb44110353
Zernov N, Ghamaryan V, Makichyan A, Melenteva D, Hunanyan L, Popugaeva E. Piperazine Derivative Stabilizes Actin Filaments in Primary Fibroblasts and Binds G-Actin In Silico. Current Issues in Molecular Biology. 2022; 44(11):5191-5208. https://doi.org/10.3390/cimb44110353
Chicago/Turabian StyleZernov, Nikita, Viktor Ghamaryan, Ani Makichyan, Daria Melenteva, Lernik Hunanyan, and Elena Popugaeva. 2022. "Piperazine Derivative Stabilizes Actin Filaments in Primary Fibroblasts and Binds G-Actin In Silico" Current Issues in Molecular Biology 44, no. 11: 5191-5208. https://doi.org/10.3390/cimb44110353
APA StyleZernov, N., Ghamaryan, V., Makichyan, A., Melenteva, D., Hunanyan, L., & Popugaeva, E. (2022). Piperazine Derivative Stabilizes Actin Filaments in Primary Fibroblasts and Binds G-Actin In Silico. Current Issues in Molecular Biology, 44(11), 5191-5208. https://doi.org/10.3390/cimb44110353