

Increased Levels of Autoantibodies against ROS-Modified Proteins in Depressed Individuals with Decrease in Antibodies against SARS-CoV-2 Antigen (S1-RBD)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Serum Samples
2.2. Modification of BSA and Lysozyme by ROS
2.3. Structural Modifications
2.4. Detection of Serum MDA Contents
- ΔA1 represents OD of sample—OD of control
- ΔA2 represents OD of standard—OD of blank
- C is the concentration of standard (10 nmol/mL)
2.5. Cytokines IFN-γ and TNF-α
2.6. Determination of Protein Bound Carbonyl Contents
2.7. Direct and Inhibition ELISA
2.8. Statistical Evaluation
3. Results
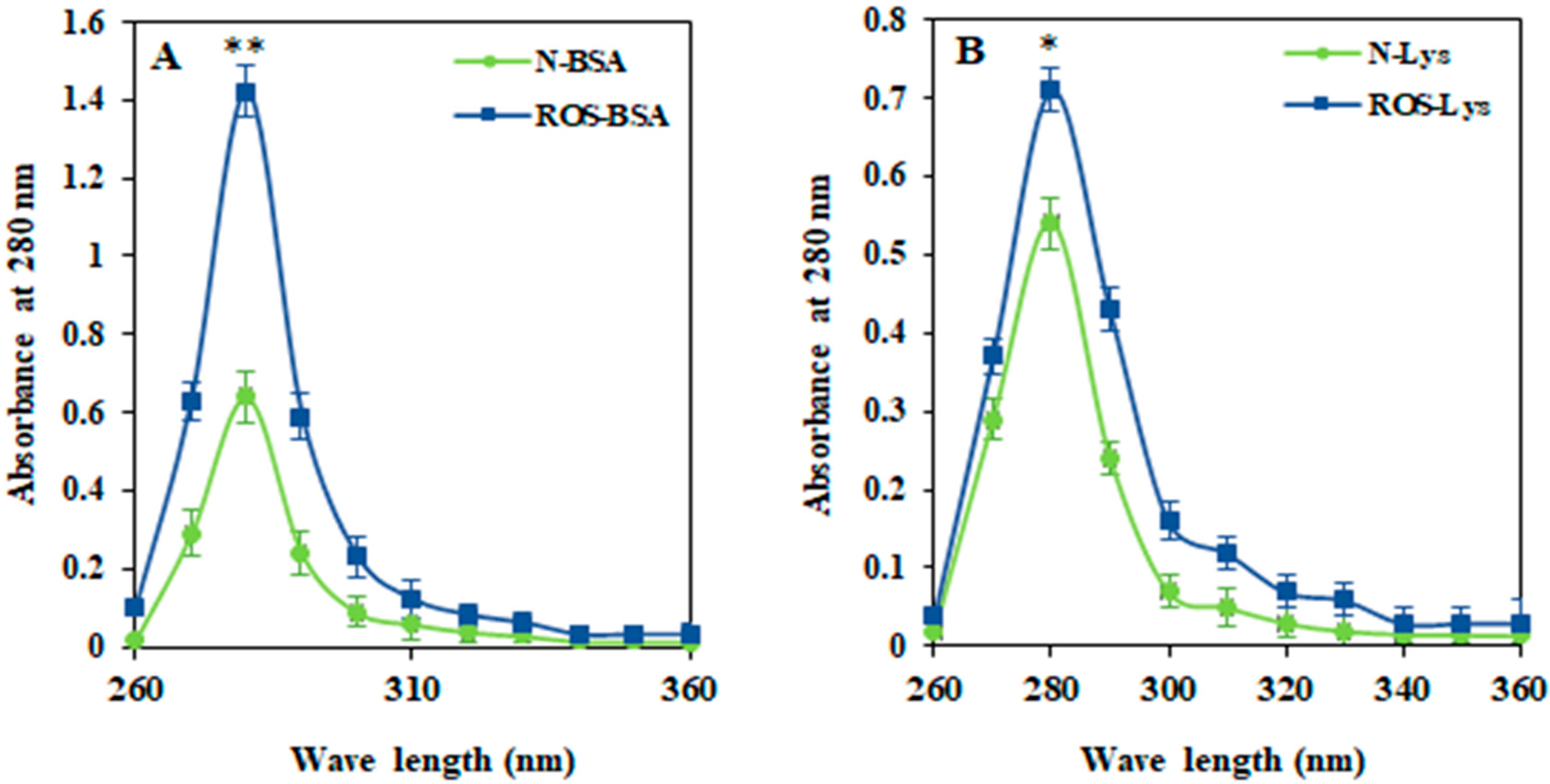
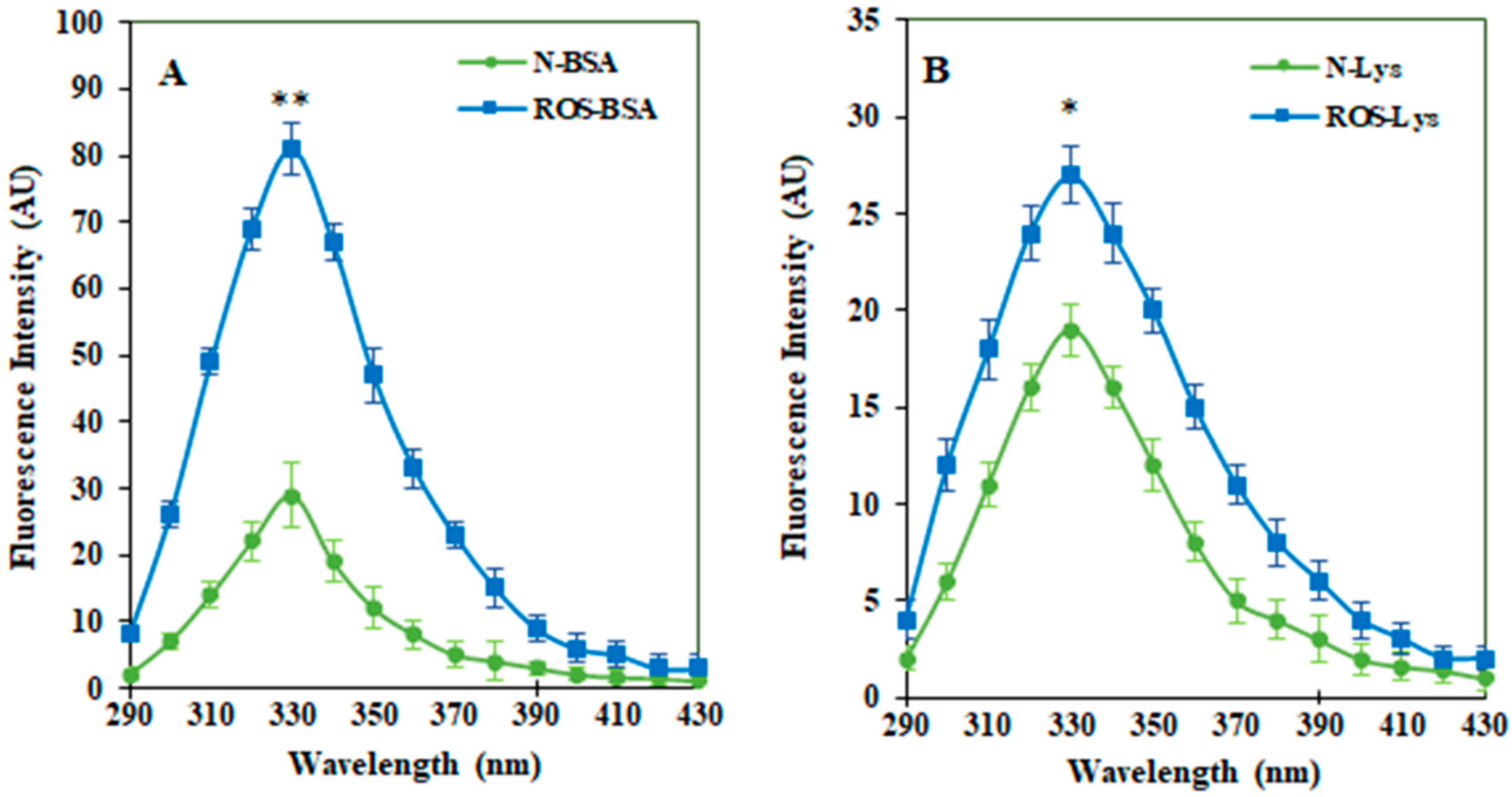
3.1. Characterization of ROS-HSA by Spectroscopic Studies
3.2. Biochemical and Immunological Analysis in Serum Samples
Proinflammatory Cytokines in Serum Samples
3.3. Serum Carbonyl Content
3.4. Serum MDA Levels
3.5. Clinical Characterization Analyses
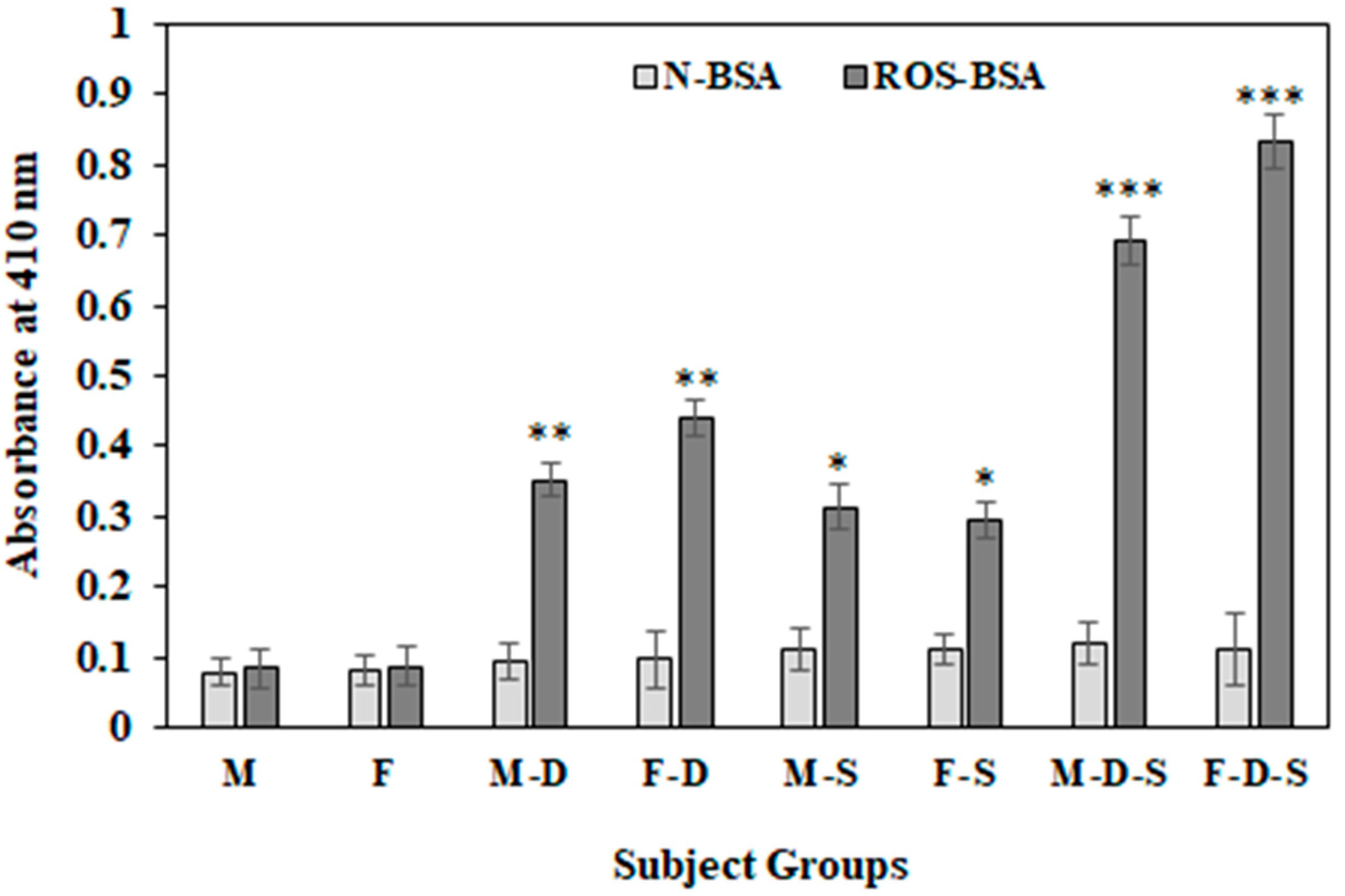
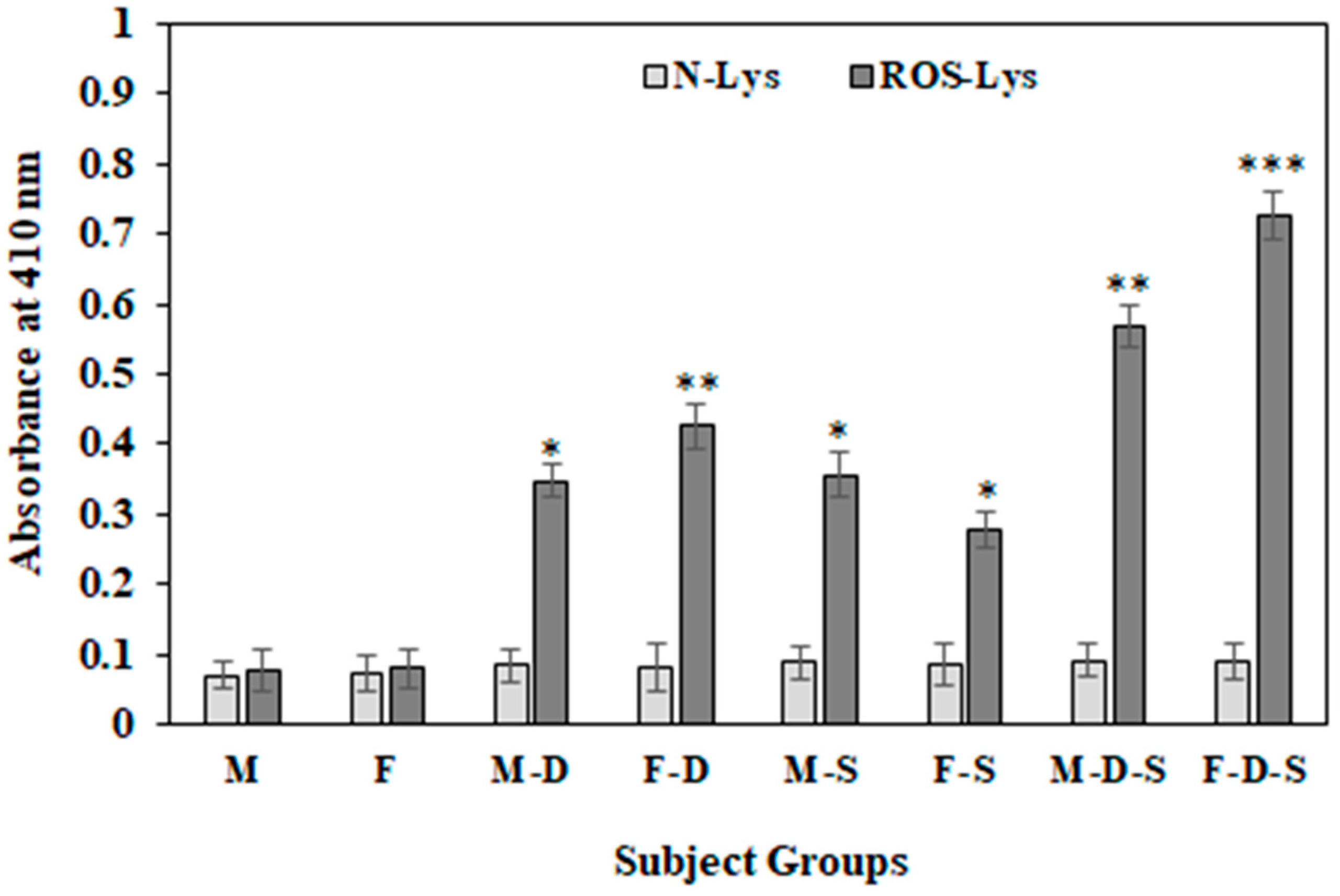
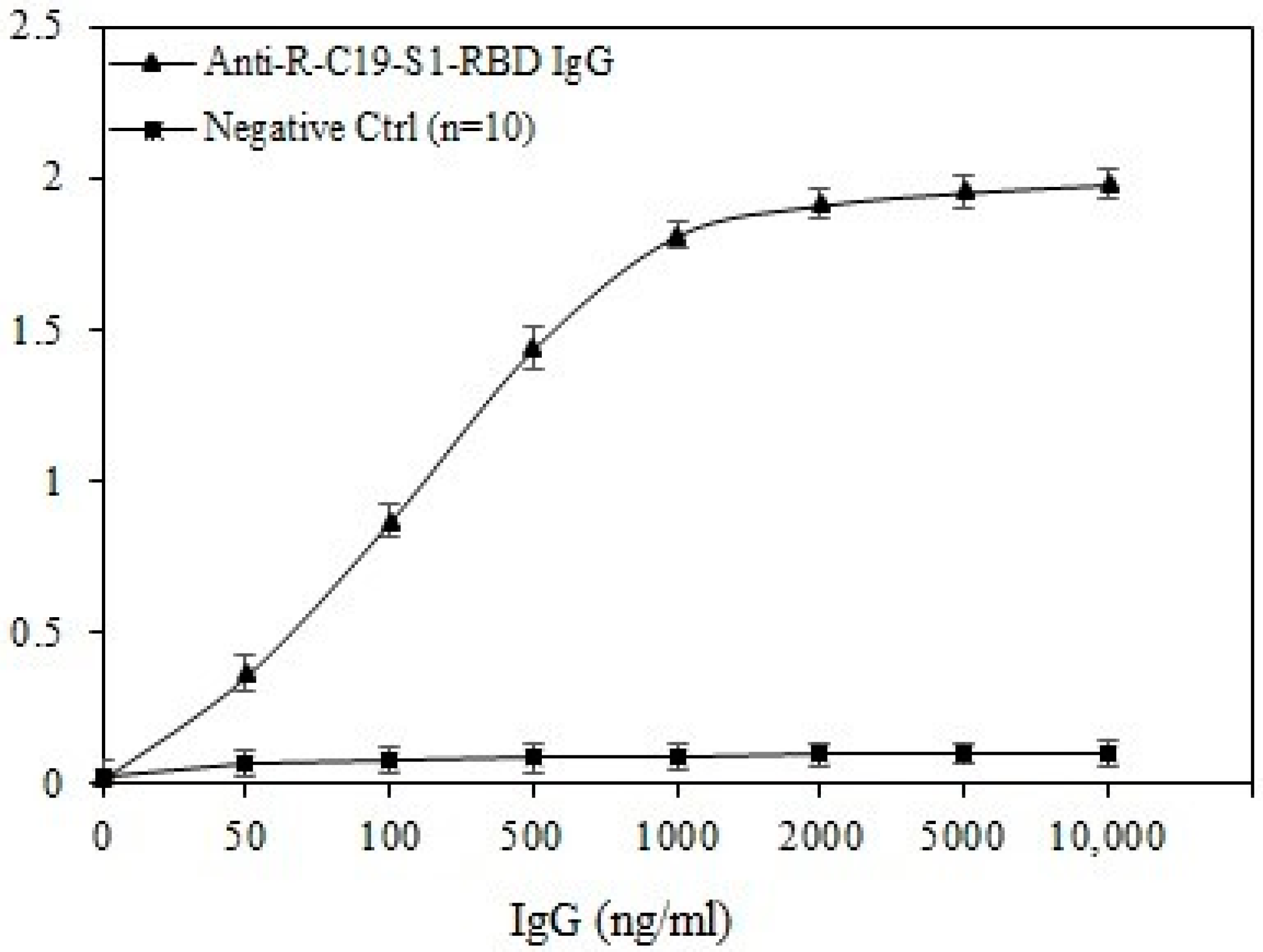
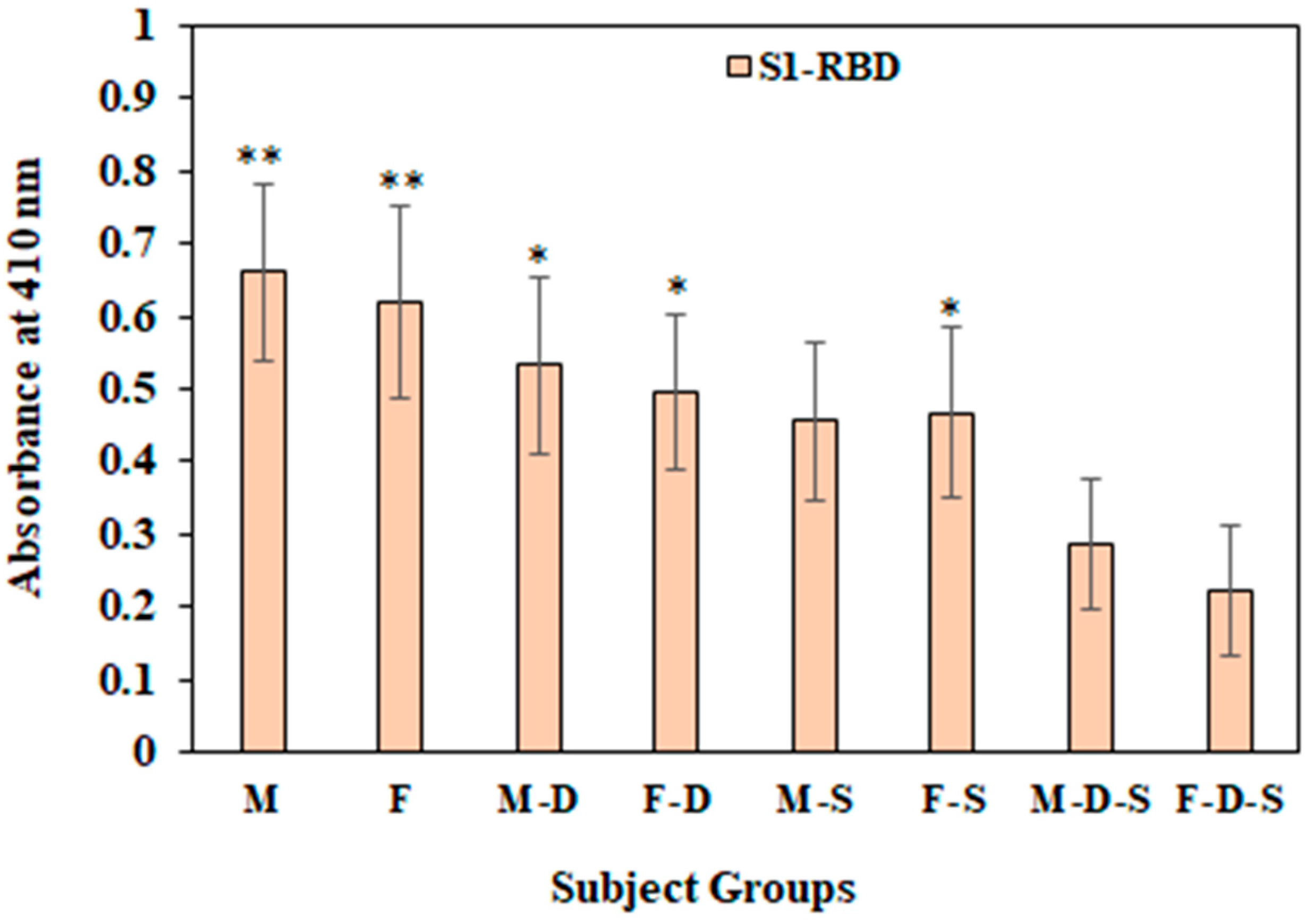
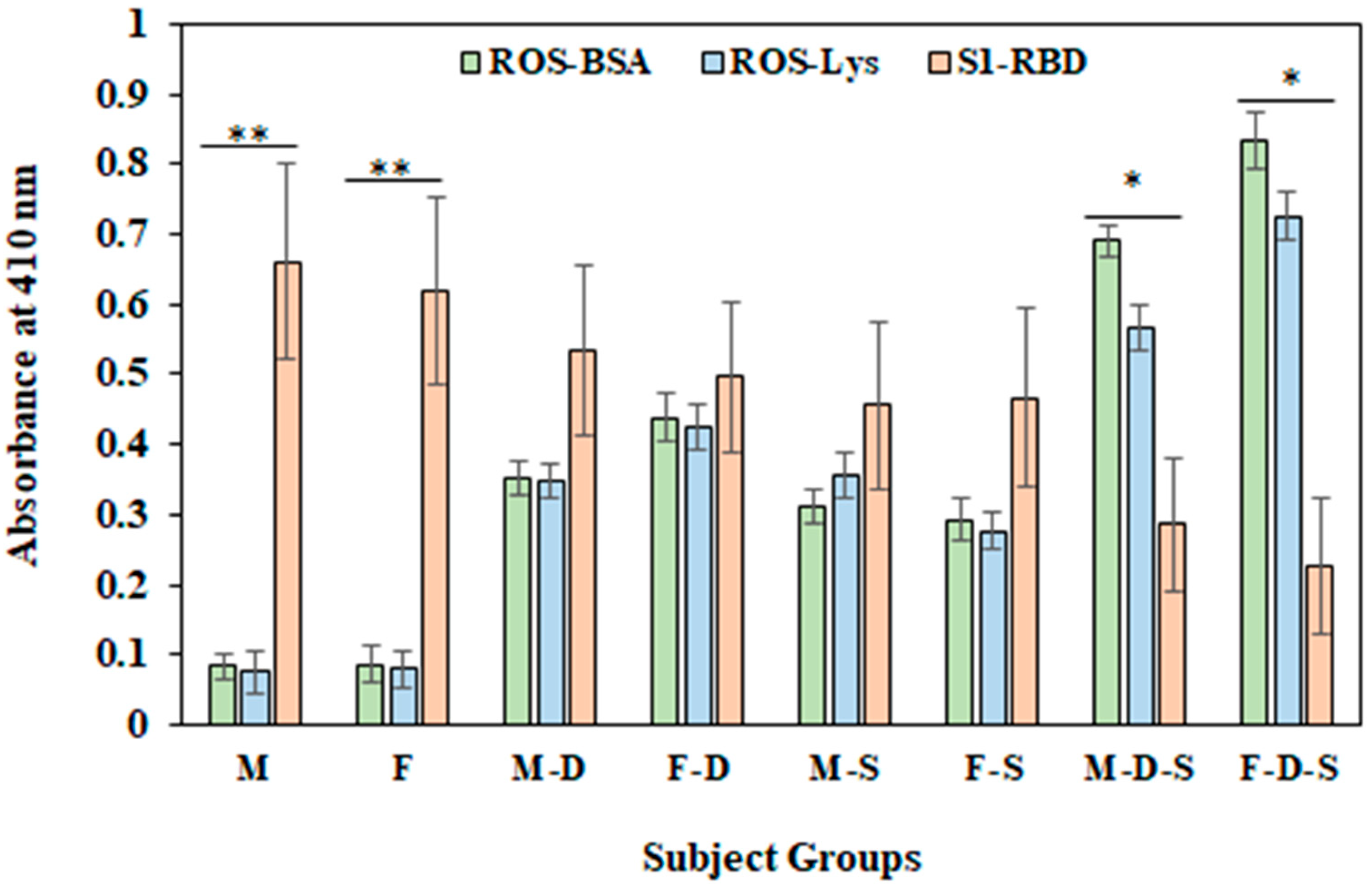
3.6. Direct Binding ELISA
3.7. Inhibition ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. China Novel Coronavirus Investigating and Research Team. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta BioMed. 2020, 91, 157–160. [Google Scholar]
- The Impact of COVID-19 on Mental Health Cannot Be Made Light of. WHO. Available online: https://www.who.int/news-room/feature-stories/detail/the-impact-of-covid-19-on-mental-health-cannot-be-made-light-of (accessed on 17 September 2022).
- Sherwani, S.; Khan, M.W.A. Cytokine Response in SARS-CoV-2 Infection in the Elderly. J. Inflamm. Res. 2020, 13, 737–747. [Google Scholar] [CrossRef]
- Chvatal-Medina, M.; Mendez-Cortina, Y.; Patino, P.J.; Velilla, P.A.; Rugeles, M.T. Antibody responses in Covid-19: A review. Front. Immunol. 2021, 12, 633184. [Google Scholar] [CrossRef]
- Kreer, C.; Zehner, M.; Weber, T.; Ercanoglu, M.S.; Gieselmann, L.; Rohde, C.; Halwe, S.; Korenkov, M.; Schommers, P.; Vanshylla, K.; et al. Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients. Cell 2020, 182, 843–854. [Google Scholar] [CrossRef]
- Addetia, A.; Crawford, K.H.; Dingens, A.; Zhu, H.; Roychoudhury, P.; Huang, M.L.; Jerome, K.R.; Bloom, J.D.; Greninger, A.L. Neutralizing Antibodies Correlate with Protection from SARS-CoV-2 in Humans during a Fishery Vessel Outbreak with a High Attack Rate. J. Clin. Microbiol. 2020, 58, e02107-20. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef]
- Chia, W.N.; Zhu, F.; Ong, S.W.X.; Young, B.E.; Fong, S.W.; Le Bert, N.; Tan, C.W.; Tiu, C.; Zhang, J.; Tan, S.Y.; et al. Dynamics of SARS-CoV-2 neutralising antibody responses and duration of immunity: A longitudinal study. Lancet Microbe 2021, 2, 240–249. [Google Scholar] [CrossRef]
- van der Werf, E.T.; Busch, M.; Jong, M.C.; Hoenders, H.J.R. Lifestyle changes during the first wave of the COVID-19 pandemic: A cross-sectional survey in the Netherlands. BMC Public Health 2021, 21, 1226. [Google Scholar] [CrossRef]
- Trabelsi, K.; Ammar, A.; Masmoudi, L.; Boukhris, O.; Chtourou, H.; Bouaziz, B.; Brach, M.; Bentlage, E.; How, D.; Ahmed, M.; et al. Globally altered sleep patterns and physical activity levels by confinement in 5056 individuals: ECLB COVID-19 international online survey. Biol. Sport 2021, 38, 495–506. [Google Scholar] [CrossRef]
- Goodwin, G.M. Depression and associated physical diseases and symptoms. Dialogues Clin. Neurosci. 2006, 8, 259–265. [Google Scholar] [CrossRef]
- Kwong, A.S.F.; López, J.; Hammerton, G.; Manley, D.; Timpson, N.; Leckie, G.; Pearson, R.M. Genetic and Environmental Risk Factors Associated With Trajectories of Depression Symptoms From Adolescence to Young Adulthood. JAMA Netw. Open 2019, 2, e196587. [Google Scholar] [CrossRef] [Green Version]
- Luppino, F.S.; de Wit, L.M.; Bouvy, P.F.; Stijnen, T.; Cuijpers, P.; Penninx, B.W.; Zitman, F.G. Overweight, obesity, and depression: A systematic review and meta-analysis of longitudinal studies. Arch. Gen. Psychiatry 2010, 67, 220–229. [Google Scholar] [CrossRef]
- Mezuk, B.; Eaton, W.W.; Albrecht, S.; Golden, S.H. Depression and type 2 diabetes over the lifespan: A meta-analysis. Diabetes Care 2008, 31, 2383–2390. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, A.; Kuper, H.; Hemingway, H. Depression as an aetiologic and prognostic factor in coronary heart disease: A meta-analysis of 6362 events among 146538 participants in 54 observational studies. Eur. Heart J. 2006, 27, 2763–2774. [Google Scholar] [CrossRef] [Green Version]
- Chida, Y.; Hamer, M.; Wardle, J.; Steptoe, A. Do stress-related psychosocial factors contribute to cancer incidence and survival? Nat. Clin. Pract. Oncol. 2008, 5, 466–475. [Google Scholar] [CrossRef]
- Bueno-Notivol, J.; Gracia-García, P.; Olaya, B.; Lasheras, I.; López-Antón, R.; Santabárbara, J. Prevalence of depression during the COVID-19 outbreak: A meta-analysis of community-based studies. Int. J. Clin. Health Psychol. 2021, 21, 100196. [Google Scholar] [CrossRef]
- Saraswathi, I.; Saikarthik, J.; Kumar, S.; Srinivasan, M.K.; Ardhanaari, M.; Gunapriya, R. Impact of COVID-19 outbreak on the mental health status of undergraduate medical students in a COVID-19 treating medical college: A prospective longitudinal study. Peer J. 2020, 8, e10164. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, A.J.; Pantaleón, Y.; Dios, I.; Falla, D. Fear of COVID-19, Stress, and Anxiety in University Undergraduate Students: A Predictive Model for Depression. Front. Psychol. 2020, 11, 591797. [Google Scholar] [CrossRef]
- Black, C.N.; Bot, M.; Scheffer, P.G.; Cuijpers, P.; Penninx, B.W. Is depression associated with increased oxidative stress? A systematic review and meta-analysis. Psychoneuroendocrinology 2015, 51, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Salim, S. Oxidative stress and psychological disorders. Curr. Neuropharmacol. 2014, 12, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Alouffi, S.; Sherwani, S.; Al-Mogbel, M.S.; Sherwani, M.K.A.; Khan, M.W.A. Depression and Smoking Augment the Production of Circulating Autoantibodies against Glycated HSA in Rheumatoid Arthritis Patients. Int. Arch. Allergy Immunol. 2018, 177, 170–180. [Google Scholar] [CrossRef]
- Khan, W.A.; Zaman, G.S.; Alouffi, S.; Khan, M.W.A. Depression and its related parameters increased the production of autoantibodies against 16α-hydroxyestrone-albumin complex in systemic lupus erythematosus. Int. Immunopharmacol. 2019, 71, 215–223. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Kamceva, G.; Arsova-Sarafinovska, Z.; Ruskovska, T.; Zdravkovska, M.; Kamceva-Panova, L.; Stikova, E. Cigarette Smoking and Oxidative Stress in Patients with Coronary Artery Disease. Open Access Maced J. Med. Sci. 2016, 4, 636–640. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.W.; Zhang, H.Z.; Liu, C.; Dong, K. Autoantibodies in COVID-19: Frequency and function. Autoimmun. Rev. 2021, 20, 102754. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, T.; Chen, J.; Hou, C.; Hua, L.; He, S.; Guo, Y.; Zhang, S.; Wang, Y.; Yuan, J.; et al. Clinical and Autoimmune Characteristics of Severe and Critical Cases of COVID-19. Clin. Transl. Sci. 2020, 13, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Vlachoyiannopoulos, P.G.; Magira, E.; Alexopoulos, H.; Jahaj, E.; Theophilopoulou, K.; Kotanidou, A.; Tzioufas, A.G. Autoantibodies related to systemic autoimmune rheumatic diseases in severely ill patients with COVID-19. Ann. Rheum. Dis. 2020, 79, 1661–1663. [Google Scholar] [CrossRef]
- Pascolini, S.; Vannini, A.; Deleonardi, G.; Ciordinik, M.; Sensoli, A.; Carletti, I.; Veronesi, L.; Ricci, C.; Pronesti, A.; Mazzanti, L.; et al. COVID-19 and Immunological Dysregulation: Can Autoantibodies be Useful? Clin. Transl. Sci. 2021, 14, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, X. Depressive Syndromes in Autoimmune Disorders of the Nervous System: Prevalence, Etiology, and Influence. Front. Psychiatry 2018, 9, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, W.A.; Malik, A.; Khan, M.W.A. Depression linked to higher antibodies production against estrogenized insulin in type 1 diabetes. Int. Immunopharmacol. 2020, 86, 106712. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L. The PHQ-9: A new depression and diagnostic severity measure. Psychiat. Ann. 2002, 32, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.W.A.; Sherwani, S.; Khan, W.A.; Moinuddin.; Ali, R. Characterization of hydroxyl radical modified GAD65: A potential autoantigen in type 1 diabetes. Autoimmunity 2009, 42, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.W.A.; Rasheed, Z.; Khan, W.A.; Ali, R. Biochemical, biophysical and thermodynamic analysis of in vitro glycated human serum albumin. Biochemistry 2007, 72, 146–152. [Google Scholar]
- Khan, M.W.; Al Otaibi, A.; Al-Zahrani, S.A.; Alshammari, E.M.; Haque, A.; Alouffi, S.; Khan, W.A.; Khan, S.N. Experimental and theoretical insight into resistance to glycation of bovine serum albumin. J. Mol. Struc. 2021, 1230, 129645. [Google Scholar] [CrossRef]
- Chernyak, B.V.; Popova, E.N.; Prikhodko, A.S.; Grebenchikov, O.A.; Zinovkina, L.A.; Zinovkin, R.A. COVID-19 and Oxidative Stress. Biochemistry 2020, 85, 1543–1553. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Levine, L.R.; Garland, D.; Oliver, N.C.; Amici, A.; Climent, I.; Lenz, A.G.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar]
- Khan, M.; Al Otaibi, A.; Sherwani, S.; Khan, W.; Alshammari, E.; Al-Zahrani, S.; Saleem, M.; Khan, S.; Alouffi, S. Glycation and oxidative stress increase autoantibodies in the elderly. Molecules 2020, 25, 3675. [Google Scholar] [CrossRef] [PubMed]
- Sherwani, S.; Khan, M.W.A.; Mallik, A.; Khan, M.; Saleem, M.; Raafat, M.; Shati, A.A.; Alam, N. Seroprevalence of Anti-S1-RBD Antibod;ies in Pre-pandemic and Pandemic Subjects From Hail Region, KSA. Front. Public Health 2022, 10, 874741. [Google Scholar] [CrossRef]
- Cai, Y.; Shi, S.; Yang, F.; Yi, B.; Chen, X.; Li, J.; Wen, Z. Fasting blood glucose level is a predictor of mortality in patients with COVID-19 independent of diabetes history. Diabetes Res. Clin. Pract. 2020, 169, 108437. [Google Scholar] [CrossRef] [PubMed]
- Khan, L.S.; McIntyre, R.S.; Rafalson, L.; Berdine, D.E.; Fox, C.H. Fasting Blood Glucose and Depressive Mood among Patients with Mental Illness in a Medicaid Managed Care Program. Depress. Res. Treat. 2011, 2011, 862708. [Google Scholar] [CrossRef] [PubMed]
- Merzon, E.; Green, I.; Shpigelman, M.; Vinker, S.; Raz, I.; Golan-Cohen, A.; Eldor, R. Haemoglobin A1c is a predictor of COVID-19 severity in patients with diabetes. Diabetes Metab. Res. Rev. 2021, 37, e3398. [Google Scholar] [CrossRef]
- Schmitz, N.; Deschênes, S.; Burns, R.; Smith, K. Depressive symptoms and glycated hemoglobin A1c: A reciprocal relationship in a prospective cohort study. Psychol. Med. 2016, 46, 945–955. [Google Scholar] [CrossRef]
- Dadras, O.; Alinaghi, S.A.S.; Karimi, A.; MohsseniPour, M.; Barzegary, A.; Vahedi, F.; Pashaei, Z.; Mirzapour, P.; Fakhfouri, A.; Zargari, G.; et al. Effects of COVID-19 prevention procedures on other common infections: A systematic review. Eur. J. Med. Res. 2021, 26, 67. [Google Scholar] [CrossRef]
- Dadras, O.; Afsahi, A.M.; Pashaei, Z.; Mojdeganlou, H.; Karimi, A.; Habibi, P.; Barzegary, A.; Fakhfouri, A.; Mirzapour, P.; Janfaza, N.; et al. The relationship between COVID-19 viral load and disease severity: A systematic review. Immun. Inflamm. Dis. 2022, 10, e580. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration. Coronavirus (COVID-19) Update: Daily Roundup 2 April 2020. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-daily-roundup-april-2-2020 (accessed on 22 October 2022).
- Mehraeen, E.; Dadras, O.; Afsahi, A.M.; Karimi, A.; Pour, M.M.; Mirzapour, P.; Barzegary, A.; Behnezhad, F.; Habibi, P.; Salehi, M.A.; et al. Vaccines for COVID-19: A Systematic Review of Feasibility and Effectiveness. Infect. Disord. Drug Targets 2022, 22, e230921196758. [Google Scholar] [CrossRef]
- Schlickeiser, S.; Schwarz, T.; Steiner, S.; Wittke, K.; Al Besher, N.; Meyer, O.; Kalus, U.; Pruß, A.; Kurth, F.; Zoller, T.; et al. Disease Severity, Fever, Age, and Sex Correlate With SARS-CoV-2 Neutralizing Antibody Responses. Front. Immunol. 2021, 11, 628971. [Google Scholar] [CrossRef]
- Salari, N.; Hosseinian-Far, A.; Jalali, R.; Vaisi-Raygani, A.; Rasoulpoor, S.; Mohammadi, M.; Rasoulpoor, S.; Khaledi-Paveh, B. Prevalence of stress, anxiety, depression among the general population during the COVID-19 pandemic: A systematic review and meta-analysis. Glob. Health 2020, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Mental Disorders Collaborators. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- Sherwani, S.; Rajendrasozhan, S.; Khan, M.W.A.; Saleem, M.; Khan, M.; Khan, S.; Raafat, M.; Othman Alqahtani, F. Pharmacological Profile of Nigella sativa Seeds in Combating COVID-19 through In-Vitro and Molecular Docking Studies. Processes 2022, 10, 1346. [Google Scholar] [CrossRef]
- NIDA Do People with Mental Illness and Substance Use Disorders Use Tobacco More Often? 12 April 2021. Available online: https://nida.nih.gov/publications/research-reports/tobacco-nicotine-e-cigarettes/do-people-mental-illness-substance-use-disorders-use-tobacco-more-often (accessed on 23 October 2022).
- Xiao, M.M.; Li, J.B.; Jiang, L.L.; Shao, H.; Wang, B.L. Plasma nesfatin-1 level is associated with severity of depression in Chinese depressive patients. BMC Psychiatry 2018, 18, 88. [Google Scholar] [CrossRef]
- Metsios, G.S.; Stavropoulos-Kalinoglou, A.; Nevill, A.M.; Douglas, K.M.; Koutedakis, Y.; Kitas, G.D. Cigarette smoking significantly increases basal metabolic rate in patients with rheumatoid arthritis. Ann. Rheum. Dis. 2008, 67, 70–73. [Google Scholar] [CrossRef] [Green Version]
- Lazzer, S.; Bedogni, G.; Lafortuna, C.; Marazzi, N.; Busti, C.; Galli, R.; De Col, A.; Agosti, F.; Sartorio, A. Relationship Between Basal Metabolic Rate, Gender, Age, and Body Composition in 8780 White Obese Subjects. Obesity 2009, 18, 71–78. [Google Scholar] [CrossRef]
- Lotrich, F.E. Inflammatory cytokine-associated depression. Brain Res. 2015, 1617, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Archer, J.A.; Hutchison, I.L.; Dorudi, S.; Stansfeld, S.A.; Korszun, A. Interrelationship of depression, stress and inflammation in cancer patients: A preliminary study. J. Affect. Disord. 2012, 143, 39–46. [Google Scholar] [CrossRef]
- Groer, M.W.; Morgan, M. Immune, health and endocrine characteristics of depressed postpartum mothers. Psychoneuroendocrinology 2007, 32, 133–139. [Google Scholar] [CrossRef]
- Bennett, B.K.; Hickie, I.B.; Quigley, B.; Brennan, C.M.; Vollmer-Conna, U.S.; Wakefield, D.; Douglas, M.P.; Hansen, G.R.; Tahmindjis, A.J.; Lloyd, A.R. The relationship between fatigue, psychological and immunological variables in acute infectious illness. Aust. N. Z. J. Psychiatry 1998, 32, 180–186. [Google Scholar] [CrossRef]
- Kahl, K.G.; Kruse, N.; Faller, H.; Weiss, H.; Rieckmann, P. Expression of tumor necrosis factor-a and interferon-g mRNA in blood cells correlates with depression scores during an acute attack in patients with multiple sclerosis. Psychoneuroendocrinology 2002, 27, 671–681. [Google Scholar] [CrossRef]
- Postal, M.; Pelicari, K.O.; Sinicato, N.A.; Marini, R.; Costallat, L.T.L.; Appenzeller, S. Th1/Th2 cytokine profile in childhood-onset systemic lupus erythematosus. Cytokine 2013, 61, 785–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, A.; Tang, C.S.; Ho, R.C. Serum tumour necrosis factor-alpha is associated with poor health-related quality of life and depressive symptoms in patients with systemic lupus erythematosus. Lupus 2013, 22, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Dieperink, E.; Willenbring, M.; Ho, S.B. Neuropsychiatric symptoms associated with hepatitis C and interferon alpha: A review. Am. J. Psychiatry 2000, 157, 867–876. [Google Scholar] [CrossRef]
- Pomova, N.I.; Ivanikov, I.O.; Siutkin, V.E. Use of peg-intron in combined treatment of chronic liver disease caused by HIV infection. Eksp. Klin. Gastroenterol. 2003, 1, 42–45. [Google Scholar]
- Salk, R.H.; Hyde, J.S.; Abramson, L.Y. Gender differences in depression in representative national samples: Meta-analyses of diagnoses and symptoms. Psychol. Bull. 2017, 143, 783–822. [Google Scholar] [CrossRef] [PubMed]
- Wharton, W.; Gleason, C.E.; Olson, S.R.; Carlsson, C.M.; Asthana, S. Neurobiological underpinnings of the estrogen-mood relationship. Curr. Psychiatry Rev. 2012, 8, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Scapagnini, G.; Davinelli, S.; Drago, F.; de Lorenzo, A.; Oriani, G. Antioxidants as antidepressants: Fact or fiction? CNS Drugs 2012, 26, 477–490. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Bouayed, J.; Rammal, H.; Soulimani, R. Oxidative stress and anxiety relationship and cellular pathways. Oxid. Med. Cell Longev. 2009, 2, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Hovatta, I.; Juhila, J.; Donner, J. Oxidative stress in anxiety and comorbid disorders. Neurosci. Res. 2010, 68, 261–275. [Google Scholar] [CrossRef]
- Padmavathi, P.; Raghu, P.S.; Reddy, V.D.; Bulle, S.; Marthadu, S.B.; Maturu, P.; Varadacharyulu, N. Chronic cigarette smoking-induced oxidative/nitrosative stress in human erythrocytes and platelets. Mol. Cell. Toxicol. 2018, 14, 27–34. [Google Scholar] [CrossRef]
- Newton, A.H.; Cardani, A.; Braciale, T.J. The host immune response in respiratory virus infection: Balancing virus clearance and immunopathology. Semin. Immunopathol. 2016, 38, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A.; Pirofski, L.A. The convalescent sera option for containing COVID-19. J. Clin. Investig. 2020, 130, 1545–1548. [Google Scholar] [CrossRef] [Green Version]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc. Natl. Acad. Sci. USA 2020, 117, 9490–9496. [Google Scholar] [CrossRef]
- Langhi, D.M.; Santis, G.C.; Bordin, J.O. COVID-19 convalescent plasma transfusion. Hematol. Transfus. Cell Ther. 2020, 42, 113–115. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Redox Homeostasis and Immune Alterations in Coronavirus Disease-19. Biology 2022, 11, 159. [Google Scholar] [CrossRef]
- Pincemail, J.; Cavalier, E.; Charlier, C.; Cheramy–Bien, J.-P.; Brevers, E.; Courtois, A.; Fadeur, M.; Meziane, S.; Goff, C.; Misset, B.; et al. Oxidative Stress Status in COVID-19 Patients Hospitalized in Intensive Care Unit for Severe Pneumonia. A Pilot Study. Antioxidants 2021, 10, 257. [Google Scholar] [CrossRef]
- Frasca, D.; Reidy, L.; Romero, M.; Diaz, A.; Cray, C.; Kahl, K.; Blomberg, B.B. The majority of SARS-CoV-2-specific antibodies in COVID-19 patients with obesity are autoimmune and not neutralizing. Int. J. Obes. 2022, 46, 427–432. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups (21–60 Years) n = 15 | Age (Years ± SD) | BMR (cal/sq.m/hr) | FBG (mg/dL) | HbA1C (%) | IFN-γ (pg/mL) | TNF-α (pg/mL) | Smoking Duration (Years ± SD) | MDA Content nmol/mL | Carbonyl Content (nmol/mg Protein) |
|---|---|---|---|---|---|---|---|---|---|
| M | 38 ± 9.3 | 36.3 ± 3.4 | 87.9 ± 8.1 | 5.4 ± 0.3 | 4.6 ± 0.41 | 0.95 ± 0.15 | ― | 0.77 ± 0.17 | 0.71 ± 0.08 |
| F | 40 ± 10.8 | 32.1 ± 3.1 | 88.6 ± 9.5 | 5.5 ± 0.3 | 4.5 ± 0.43 | 0.96 ± 0.14 | ― | 0.79 ± 0.16 | 0.68 ± 0.07 |
| M-D | 39 ± 8.5 | 37.5 ± 3.9 | 90.5 ± 7.5 | 5.6 ± 0.3 | 5.3 ± 0.48 * | 1.06 ± 0.11 * | ― | 1.21 ± 0.21 * | 0.88 ± 0.13 * |
| F-D | 37 ± 11.1 | 32.8 ± 4.1 | 91.1 ± 6.6 | 5.5 ± 0.4 | 5.6 ± 0.49 * | 1.16 ± 0.18 * | ― | 1.47 ± 0.27 * | 1.13 ± 0.17 ** |
| M-S | 41 ± 10.2 | 41.4 ± 3.8 * | 88.7 ± 7.8 | 5.4 ± 0.4 | 4.8 ± 0.51 | 1.0 ± 0.16 | 19.2 ± 9.2 * | 0.87 ± 0.16 | 0.99 ± 0.16 * |
| F-S | 38 ± 12.4 | 35.8 ± 3.7 * | 89.4 ± 8.3 | 5.4 ± 0.3 | 4.7 ± 0.52 | 1.03 ± 0.12 | 8.5 ± 4.1 | 0.84 ± 0.19 | 0.89 ± 0.11 * |
| M-D-S | 41 ± 11.7 | 42.8 ± 4.4 * | 93.8 ± 7.7 | 5.6 ± 0.3 | 6.6 ± 0.65 *** | 1.29 ± 0.19 ** | 18.9 ± 5.4 * | 1.81 ± 0.31 ** | 2.11 ± 0.34 *** |
| F-D-S | 40 ± 9.9 | 36.9 ± 3.9 * | 94.1 ± 6.9 * | 5.7 ± 0.3 | 7.4 ± 0.73 *** | 1.37 ± 0.18 ** | 12.9 ± 6.3 | 2.19 ± 0.42 *** | 2.42 ± 0.38 *** |
| Groups | Anti-ROS-BSA-Ab | Anti-ROS-Lys-Ab | Anti-S1-RBD-Ab |
|---|---|---|---|
| M | 6.1 ± 1.4 | 7.7 ± 0.8 | 10 * (86.9 ± 1.7) |
| F | 6.6 ± 1.6 | 7.9 ± 1.0 | 10 * (87.9 ± 1.9) |
| M-D | 10 * (38.7 ± 3.3) | 9 * (35.8 ± 3.9) | 11 * (80.8 ± 3.9) |
| F-D | 10 * (51.5 ± 4.2) | 10 * (46.1 ± 4.5) | 10 * (73.1 ± 4.5) |
| M-S | 11 * (40.3 ± 4.1) | 10 * (37.5 ± 3.8) | 11 * (81.5 ± 3.8) |
| F-S | 9 * (35.3 ± 3.3) | 8 * (33.1 ± 3.0) | 12 * (83.1 ± 3.0) |
| M-D-S | 11 * (59.1 ± 5.5) | 10 * (57.3 ± 5.3) | 10 * (68.3 ± 5.3) |
| F-D-S | 12 * (77.7 ± 6.2) | 12 * (73.8 ± 6.0) | 9 * (59.8 ± 6.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherwani, S.; Raafat, M.; Rajendrasozhan, S.; Khan, M.; Saleem, M.; Husain, Q.; Khan, S.; Alam, N.; Khan, M.W.A. Increased Levels of Autoantibodies against ROS-Modified Proteins in Depressed Individuals with Decrease in Antibodies against SARS-CoV-2 Antigen (S1-RBD). Curr. Issues Mol. Biol. 2022, 44, 5260-5276. https://doi.org/10.3390/cimb44110358
Sherwani S, Raafat M, Rajendrasozhan S, Khan M, Saleem M, Husain Q, Khan S, Alam N, Khan MWA. Increased Levels of Autoantibodies against ROS-Modified Proteins in Depressed Individuals with Decrease in Antibodies against SARS-CoV-2 Antigen (S1-RBD). Current Issues in Molecular Biology. 2022; 44(11):5260-5276. https://doi.org/10.3390/cimb44110358
Chicago/Turabian StyleSherwani, Subuhi, Mohamed Raafat, Saravanan Rajendrasozhan, Mahvish Khan, Mohd Saleem, Qayyum Husain, Saif Khan, Noor Alam, and Mohd Wajid Ali Khan. 2022. "Increased Levels of Autoantibodies against ROS-Modified Proteins in Depressed Individuals with Decrease in Antibodies against SARS-CoV-2 Antigen (S1-RBD)" Current Issues in Molecular Biology 44, no. 11: 5260-5276. https://doi.org/10.3390/cimb44110358
APA StyleSherwani, S., Raafat, M., Rajendrasozhan, S., Khan, M., Saleem, M., Husain, Q., Khan, S., Alam, N., & Khan, M. W. A. (2022). Increased Levels of Autoantibodies against ROS-Modified Proteins in Depressed Individuals with Decrease in Antibodies against SARS-CoV-2 Antigen (S1-RBD). Current Issues in Molecular Biology, 44(11), 5260-5276. https://doi.org/10.3390/cimb44110358