

Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia obovata, a Typical Mangrove Plant

Abstract
:1. Introduction
2. Materials and Methods
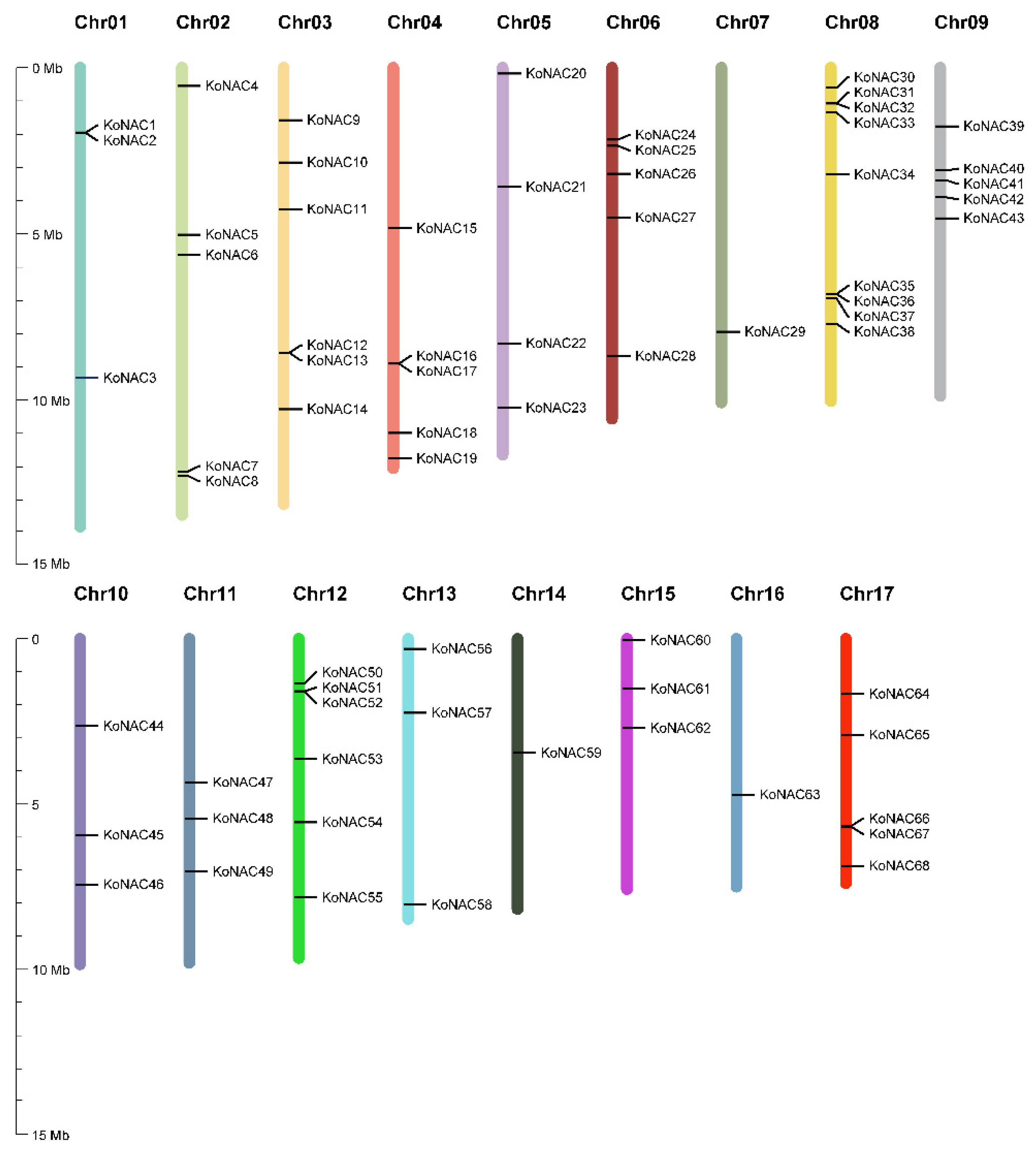
2.1. Identification and Chromosomal Distribution of NAC TFs in K. obovata
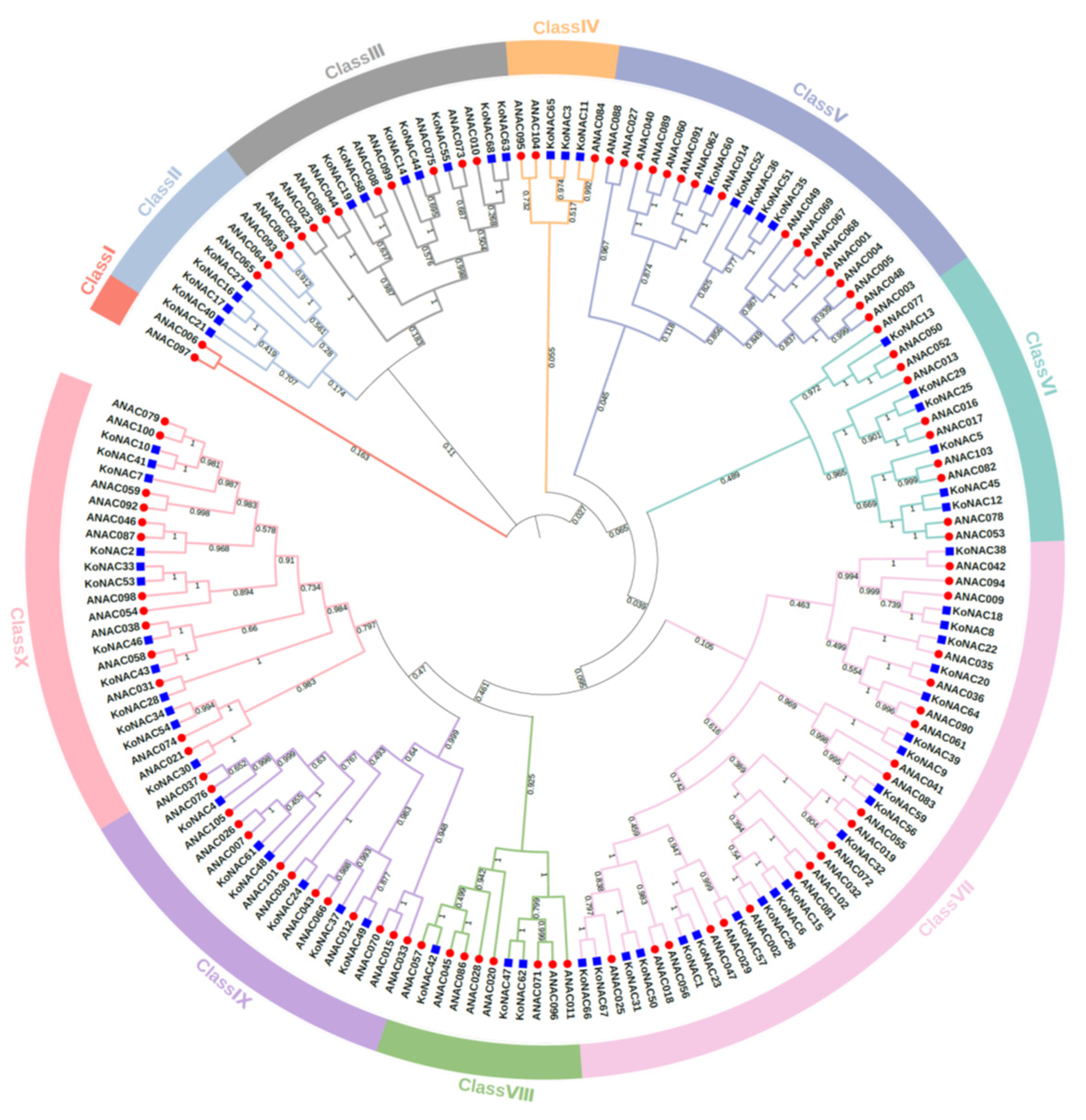
2.2. Phylogenetic Analysis of NAC Proteins
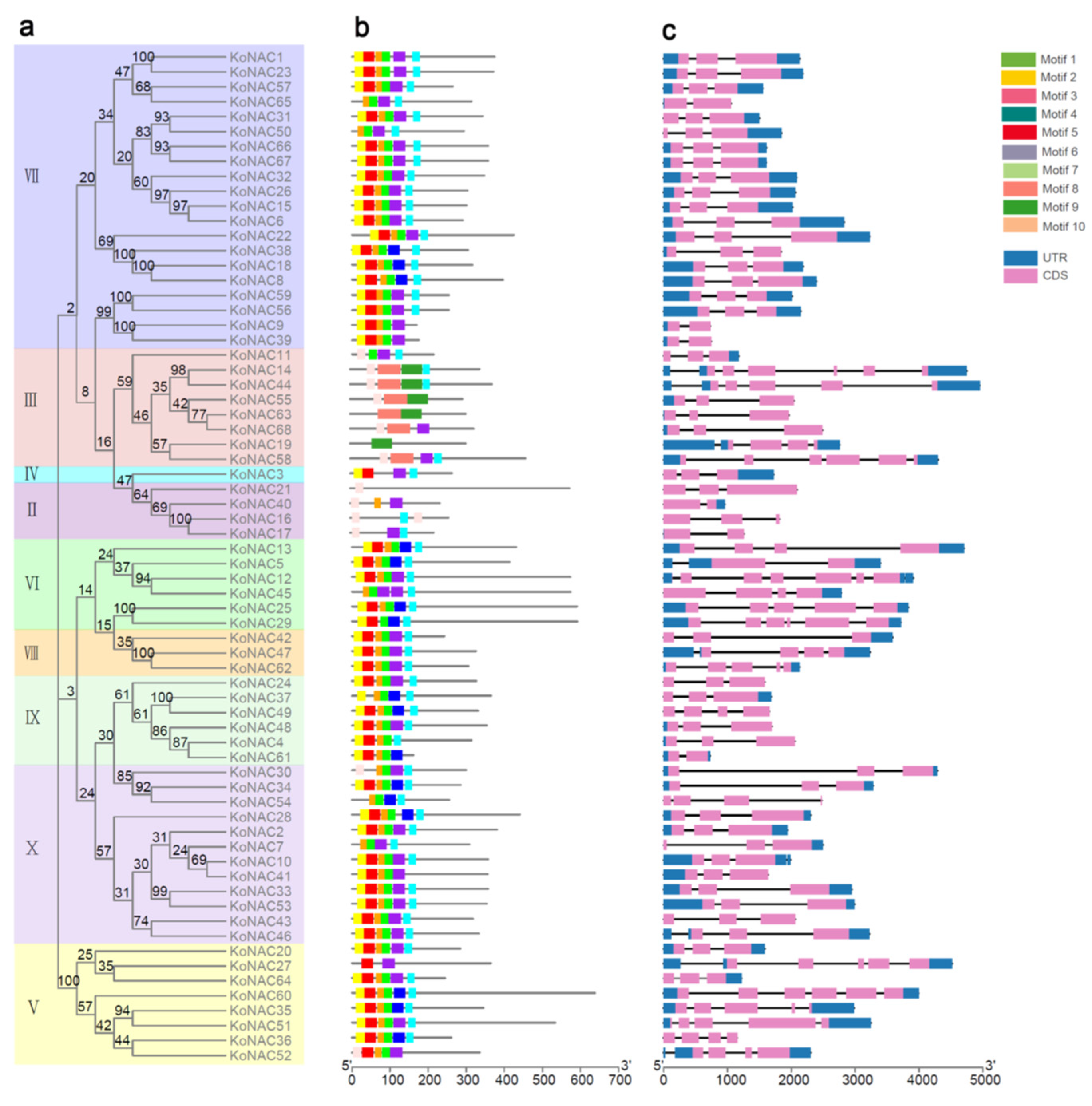
2.3. Gene Structure, Motif Identification, and Collinearity Analysis
2.4. Expression Analysis of KoNAC Genes Based on Public RNA-Seq Data
2.5. Plant Materials and Treatment
2.6. Cis-Regulatory Element Analysis of the KoNAC Genes
2.7. Quantitative Real-Time PCR Assays
3. Results
3.1. Genome-Wide Identification of the K. obovata NAC Genes
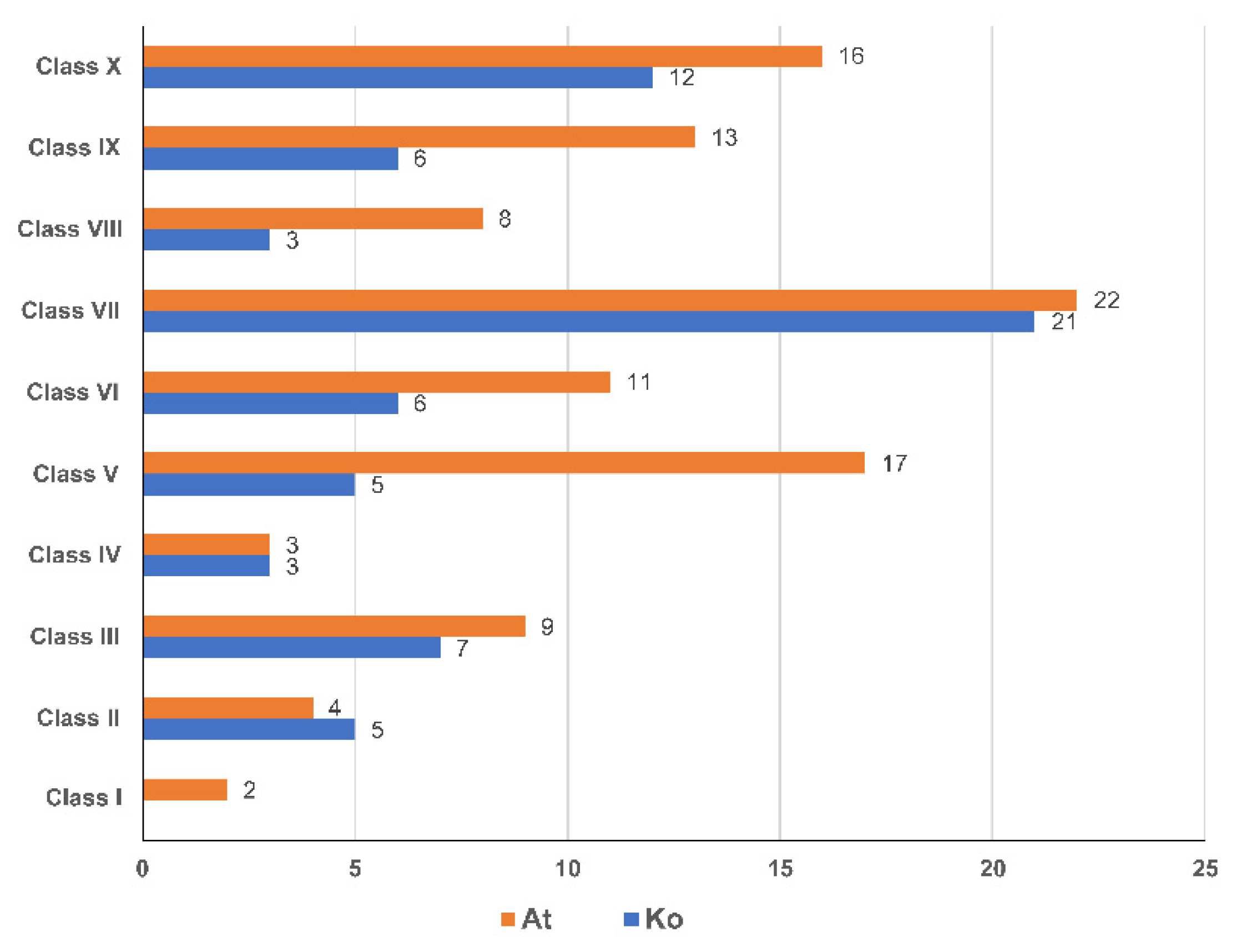
3.2. Phylogenetic Analysis and Classification of KoNAC Proteins
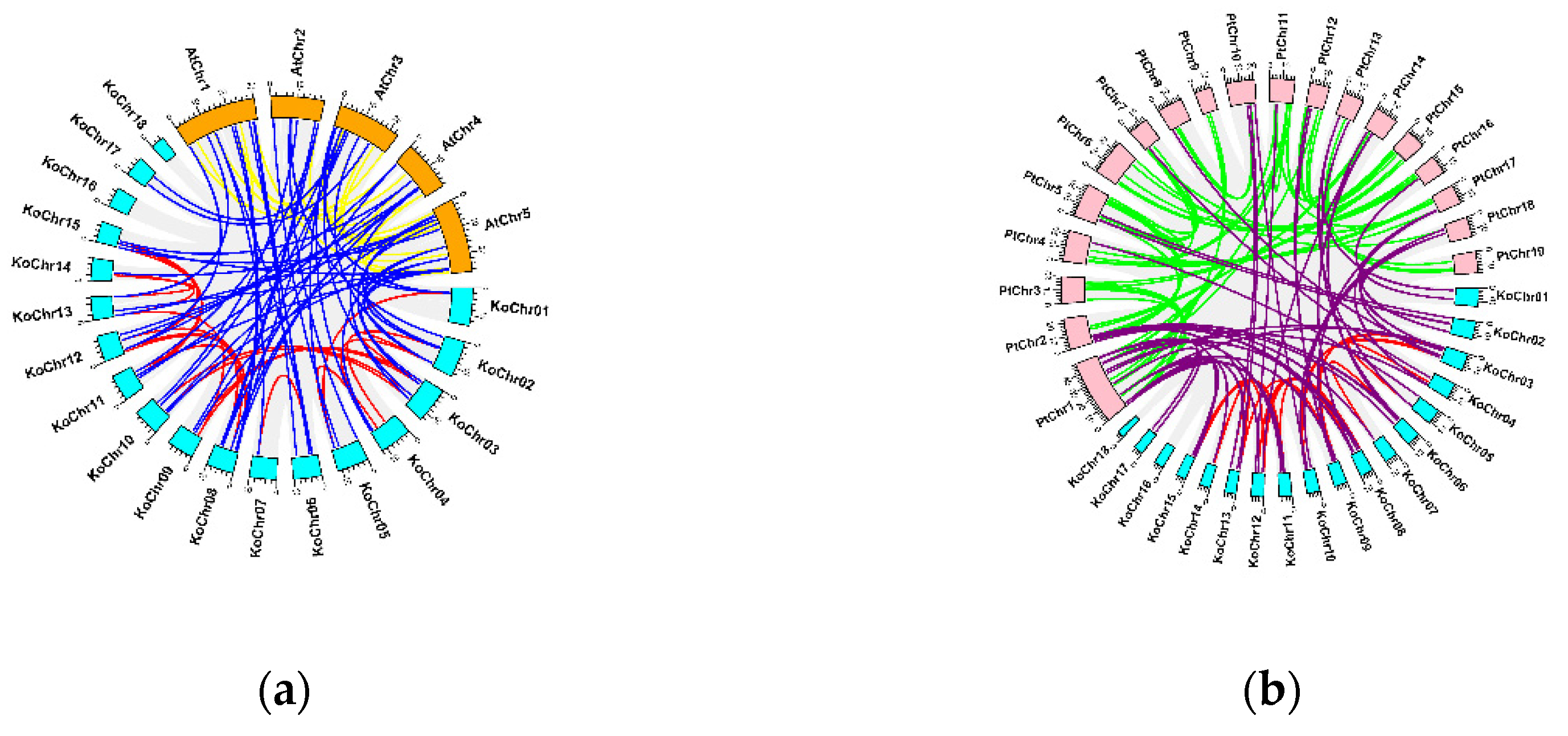
3.3. Collinearity Analysis of KoNAC Genes
3.4. Expression Patterns of KoNAC Genes in Different Organs
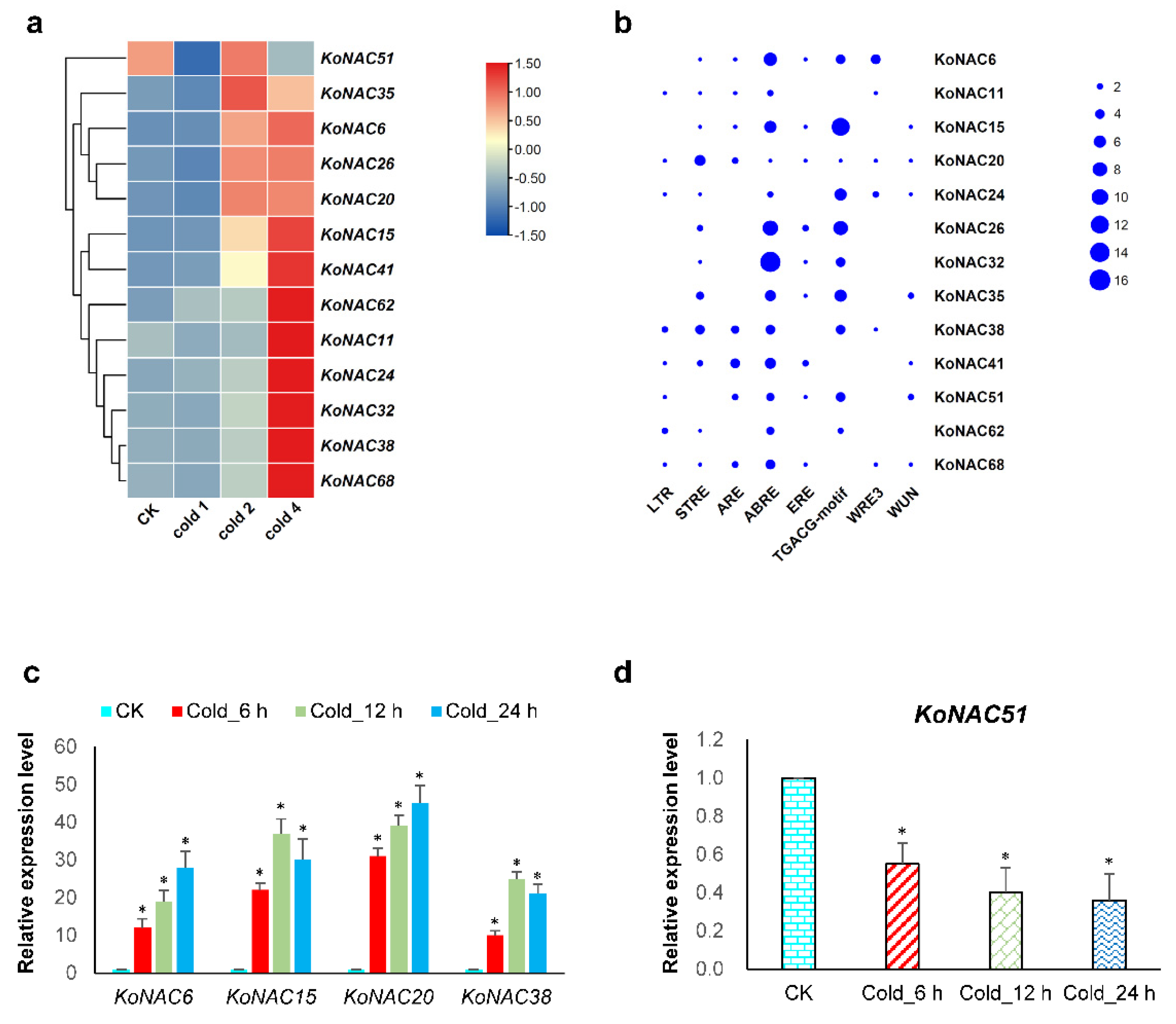
3.5. Expression Analysis of KoNAC Genes under Cold Treatment
3.6. Stress-Related Cis-Regulatory Elements Identified in KoNAC Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, K.; Foley, R.C.; Onate-Sanchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the Cup-Shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Jiang, Z.; Fu, H.; Chen, L.; Liao, G.; He, Y.; Huang, C.; Xu, X. Genome-wide identification and comprehensive analysis of NAC family genes involved in fruit development in kiwifruit (Actinidia). BMC Plant Biol. 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, J.; Zhang, Q.; Dong, A.; Wu, Q.; Zhu, X.; Zhu, X. Genome-wide identification and analysis of the NAC transcription factor gene family in garden Asparagus (Asparagus officinalis). Genes 2022, 13, 976. [Google Scholar] [CrossRef]
- Chen, S.; Lin, X.; Zhang, D.; Li, Q.; Zhao, X.; Chen, S. Genome-wide analysis of nac gene family in Betula pendula. Forests 2019, 10, 741. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Xie, F.; Liang, W.; Liang, Y.; Zhang, Z.; Zhao, J.; Hu, G.; Qin, Y. HuNAC20 and HuNAC25, two novel NAC genes from pitaya, confer cold tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 2189. [Google Scholar] [CrossRef]
- Li, X.; Cai, K.; Pei, X.; Li, Y.; Hu, Y.; Meng, F.; Song, X.; Tigabu, M.; Ding, C.; Zhao, X. Genome-wide identification of NAC transcription factor family in Juglans mandshurica and their expression analysis during the fruit development and ripening. Int. J. Mol. Sci. 2021, 22, 12414. [Google Scholar] [CrossRef]
- He, F.; Zhang, L.; Zhao, G.; Kang, J.; Long, R.; Li, M.; Yang, Q.; Chen, L. Genome-wide identification and expression analysis of the NAC gene family in Alfalfa revealed its potential roles in response to multiple abiotic stresses. Int. J. Mol. Sci. 2022, 23, 10015. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Yang, Z.; He, J.; Liu, A.; Chen, J.; Wang, S.; Wang, X.; Feng, G.; Li, D.; Peng, Y.; et al. Genome-wide investigation of the NAC transcription factor family in Miscanthus sinensis and expression analysis under various abiotic stress. Front. Plant Sci. 2021, 12, 766550. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fan, R.; Yang, Q.; Hu, C.; Sheng, O.; Deng, G.; Dong, T.; Li, C.; Peng, X.; Bi, F.; et al. Genome-wide identification and characterization of the NAC transcription factor family in Musa Acuminata and expression analysis during fruit ripening. Int. J. Mol. Sci. 2020, 21, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Fan, L.; Yu, X.; Zhang, X.; Hao, P.; Wei, D.; Zhang, G. Analysis of the NAC gene family in Salix and the identification of SpsNAC005 gene contributing to salt and drought tolerance. Forests 2022, 13, 971. [Google Scholar] [CrossRef]
- Jin, J.F.; Wang, Z.Q.; He, Q.Y.; Wang, J.Y.; Li, P.F.; Xu, J.M.; Zheng, S.J.; Fan, W.; Yang, J.L. Genome-wide identification and expression analysis of the NAC transcription factor family in tomato (Solanum lycopersicum) during aluminum stress. BMC Genom. 2020, 21, 288. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Ma, L.; Chen, X.; Fei, X.; He, B.; Luo, Y.; Liu, Y.; Wei, A. Genome-wide identification of the NAC gene family in Zanthoxylum bungeanum and their transcriptional responses to drought stress. Int. J. Mol. Sci. 2022, 23, 4769. [Google Scholar] [CrossRef]
- Peng, X.; Zhao, Y.; Li, X.; Wu, M.; Chai, W.; Sheng, L.; Wang, Y.; Dong, Q.; Jiang, H.; Cheng, B. Genome-wide identification, classification and analysis of NAC type gene family in maize. J. Genet. 2015, 94, 377–390. [Google Scholar] [CrossRef]
- Diao, P.; Chen, C.; Zhang, Y.; Meng, Q.; Lv, W.; Ma, N. The role of NAC transcription factor in plant cold response. Plant Signal. Behav. 2020, 15, 1785668. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Shan, W.; Kuang, J.F.; Lu, W.J.; Chen, J.Y. Banana fruit NAC transcription factor MaNAC1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1. Plant Cell Environ. 2014, 37, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Du, M.; Zhou, Z.; Wang, S.; Li, T.; Han, J.; Xu, T.; Yang, G. Overexpression of a Malus baccata NAC transcription factor gene MbNAC25 increases cold and salinity tolerance in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Tang, M.; Huang, Z.; Song, J.; Xu, J.; Ahammed, G.J.; Yu, J.; Zhou, Y. The miR164a-NAM3 module confers cold tolerance by inducing ethylene production in tomato. Plant J. 2022, 111, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Y.; Hu, H.; Chen, L.; Zhang, H.; Chen, R. CabHLH79 acts upstream of CaNAC035 to regulate cold stress in pepper. Int. J. Mol. Sci. 2022, 23, 2537. [Google Scholar] [CrossRef]
- Hou, X.M.; Zhang, H.F.; Liu, S.Y.; Wang, X.K.; Zhang, Y.M.; Meng, Y.C.; Luo, D.; Chen, R.G. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- An, J.P.; Li, R.; Qu, F.J.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor negatively regulates cold tolerance via CBF-dependent pathway. J. Plant Physiol. 2018, 221, 74–80. [Google Scholar] [CrossRef]
- Song, C.; Wu, M.; Zhou, Y.; Gong, Z.; Yu, W.; Zhang, Y.; Yang, Z. NAC-mediated membrane lipid remodeling negatively regulates fruit cold tolerance. Hortic. Res. 2022, 9, uhac039. [Google Scholar] [CrossRef]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How mangrove forests adjust to rising sea level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef] [Green Version]
- Nizam, A.; Meera, S.P.; Kumar, A. Genetic and molecular mechanisms underlying mangrove adaptations to intertidal environments. iScience 2022, 25, 103547. [Google Scholar] [CrossRef]
- Guo, Z.; Ma, D.; Li, J.; Wei, M.; Zhang, L.; Zhou, L.; Zhou, X.; He, S.; Wang, L.; Shen, Y.; et al. Genome-wide identification and characterization of aquaporins in mangrove plant Kandelia obovata and its role in response to the intertidal environment. Plant Cell Environ. 2022, 45, 1698–1718. [Google Scholar] [CrossRef]
- Su, W.; Ye, C.; Zhang, Y.; Hao, S.; Li, Q.Q. Identification of putative key genes for coastal environments and cold adaptation in mangrove Kandelia obovata through transcriptome analysis. Sci. Total Environ. 2019, 681, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, W.; Li, Q.Q.; Zhang, Y.; Yang, S.; Osland, M.J.; Huang, J.; Peng, C. Mangrove species’ responses to winter air temperature extremes in China. Ecosphere 2017, 8, e01865. [Google Scholar] [CrossRef]
- Peng, Y.-L.; Wang, Y.-S.; Fei, J.; Sun, C.-C.; Cheng, H. Ecophysiological differences between three mangrove seedlings (Kandelia obovata, Aegiceras corniculatum, and Avicennia marina) exposed to chilling stress. Ecotoxicology 2015, 24, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-M.; Wang, Y.-S.; Su, B.-Y.; Zhou, Y.-Y.; Chang, L.-F.; Ma, X.-Y.; Li, X.-M. Ecophysiological responses of five mangrove species (Bruguiera gymnorrhiza, Rhizophora stylosa, Aegiceras corniculatum, Avicennia marina, and Kandelia obovata) to chilling stress. Front. Mar. Sci. 2022, 9, 846566. [Google Scholar] [CrossRef]
- Hu, M.J.; Sun, W.H.; Tsai, W.C.; Xiang, S.; Lai, X.K.; Chen, D.Q.; Liu, X.D.; Wang, Y.F.; Le, Y.X.; Chen, S.M.; et al. Chromosome-scale assembly of the Kandelia obovata genome. Hortic. Res. 2020, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Waack, S.; Keller, O.; Asper, R.; Brodag, T.; Damm, C.; Fricke, W.F.; Surovcik, K.; Meinicke, P.; Merkl, R. Score-based prediction of genomic islands in prokaryotic genomes using hidden Markov models. BMC Bioinform. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M.; et al. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; You, S.; Zhao, X.; Xiong, L.; Li, J. Genome-wide identification of WRKY genes and their responses to chilling stress in Kandelia obovata. Front. Genet. 2022, 13, 875316. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Liao, H.Z.; Zhu, M.M.; Cui, H.H.; Du, X.Y.; Tang, Y.; Chen, L.Q.; Ye, D.; Zhang, X.Q. MARIS plays important roles in Arabidopsis pollen tube and root hair growth. J. Integr. Plant Biol. 2016, 58, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Yue, X.; Zeng, H.; Zhu, J. The protein phosphatase RCF2 and its interacting partner NAC019 are critical for heat stress-responsive gene regulation and thermotolerance in Arabidopsis. Plant Cell 2014, 26, 438–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.Y.; Kim, Y.; Kim, S.Y.; Lee, J.S.; Ahn, J.H. Control of flowering time and cold response by a NAC-domain protein in Arabidopsis. PLoS ONE 2007, 2, e642. [Google Scholar] [CrossRef] [PubMed]
- Gabaldon, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, N.; Tang, R.; Chen, X.; Xu, Z.; Ren, Z.; Wang, L. Genome-wide identification and characterization of WOX genes in Cucumis sativus. Genome 2021, 64, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Xu, Y.P.; Zhang, X.R.; Cai, X.Z. Brassica napus genome possesses extraordinary high number of CAMTA genes and CAMTA3 contributes to PAMP triggered immunity and resistance to Sclerotinia sclerotiorum. Front. Plant Sci. 2016, 7, 581. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Sardesai, N.; Fujinuma, T.; Chan, C.W.; Veena; Gelvin, S. B. Constitutive expression exposes functional redundancy between the Arabidopsis histone H2A gene HTA1 and other H2A gene family members. Plant Cell 2006, 18, 1575–1589. [Google Scholar] [CrossRef] [Green Version]
- Mao, C.; He, J.; Liu, L.; Deng, Q.; Yao, X.; Liu, C.; Qiao, Y.; Li, P.; Ming, F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol. J. 2020, 18, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Hernandez, B.Y.; Nunez-Munoz, L.; Calderon-Perez, B.; Xoconostle-Cazares, B.; Ruiz-Medrano, R. The NAC transcription factor ANAC087 induces aerial rosette development and leaf senescence in Arabidopsis. Front. Plant Sci. 2022, 13, 818107. [Google Scholar] [CrossRef]
- Martín-Pizarro, C.; Vallarino, J.G.; Osorio, S.; Meco, V.; Urrutia, M.; Pillet, J.; Casañal, A.; Merchante, C.; Amaya, I.; Willmitzer, L.; et al. The NAC transcription factor FaRIF controls fruit ripening in strawberry. Plant Cell 2021, 33, 1574–1593. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Lee, K.-H.; Du, Q.; Yang, S.; Yuan, B.; Qi, L.; Wang, H. A membrane-associated NAC domain transcription factor XVP interacts with TDIF co-receptor and regulates vascular meristem activity. New Phytol. 2020, 226, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Daneva, A.; Salanenka, Y.; Van Durme, M.; Huysmans, M.; Lin, Z.; De Winter, F.; Vanneste, S.; Karimi, M.; Van de Velde, J.; et al. KIRA1 and ORESARA1 terminate flower receptivity by promoting cell death in the stigma of Arabidopsis. Nat. Plants 2018, 4, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Alshareef, N.O.; Otterbach, S.L.; Allu, A.D.; Woo, Y.H.; de Werk, T.; Kamranfar, I.; Mueller-Roeber, B.; Tester, M.; Balazadeh, S.; Schmockel, S.M. NAC transcription factors ATAF1 and ANAC055 affect the heat stress response in Arabidopsis. Sci. Rep. 2022, 12, 11264. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, J.; Wu, Y. Arabidopsis ATAF1 enhances the tolerance to salt stress and ABA in transgenic rice. J. Plant Res. 2016, 129, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Deng, Z.; Lai, J.; Zhang, Y.; Yang, C.; Yin, B.; Zhao, Q.; Zhang, L.; Li, Y.; Yang, C.; et al. Dual function of Arabidopsis ATAF1 in abiotic and biotic stress responses. Cell Res. 2009, 19, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Delessert, C.; Kazan, K.; Wilson, I.W.; Van Der Straeten, D.; Manners, J.; Dennis, E.S.; Dolferus, R. The transcription factor ATAF2 represses the expression of pathogenesis-related genes in Arabidopsis. Plant J. 2005, 43, 745–757. [Google Scholar] [CrossRef]
- Wang, X.; Culver, J.N. DNA binding specificity of ATAF2, a NAC domain transcription factor targeted for degradation by Tobacco mosaic virus. BMC Plant Biol. 2012, 12, 157. [Google Scholar] [CrossRef] [Green Version]
- Christianson, J.A.; Wilson, I.W.; Llewellyn, D.J.; Dennis, E.S. The low-oxygen-induced NAC domain transcription factor ANAC102 affects viability of Arabidopsis seeds following low-oxygen treatment. Plant Physiol. 2009, 149, 1724–1738. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, S.; Ksas, B.; Havaux, M. Decoding β-cyclocitral-mediated retrograde signaling reveals the role of a detoxification response in plant tolerance to photooxidative stress. Plant Cell 2018, 30, 2495–2511. [Google Scholar] [CrossRef]
- Yang, X.; Wang, X.; Ji, L.; Yi, Z.; Fu, C.; Ran, J.; Hu, R.; Zhou, G. Overexpression of a Miscanthus lutarioriparius NAC gene MlNAC5 confers enhanced drought and cold tolerance in Arabidopsis. Plant Cell Rep. 2015, 34, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, Q.; Jiang, H.; Li, C.B.; Zhai, Q.; Zhang, J.; Wu, X.; Sun, J.; Xie, Q.; Li, C. Role of the Arabidopsis thaliana NAC transcription factors ANAC019 and ANAC055 in regulating jasmonic acid-signaled defense responses. Cell Res. 2008, 18, 756–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Li, H.; Bu, Q.; Li, C. The RHA2a-interacting proteins ANAC019 and ANAC055 may play a dual role in regulating ABA response and jasmonate response. Plant Signal. Behav. 2009, 4, 464–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimian-Motlagh, S.; Ribone, P.A.; Thirumalaikumar, V.P.; Allu, A.D.; Chan, R.L.; Mueller-Roeber, B.; Balazadeh, S. JUNGBRUNNEN1 confers drought tolerance downstream of the HD-Zip I transcription factor AtHB13. Front. Plant Sci. 2017, 8, 2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Bulbul, S.; Piao, W.; Choi, G.; Paek, N.C. Arabidopsis EARLY FLOWERING3 increases salt tolerance by suppressing salt stress response pathways. Plant J. 2017, 92, 1106–1120. [Google Scholar] [CrossRef] [Green Version]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat. Plants 2016, 2, 16013. [Google Scholar] [CrossRef]
- Qian, W.; Xiao, B.; Wang, L.; Hao, X.; Yue, C.; Cao, H.; Wang, Y.; Li, N.; Yu, Y.; Zeng, J.; et al. CsINV5, a tea vacuolar invertase gene enhances cold tolerance in transgenic Arabidopsis. BMC Plant Biol. 2018, 18, 228. [Google Scholar] [CrossRef]
- Wu, C.; Zheng, C.; Ji, G.; Jiang, P. Synergistic effects of HSE and LTR elements from hsp70 gene promoter of Ulva prolifera (Ulvophyceae, Chlorophyta) upon temperature induction. J. Phycol. 2019, 55, 738–743. [Google Scholar] [CrossRef]
- Ebeed, H.T. Genome-wide analysis of polyamine biosynthesis genes in wheat reveals gene expression specificity and involvement of STRE and MYB-elements in regulating polyamines under drought. BMC Genom. 2022, 23, 734. [Google Scholar] [CrossRef]
- Fan, S.; Liu, A.; Zhang, Z.; Zou, X.; Jiang, X.; Huang, J.; Fan, L.; Zhang, Z.; Deng, X.; Ge, Q.; et al. Genome-wide identification and expression analysis of the metacaspase gene family in Gossypium species. Genes 2019, 10, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.A.; Homayouni, A.; Gu, L.; Huang, K.Y.; Ho, T.D.; Shen, Q.J. Transcriptomic analysis of rice aleurone cells identified a novel abscisic acid response element. Plant Cell Environ. 2017, 40, 2004–2016. [Google Scholar] [CrossRef] [PubMed]
- Itzhaki, H.; Maxson, J.M.; Woodson, W.R. An ethylene-responsive enhancer element is involved in the senescence-related expression of the carnation glutathione-S-transferase (GST1) gene. Proc. Natl. Acad. Sci. USA 1994, 91, 8925–8929. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Zhang, B.; Chen, L.; Zhang, J.; Zhang, X.; Zhu, C. Isolation and functional characterization of the promoters of miltiradiene synthase genes, TwTPS27a and TwTPS27b, and interaction analysis with the transcription factor TwTGA1 from Tripterygium wilfordii. Plants 2021, 10, 418. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Liu, X.; Teixeira da Silva, J.A.; Wang, H.; Peng, T.; Zhang, M.; Si, C.; Yu, Z.; Tan, J.; Zhang, J.; et al. Characterization of LEA genes in Dendrobium officinale and one gene in induction of callus. J. Plant Physiol. 2021, 258–259, 153356. [Google Scholar] [CrossRef] [PubMed]
- Tanin, M.J.; Saini, D.K.; Sandhu, K.S.; Pal, N.; Gudi, S.; Chaudhary, J.; Sharma, A. Consensus genomic regions associated with multiple abiotic stress tolerance in wheat and implications for wheat breeding. Sci. Rep. 2022, 12, 13680. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Class | Chromosome Position | Intron Number | Average Intron Length (bp) | Protein Length (aa) | pI |
|---|---|---|---|---|---|---|---|
| KoNAC1 | GWHPACBH000260.1 | VII | Chr01: 1952420-1954549 | 2 | 210 | 375 | 8.33 |
| KoNAC2 | GWHPACBH000261.1 | X | Chr01: 1966355-1968293 | 2 | 213 | 381 | 6.38 |
| KoNAC3 | GWHPACBH001011.1 | IV | Chr01: 9324213-9325936 | 2 | 179 | 268 | 9.76 |
| KoNAC4 | GWHPACBH001737.1 | IX | Chr02: 545874-547928 | 2 | 540 | 314 | 4.72 |
| KoNAC5 | GWHPACBH002133.1 | VI | Chr02: 5018997-5022392 | 2 | 627 | 414 | 4.56 |
| KoNAC6 | GWHPACBH002150.1 | VII | Chr02: 5630019-5632845 | 2 | 557 | 291 | 6.26 |
| KoNAC7 | GWHPACBH002927.1 | X | Chr02: 12147292-12149791 | 2 | 695 | 308 | 9.72 |
| KoNAC8 | GWHPACBH002942.1 | VII | Chr02: 12271496-12273886 | 2 | 265 | 397 | 6.91 |
| KoNAC9 | GWHPACBH003351.1 | VII | Chr03: 1575855-1576587 | 1 | 152 | 170 | 10.01 |
| KoNAC10 | GWHPACBH003542.1 | X | Chr03: 2845984-2847971 | 3 | 82 | 358 | 8.29 |
| KoNAC11 | GWHPACBH003714.1 | IV | Chr03: 4257070-4258248 | 2 | 188 | 215 | 10.07 |
| KoNAC12 | GWHPACBH004035.1 | VI | Chr03: 8571932-8575841 | 7 | 267 | 573 | 4.35 |
| KoNAC13 | GWHPACBH004037.1 | VI | Chr03: 8580877-8585584 | 3 | 922 | 432 | 5.87 |
| KoNAC14 | GWHPACBH004257.1 | III | Chr03: 10268576-10273325 | 6 | 484 | 340 | 8.05 |
| KoNAC15 | GWHPACBH005193.1 | VII | Chr04: 4833704-4835719 | 2 | 237 | 301 | 6.63 |
| KoNAC16 | GWHPACBH005487.1 | II | Chr04: 8899908-8901721 | 2 | 518 | 259 | 7.96 |
| KoNAC17 | GWHPACBH005488.1 | II | Chr04: 8902766-8904022 | 1 | 590 | 220 | 8.48 |
| KoNAC18 | GWHPACBH005795.1 | VII | Chr04: 10981103-10983284 | 2 | 229 | 317 | 9.64 |
| KoNAC19 | GWHPACBH005903.1 | III | Chr04: 11744358-11747114 | 4 | 148 | 304 | 5.55 |
| KoNAC20 | GWHPACBH005980.1 | VII | Chr05: 171989-173574 | 2 | 182 | 285 | 8.57 |
| KoNAC21 | GWHPACBH006496.1 | II | Chr05: 3585985-3588070 | 2 | 179 | 576 | 5.03 |
| KoNAC22 | GWHPACBH006945.1 | VII | Chr05: 8286321-8289550 | 2 | 626 | 425 | 7.89 |
| KoNAC23 | GWHPACBH007161.1 | VII | Chr05: 10223818-10225996 | 2 | 171 | 372 | 8.40 |
| KoNAC24 | GWHPACBH007671.1 | IX | Chr06: 2167096-2168677 | 2 | 300 | 327 | 6.13 |
| KoNAC25 | GWHPACBH007697.1 | VI | Chr06: 2347486-2351319 | 4 | 385 | 591 | 4.47 |
| KoNAC26 | GWHPACBH007806.1 | VII | Chr06: 3191353-3193417 | 2 | 295 | 303 | 6.78 |
| KoNAC27 | GWHPACBH007934.1 | II | Chr06: 4507683-4512207 | 5 | 551 | 365 | 5.29 |
| KoNAC28 | GWHPACBH008265.1 | X | Chr06: 8671993-8674297 | 2 | 374 | 441 | 6.35 |
| KoNAC29 | GWHPACBH009271.1 | VI | Chr07: 7943036-7946752 | 5 | 272 | 592 | 4.37 |
| KoNAC30 | GWHPACBH009626.1 | X | Chr08: 595220-599512 | 2 | 1624 | 300 | 7.03 |
| KoNAC31 | GWHPACBH009686.1 | VII | Chr08: 1067480-1068977 | 2 | 114 | 343 | 9.60 |
| KoNAC32 | GWHPACBH009687.1 | VII | Chr08: 1074862-1076943 | 2 | 171 | 347 | 8.70 |
| KoNAC33 | GWHPACBH009711.1 | X | Chr08: 1347371-1350316 | 2 | 636 | 357 | 7.67 |
| KoNAC34 | GWHPACBH009970.1 | X | Chr08: 3208565-3211847 | 2 | 1096 | 286 | 7.10 |
| KoNAC35 | GWHPACBH010230.1 | V | Chr08: 6804416-6807401 | 4 | 274 | 345 | 5.82 |
| KoNAC36 | GWHPACBH010231.1 | V | Chr08: 6808948-6810099 | 3 | 123 | 261 | 4.47 |
| KoNAC37 | GWHPACBH010248.1 | IX | Chr08: 6936960-6938647 | 2 | 191 | 366 | 7.37 |
| KoNAC38 | GWHPACBH010352.1 | VII | Chr08: 7704131-7705968 | 2 | 435 | 305 | 6.78 |
| KoNAC39 | GWHPACBH010982.1 | VII | Chr09: 1762983-1763731 | 1 | 160 | 176 | 9.98 |
| KoNAC40 | GWHPACBH011181.1 | II | Chr09: 3089069-3090025 | 1 | 112 | 236 | 6.50 |
| KoNAC41 | GWHPACBH011224.1 | X | Chr09: 3378802-3380434 | 2 | 115 | 356 | 7.92 |
| KoNAC42 | GWHPACBH011287.1 | VIII | Chr09: 3895422-3899009 | 2 | 1261 | 243 | 4.62 |
| KoNAC43 | GWHPACBH011356.1 | X | Chr09: 4526977-4529037 | 2 | 553 | 318 | 7.21 |
| KoNAC44 | GWHPACBH012132.1 | III | Chr10: 2632918-2637871 | 5 | 584 | 373 | 7.08 |
| KoNAC45 | GWHPACBH012473.1 | VI | Chr10: 5929512-5932298 | 3 | 255 | 574 | 4.20 |
| KoNAC46 | GWHPACBH012540.1 | X | Chr10: 7425092-7428318 | 3 | 579 | 333 | 9.19 |
| KoNAC47 | GWHPACBH012930.1 | VIII | Chr11: 4340404-4343638 | 4 | 342 | 326 | 5.37 |
| KoNAC48 | GWHPACBH013040.1 | IX | Chr11: 5432389-5434082 | 2 | 285 | 354 | 6.19 |
| KoNAC49 | GWHPACBH013264.1 | IX | Chr11: 7036403-7038056 | 3 | 165 | 331 | 6.75 |
| KoNAC50 | GWHPACBH013622.1 | VII | Chr12: 1361271-1363117 | 2 | 215 | 294 | 9.80 |
| KoNAC51 | GWHPACBH013650.1 | V | Chr12: 1584267-1587516 | 4 | 219 | 534 | 4.43 |
| KoNAC52 | GWHPACBH013651.1 | V | Chr12: 1588784-1591086 | 4 | 165 | 336 | 6.58 |
| KoNAC53 | GWHPACBH013779.1 | X | Chr12: 3635952-3638945 | 2 | 591 | 354 | 8.79 |
| KoNAC54 | GWHPACBH013961.1 | X | Chr12: 5551002-5553482 | 3 | 571 | 256 | 9.26 |
| KoNAC55 | GWHPACBH014293.1 | III | Chr12: 7828847-7830888 | 2 | 494 | 296 | 8.61 |
| KoNAC56 | GWHPACBH014606.1 | VII | Chr13: 304056-306202 | 2 | 230 | 255 | 9.29 |
| KoNAC57 | GWHPACBH014849.1 | VII | Chr13: 2243283-2244839 | 2 | 109 | 265 | 7.90 |
| KoNAC58 | GWHPACBH015137.1 | III | Chr13: 8039942-8044242 | 5 | 466 | 461 | 4.62 |
| KoNAC59 | GWHPACBH015463.1 | VII | Chr14: 3448051-3450062 | 2 | 222 | 255 | 9.64 |
| KoNAC60 | GWHPACBH015782.1 | V | Chr15: 43374-47367 | 5 | 324 | 638 | 4.57 |
| KoNAC61 | GWHPACBH015958.1 | IX | Chr15: 1512975-1513703 | 1 | 125 | 162 | 9.55 |
| KoNAC62 | GWHPACBH016068.1 | VIII | Chr15: 2698343-2700471 | 4 | 261 | 306 | 9.33 |
| KoNAC63 | GWHPACBH016810.1 | III | Chr16: 4714504-4716464 | 2 | 516 | 304 | 9.25 |
| KoNAC64 | GWHPACBH017115.1 | VII | Chr17: 1665396-1666616 | 2 | 119 | 245 | 8.31 |
| KoNAC65 | GWHPACBH017251.1 | IV | Chr17: 2901510-2902567 | 1 | 103 | 314 | 7.77 |
| KoNAC66 | GWHPACBH017590.1 | VII | Chr17: 5655463-5657073 | 2 | 147 | 358 | 8.88 |
| KoNAC67 | GWHPACBH017594.1 | VII | Chr17: 5688319-5689928 | 2 | 146 | 358 | 8.88 |
| KoNAC68 | GWHPACBH017666.1 | III | Chr17: 6866174-6868665 | 2 | 727 | 325 | 7.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.-M.; Liu, X.; Huang, X.-J.; Yang, J.-J.; Qin, P.-T.; Zhou, H.; Jiang, M.-G.; Liao, H.-Z. Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia obovata, a Typical Mangrove Plant. Curr. Issues Mol. Biol. 2022, 44, 5622-5637. https://doi.org/10.3390/cimb44110381
Sun M-M, Liu X, Huang X-J, Yang J-J, Qin P-T, Zhou H, Jiang M-G, Liao H-Z. Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia obovata, a Typical Mangrove Plant. Current Issues in Molecular Biology. 2022; 44(11):5622-5637. https://doi.org/10.3390/cimb44110381
Chicago/Turabian StyleSun, Man-Man, Xiu Liu, Xiao-Juan Huang, Jing-Jun Yang, Pei-Ting Qin, Hao Zhou, Ming-Guo Jiang, and Hong-Ze Liao. 2022. "Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia obovata, a Typical Mangrove Plant" Current Issues in Molecular Biology 44, no. 11: 5622-5637. https://doi.org/10.3390/cimb44110381
APA StyleSun, M. -M., Liu, X., Huang, X. -J., Yang, J. -J., Qin, P. -T., Zhou, H., Jiang, M. -G., & Liao, H. -Z. (2022). Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia obovata, a Typical Mangrove Plant. Current Issues in Molecular Biology, 44(11), 5622-5637. https://doi.org/10.3390/cimb44110381