
Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature

 , , ,
, , ,  , , and
, , and 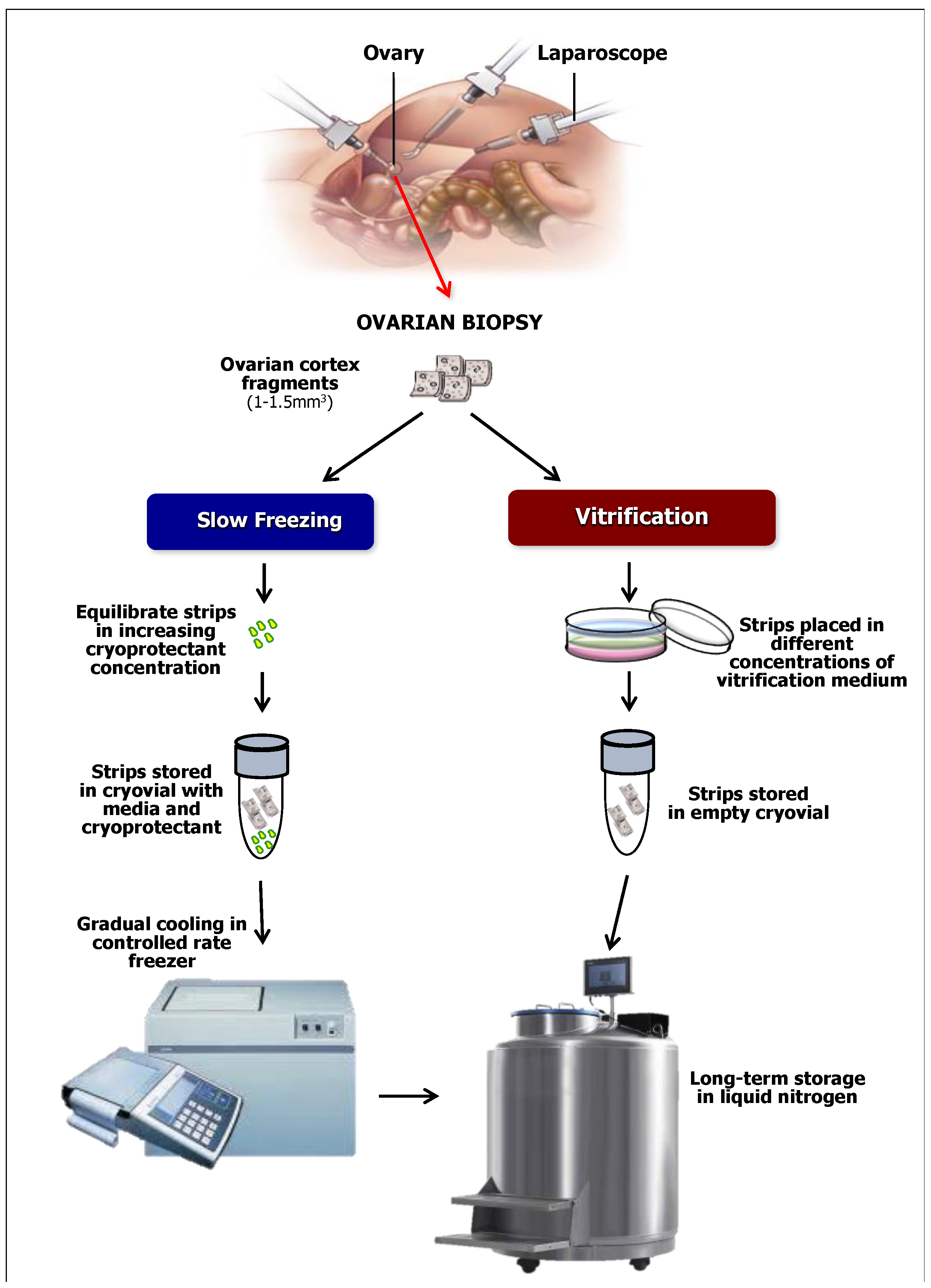{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Ovarian Stem Cells to Regenerate Folliculogenesis
3. The Cryostored Ovarian Cortical Strips as a Source of OSCs
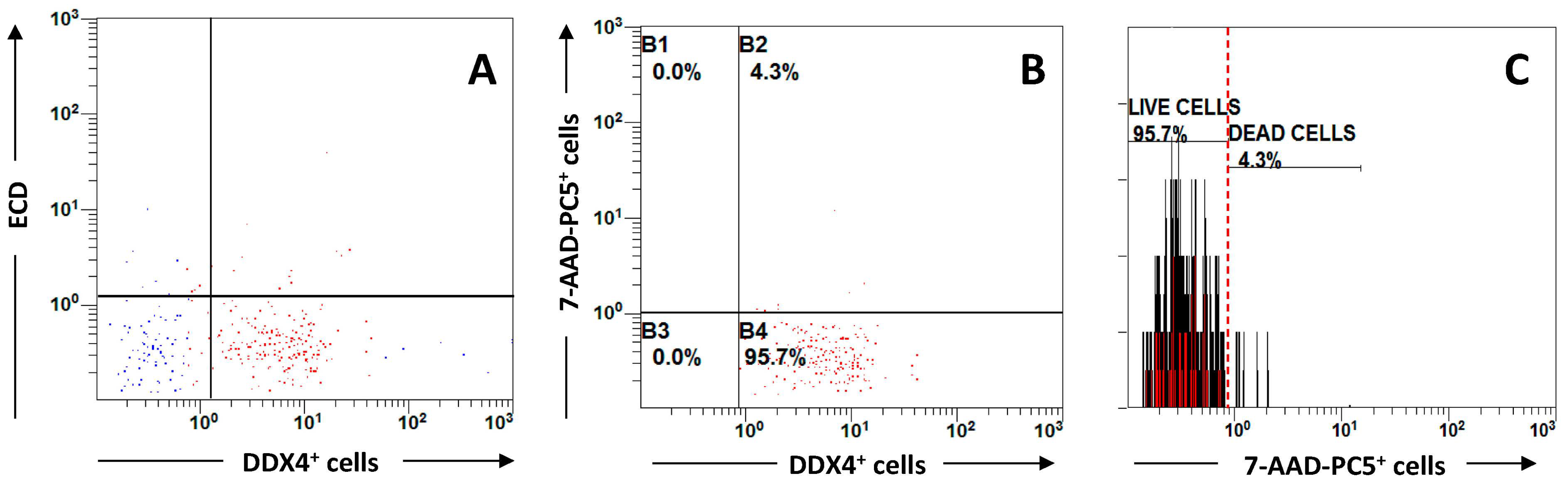
4. OSC from Cryopreserved Ovarian Cortical Fragments: A Preliminary Observation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akahori, T.; Woods, D.C.; Tilly, J.L. Female fertility preservation through stem cell-based ovarian tissue reconstitution in vitro and ovarian regeneration in vivo. Clin. Med. Insights Reprod. Health 2019, 13, 1179558119848007. [Google Scholar] [CrossRef] [PubMed]
- Cancer Statistics Center. American Cancer Society. 2020. Available online: http://cancerstatisticscenter.cancer.org (accessed on 8 January 2020).
- Italian Cancer Numbers. 2020. Available online: http://knowandbe.live/en/news/63-italian-cancer-numbers-2020.html (accessed on 10 May 2021).
- Aakash, D.; Turab, J.M.; Duma, N.; Garassino, M.C.; Hicks, L.K.; Kuderer, N.M.; Lyman, G.H.; Mishra, S.; Pinato, D.J.; Rini, B.I.; et al. COVID-19 and cancer a review of the registry-based pandemic response. JAMA Oncol. 2021, 7, 12. [Google Scholar]
- Waxman, J. Chemotherapy and the adult gonad: A review. J. R. Soc. Med. 1983, 76, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichman, B.S.; Green, K.B. Breast cancer in young women: Effect of chemotherapy on ovarian function, fertility and birth defects. J. Natl. Cancer Inst. Monogr. 1994, 16, 125–129. [Google Scholar]
- Chemaitilly, W.; Mertens, A.C.; Mitby, P.; Whitton, J.; Stovall, M.; Yasui, Y.; Robison, L.L.; Sklar, C.A. Acute ovarian failure in the childhood cancer survivor study. J. Clin. Endocrinol. Metab. 2006, 91, 1723–1728. [Google Scholar] [CrossRef]
- Sklar, C.A.; Mertens, A.C.; Mitby, P.; Whitton, J.; Stovall, M.; Kasper, C.; Mulder, J.; Green, D.; Nicholson, H.S.; Yasui, Y.; et al. Premature menopause in survivors of childhood cancer: A report from the childhood cancer survivor study. J. Natl. Cancer Inst. 2006, 98, 890–896. [Google Scholar] [CrossRef]
- Levine, J.M.; Kelvin, J.F.; Quinn, G.P.; Gracia, C.R. Infertility in reproductive-age female cancer survivors. Cancer 2015, 121, 1532–1539. [Google Scholar] [CrossRef]
- Blumenfeld, Z.; Von Wolff, M. GnRH-analogues and oral contraceptives for fertility preservation in women during chemotherapy. Hum. Reprod. Update 2008, 14, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Blumenfeld, Z. Fertility preservation by endocrine suppression of ovarian function using gonadotropin-releasing hormone agonists: The end of the controversy? J. Clin. Oncol. 2018, 36, 1895–1897. [Google Scholar] [CrossRef]
- Lambertini, M.; Anserini, P.; Fonana, V.; Poggio, F.; Iacono, G.; Abate, A.; Levaggi, A.; Miglietta, L.; Bighin, C.; Giraudi, S.; et al. The PREgnancy and FERtility (PREFER) study: An Italian multicenter prospective cohort study on fertility preservation and pregnancy issues in young breast cancer patients. BMC Cancer 2017, 17, 346. [Google Scholar] [CrossRef]
- Lambertini, M.; Moore, C.F.; Leonard, R.C.F.; Sibylle, L.; Munster, P.; Bruzzone, M.; Boni, L.; Unger, J.M.; Anderson, R.A.; Mehta, K.; et al. Gonadotropin-releasing hormone agonists during chemotherapy for preservation of ovarian function and fertility in premenopausal patients with early breast cancer: A systematic review and meta- analysis of individual patient-level data. J. Clin. Oncol. 2018, 36, 1981–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeestere, I.; Brice, P.; Peccatori, F.A.; Kentos, A.; Gaillard, I.; Zachee, P.; Casasnovas, R.O.; Van Den Neste, E.; Dechene, J.; De Maertelaer, V.; et al. Gonadotropin-releasing hormone agonist for the prevention of chemotherapy-induced ovarian failure in patients with lymphoma: 1-year follow-up of a prospective randomized trial. J. Clin. Oncol. 2013, 31, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Turan, V.; Bedoschi, G.; Rodriguez-Wallberg, K.; Sonmezer, M.; Silva Pacheco, F.; Oktem, O.; Taylor, H.; Oktay, K. Utility of gonadotropin-releasing hormone agonists for fertility preservation: Lack of biologic basis and the need to prioritize proven methods. J. Clin. Oncol. 2019, 37, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Dolmans, M.M.; Taylor, H.S.; Rodriguez-Wallberg, K.A.; Blumenfeld, Z.; Lambertini, M.; Von Wolff, M.; Donnez, J. Utility of gonadotropin-releasing hormone agonists for fertility preservation in women receiving chemotherapy: Pros and cons. Fertil. Steril. 2020, 114, 725–738. [Google Scholar] [CrossRef]
- Son, W.Y.; Chung, J.T.; Demirtas, E.; Chung, J.T.; Demirtas, E.; Holzer, H.; Sylvestre, C.; Buckett, W.; Chian, R.C.; Tan, S.L. Comparison of in-vitro maturation cycles with and without in-vivo matured oocytes retrieved. Reprod. Biomed. Online 2008, 17, 59–67. [Google Scholar] [CrossRef]
- Oktay, K.; Buyuk, E.; Rodriguez-Wallberg, K.A.; Sahin, G. In vitro maturation improves oocyte or embryo cryopreservation outcome in breast cancer patients undergoing ovarian stimulation for fertility preservation. Reprod. Biomed. Online 2010, 20, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Prasath, E.B.; Chan, M.L.; Wong, W.H.; Lim, C.J.W.; Tharmalingam, M.D.; Hendricks, H.; Loh, S.F.; Chia, Y.N. First pregnancy and live birth resulting from cryopreserved embryos obtained from in vitro matured oocytes after oophorectomy in an ovarian cancer patient. Hum. Reprod. 2014, 29, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Donnez, J.; Dolmans, M.M.; Demylle, D.; Jadoul, P.; Pirard, C.; Squifflet, J.; Martinez-Madrid, B.; Van Langendonckt, A. Livebirth after orthotopic transplantation of cryopreserved ovarian tissue. Lancet 2004, 364, 1405–1410. [Google Scholar] [CrossRef]
- Oktay, K.; Buyuk, E.; Veeck, L.; Zaninovic, N.; Xu, K.; Takeuchi, T.; Opsahl, M.; Rosenwaks, Z. Embryo development after heterotopic transplantation of cryopreserved ovarian tissue. Lancet 2004, 363, 837–840. [Google Scholar] [CrossRef]
- Von Wolff, M.; Donnez, J.; Hovatta, O.; Keros, V.; Maltaris, T.; Montag, M.; Salle, B.; Sonmezer, M.; Andersen, C. Cryopreservation and autotransplantation of human ovarian tissue prior to cytotoxic therapy—A technique in its infancy but already successful in fertility preservation. Eur. J. Cancer 2009, 45, 1547–1553. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, J.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, K.; Ishizuka, B.; Hsueh, A.J. Drug-free in-vitro activation of follicles for infertility treatment in poor ovarian response patients with decreased ovarian reserve. Reprod. Biomed. Online 2020, 40, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Lunding, S.A.; Andersen, A.N.; Hardardottir, L.; Olesen, H.; Kristensen, S.G.; Andersen, C.Y.; Pors, S.E. Hippo signaling, actin polymerization, and follicle activation in fragmented human ovarian cortex. Mol. Reprod. Dev. 2020, 87, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef]
- Pacchiarotti, J.; Maki, C.; Ramos, T.; Marh, J.; Homeworton, K.; Wong, J.; Pham, J.; Anorve, S.; Chow, Y.C.; Izadyar, F. Differentiation potential of germ line stem cells derived from the postnatal mouse ovary. Differentiation 2010, 79, 159–170. [Google Scholar] [CrossRef]
- Bhartiya, D.; Patel, H. Ovarian stem cells-resolving controversies. J. Assit. Reprod. Genet. 2018, 3, 393–398. [Google Scholar] [CrossRef]
- Bothun, A.M.; Gao, Y.; Takai, Y.; Ishihara, O.; Seki, H.; Karger, B.; Tilly, J.L.; Woods, D.C. Quantitative Proteomic Profiling of the Human Ovary from Early to Mid-Gestation Reveals Protein Expression Dynamics of Oogenesis and Folliculogenesis. Stem Cells Dev. 2018, 27, 723–735. [Google Scholar] [CrossRef]
- Zuckerman, S. The number of oocytes in the mature ovary. Rec. Prog. Horm. Res. 1951, 6, 63–108. [Google Scholar]
- Silvestris, E.; Cafforio, P.; D’Oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Guo, K.; Li, C.H.; Wang, X.Y.; He, D.J.; Zheng, P. Germ stem cells are active in postnatal mouse ovary under physiological conditions. Mol. Hum. Reprod. 2016, 22, 316–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bristol-Gould, S.K.; Kreeger, P.K.; Selkirk, C.G.; Kilen, S.M.; Mayo, K.E.; Shea, L.D.; Woodruff, T.K. Fate of the initial follicle pool: Empirical and mathematical evidence supporting its sufficiency for adult fertility. Dev. Biol. 2006, 1, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wu, C.; Lyu, Q.; Yang, D.; Albertini, D.F.; Keefe, D.L.; Liu, L. Germline stem cells and neo-oogenesis in the adult human ovary. Dev. Biol. 2007, 306, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byskov, A.G.; Faddy, M.J.; Lemmen, J.G.; Andersen, C.Y. Eggs forever? Differentiation 2005, 73, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.B.; Brogan, L.; Myers, M.; Hutt, K.J.; Mladenovska, T.; Ricardo, S.; Hamza, K.; Scott, C.L.; Strasser, A.; Findlay, J.K. The primordial follicle reserve is not renewed after chemical or -irradiation mediated depletion. Reproduction 2012, 143, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zheng, W.; Shen, Y.; Adhikari, D.; Ueno, H.; Liu, K. Experimental evidence showing that no mitotically active female germline progenitors exist in postnatal mouse ovaries. Proc. Natl. Acad. Sci. USA 2012, 109, 12580–12585. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Spradling, A.C. Female mice lack adult germ-line stem cells but sustain oogenesis using stable primordial follicles. Proc. Natl. Acad. Sci. USA 2013, 110, 8585–8590. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhang, D.; Wang, L.; Liu, M.; Mao, J.; Yin, Y.; Ye, X.; Liu, N.; Han, J.; Gao, Y. No evidence for neo-oogenesis may link to ovarian senescence in adult monkey. Stem Cells 2013, 31, 2538–2550. [Google Scholar] [CrossRef]
- Bukovsky, A.; Svetlikova, M.; Caudle, M.R. Oogenesis in cultures derived from adult human ovaries. Reprod. Biol. Endocrinol. 2005, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.; Bagley, J.; Skaznik-Wikiel, M.; Lee, H.J.; Adams, G.B.; Niikura, Y.; Tschudy, K.S.; Tilly, J.C.; Cortes, M.L.; Forkert, R. Oocyte generation in adult mammalian ovaries by p.putative germ cells in bone marrow and peripheral blood. Cell 2005, 122, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Virant-Klun, I.; Zech, N.; Rozman, P.; Vogler, A.; Cvjeticanin, B.; Klemenc, P.; Malicev, E.; Meden-Vrtovec, H. Putative stem cells with an embryonic character isolated from the ovarian surface epithelium of women with no naturally present follicles and oocytes. Differentiation 2008, 76, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Niikura, Y.; Niikura, T.; Tilly, J.L. Aged mouse ovaries possess rare premeiotic germ cells that can generate oocytes following transplantation into a young host environment. Aging 2009, 1, 971–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Oktem, O.; Oktay, K. Stem cells: A perspective on oocytes. Ann. N. Y. Acad. Sci. 2008, 1127, 20–26. [Google Scholar] [CrossRef]
- Parte, S.; Bhartiya, D.; Telang, J.; Daithankar, V.; Salvi, V.; Zaveri, K.; Hinduja, I. Detection, characterization, and spontaneous differentiation in vitro of very small embryonic-like putative stem cells in adult Mammalian ovary. Stem Cells Dev. 2011, 20, 1451–1464. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Yang, Y.; Wang, S.; Shi, L.; Xie, W.; Sun, K.; Zou, K.; Wang, L.; Xiong, J. Production of transgenic mice by random recombination of targeted genes in female germline stem cells. J. Mol. Cell Biol. 2011, 3, 132–141. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Kang, J.X.; Xie, W.; Li, X.; Wu, C.; Xu, B.; Wu, J. Production of fat-1 transgenic rats using a post-natal female germline stem cell line. Mol. Hum. Reprod. 2014, 20, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Satirapod, C.; Ohguchi, Y.; Park, E.S.; Woods, D.C.; Tilly, J.L. Genetic studies in mice directly link oocytes produced during adulthood to ovarian function and natural fertility. Sci. Rep. 2017, 7, 10011. [Google Scholar] [CrossRef]
- Martin, J.J.; Woods, D.C.; Tilly, J.L. Implications and current limitations of oogenesis from female germline or oogonial stem cells in adult mammalian ovaries. Cells 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Park, E.S.; Tilly, J.L. Use of DEAD-box polypeptide-4 (Ddx4) gene promoter-driven fluorescent reporter mice to identify mitoti-cally active germ cells in post-natal mouse ovaries. Mol. Hum. Reprod. 2015, 21, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Parte, S.; Bhartiya, D.; Patel, H.; Daithankar, V.; Chauhan, A.; Zaveri, K.; Hinduja, I. Dynamics associated with spontaneous differentiation of ovarian stem cells in vitro. J. Ovarian Res. 2014, 25, 7–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, Y.A.R.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telfer, E.; ALbertini, D. The quest for human ovarian stem cells. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Panula, S.; Petropoulos, S.; Edsgärd, D.; Busayavalasa, K.; Liu, L.; Li, X.; Risal, S.; Shen, Y.; Shao, J.; et al. Adult human and mouse ovaries lack DDX4-expressing functional oogonial stem cells. Nat. Med. 2015, 21, 1116–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, S.F.; Vahidi, N.A.; Park, S.; Weitzel, R.P.; Tisdale, J.; Rueda, B.R.; Wolff, E.F. Characterization of extracellular DDX4- or Ddx4- positive ovarian cells. Nat. Med. 2015, 21, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 1, 1147. [Google Scholar] [CrossRef] [Green Version]
- Castrillon, D.H.; Quade, B.J.; Wang, T.Y.; Quigley, C.; Crum, C.P. The human VASA gene is specifically expressed in the germ cell lineage. Proc. Natl. Acad. Sci. USA 2000, 97, 9585–9590. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, J. Production of offspring from a germline stem cell line derived from prepubertal ovaries of germline reporter mice. Mol. Hum. Reprod. 2016, 22, 457–464. [Google Scholar] [CrossRef]
- Zou, K.; Hou, L.; Sun, K.; Xie, W.; Wu, J. Improved efficiency of female germline stem cell purification using fragilis-based mag- netic bead sorting. Stem Cells Dev. 2011, 20, 2197–2204. [Google Scholar] [CrossRef]
- Sequeira, R.C.; Sittadjody, S.; Criswell, T.; Atala, A.; Jackson, J.D.; Yoo, J.J. Enhanced method to select human oogonial stem cells for fertility research. Cell Tissue Res. 2021, 386, 145–156. [Google Scholar] [CrossRef]
- Truman, A.M.; Tilly, J.L.; Woods, D.C. Ovarian regeneration: The potential for stem cell contribution in the postnatal ovary to sustained endocrine function. Mol. Cell Endocrinol. 2017, 445, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anchan, R.; Gerami-Naini, B.; Lindsey, J.S.; Ho, J.W.K.; Kiezun, A.; Lipskind, S.; Ng, N.; LiCausi, J.A.; Kim, C.S.; Brezina, P.; et al. Efficient differentiation of steroidogenic and germ-like cells from epigenetically-related iPSCs derived from ovarian granulosa cells. PLoS ONE 2015, 10, e0119275. [Google Scholar] [CrossRef] [PubMed]
- Da Broi, M.G.; Giorgi, V.S.I.; Wang, F.; Keefe, D.L.; Albertini, D.; Navarro, P.A. Influence of follicular fluid and cumulus cells on oocyte quality: Clinical implications. J. Assist. Reprod. Genet. 2018, 5, 735–751. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.M.; Pellicer, A.; Diaz-Garcia, C.; Serrano, M.S.; Schmidt, K.T.; Ernst, E.; Luyckx, V.; Andersen, C.Y. Restoration of ovarian activity and pregnancy after transplantation of cryopreserved ovarian tissue: A review of 60 cases of reimplantation. Fertil. Steril. 2013, 99, 1503–1513. [Google Scholar] [CrossRef]
- Meirow, D.; Fasouliotis, S.J.; Nugent, D.; Schenker, J.G.; Gosden, R.G.; Rutherford, A.G. A laparoscopic technique for obtaining ovarian cortical biopsy specimens for fertility conservation in patients with cancer. Fertil. Steril. 1999, 71, 948–951. [Google Scholar] [CrossRef]
- Silber, S. Ovary cryopreservation and transplantation for fertility preservation. Mol. Hum. Reprod. 2011, 18, 59–67. [Google Scholar] [CrossRef]
- Ferreira, M. The effects of sample size on the outcome of ovarian tissue cryopreservation. Reprod. Domest. Anim. 2010, 45, 99–102. [Google Scholar] [CrossRef]
- Shi, Q. Vitrification versus slow freezing for human ovarian tissue cryopreservation: A systematic review and meta-anlaysis. Sci. Rep. 2017, 7, 8538. [Google Scholar] [CrossRef]
- Andersen, C.Y.; Rosendahl, M.; Byskov, A.G.; Loft, A.; Ottosen, C.; Dueholm, M.; Schmidt, K.L.T.; Andersen, A.N.; Ernst, E. Two successful pregnancies following autotransplantation of frozen/thawed ovarian tissue. Hum. Reprod. 2008, 10, 2266–2272. [Google Scholar] [CrossRef]
- Stern, C.J.; Gook, D.; Hale, L.G.; Agresta, F.; Oldham, J.; Rozen, G.; Jobling, T. First reported clinical pregnancy following heterotopic grafting of cryopreserved ovarian tissue in a woman after a bilateral oophorectomy. Hum. Reprod. 2013, 11, 2996–2999. [Google Scholar] [CrossRef] [Green Version]
- Leonel, E.C.R. Stepped vitrification technique for human ovarian tissue cryopreservation. Sci. Rep. 2019, 9, 20008. [Google Scholar] [CrossRef] [PubMed]
- Sheikhi, M. Preservation of human ovarian follicles within tissue frozen by vitrification in a xeno-free closed system using only ethylene glycol as a permeating cryoprotectant. Fertil. Steril. 2013, 100, 170–177. [Google Scholar] [CrossRef] [PubMed]
- El Cury-Silva, T.; Nunes, M.E.; Casalechi, M.; Comim, F.V.; Rodrigues, J.K.; Reis, F.M. Cryoprotectant agents for ovarian tissue vitrification: Systematic review. Cryobiology 2021, 103, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J. Optimal condition of vitrification method for cryopreservation of human ovarian cortical tissues. J. Obstet. Gynaecol. Res. 2011, 37, 1092–1101. [Google Scholar] [CrossRef]
- Amorim, C.A.; David, A.; Van Langendonckt, A.; Dolmans, M.M.; Donnez, J. Vitrification of human ovarian tissue: Effect of different solutions and procedures. Fertil. Steril. 2011, 3, 1094–1097. [Google Scholar] [CrossRef]
- Demeestere, I.; Simon, P.; Dedeken, L.; Moffa, F.; Tsépélidis, S.; Brachet, C.; Delbaere, A.; Devreker, F.; Ferster, A. Live birth after autograft of ovarian tissue cryopreserved during childhood. Hum. Reprod. 2015, 30, 2107–2109. [Google Scholar] [CrossRef] [Green Version]
- Dolmans, M.M. Transplantation of cryopreserved ovarian tissue in a series of 285 women: A review of five leading European centers. Fertil. Steril. 2021, 115, 1102–1115. [Google Scholar] [CrossRef]
- Donnez, J. Fertility preservation in women. N. Engl. J. Med. 2017, 337, 1657–1665. [Google Scholar] [CrossRef]
- Masciangelo, R.; Bosisio, C.; Donnez, J.; Amorim, C.A.; Dolmans, M.M. Safety of ovarian tissue transplantation in patients with borderline ovarian tumors. Hum. Reprod. 2018, 2, 212–219. [Google Scholar] [CrossRef]
- Silvestris, E.; De Palma Canosa, S.; Palini, S.; Dellino, M.; Revelli, A.; Paradiso, A.V. Human ovarian cortex biobanking: A fascinating resource for fertility preservation in cancer. Int. J. Mol. Sci. 2020, 21, 3245. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvestris, E.; Minoia, C.; Guarini, A.; Opinto, G.; Negri, A.; Dellino, M.; Tinelli, R.; Cormio, G.; Paradiso, A.V.; De Palma, G. Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature. Curr. Issues Mol. Biol. 2022, 44, 2309-2320. https://doi.org/10.3390/cimb44050157
Silvestris E, Minoia C, Guarini A, Opinto G, Negri A, Dellino M, Tinelli R, Cormio G, Paradiso AV, De Palma G. Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature. Current Issues in Molecular Biology. 2022; 44(5):2309-2320. https://doi.org/10.3390/cimb44050157
Chicago/Turabian StyleSilvestris, Erica, Carla Minoia, Attilio Guarini, Giuseppina Opinto, Antonio Negri, Miriam Dellino, Raffaele Tinelli, Gennaro Cormio, Angelo Virgilio Paradiso, and Giuseppe De Palma. 2022. "Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature" Current Issues in Molecular Biology 44, no. 5: 2309-2320. https://doi.org/10.3390/cimb44050157
APA StyleSilvestris, E., Minoia, C., Guarini, A., Opinto, G., Negri, A., Dellino, M., Tinelli, R., Cormio, G., Paradiso, A. V., & De Palma, G. (2022). Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature. Current Issues in Molecular Biology, 44(5), 2309-2320. https://doi.org/10.3390/cimb44050157