
6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cytotoxicity Assay
2.3. IL-6 and CXCL10 Production in TR146 Cells
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
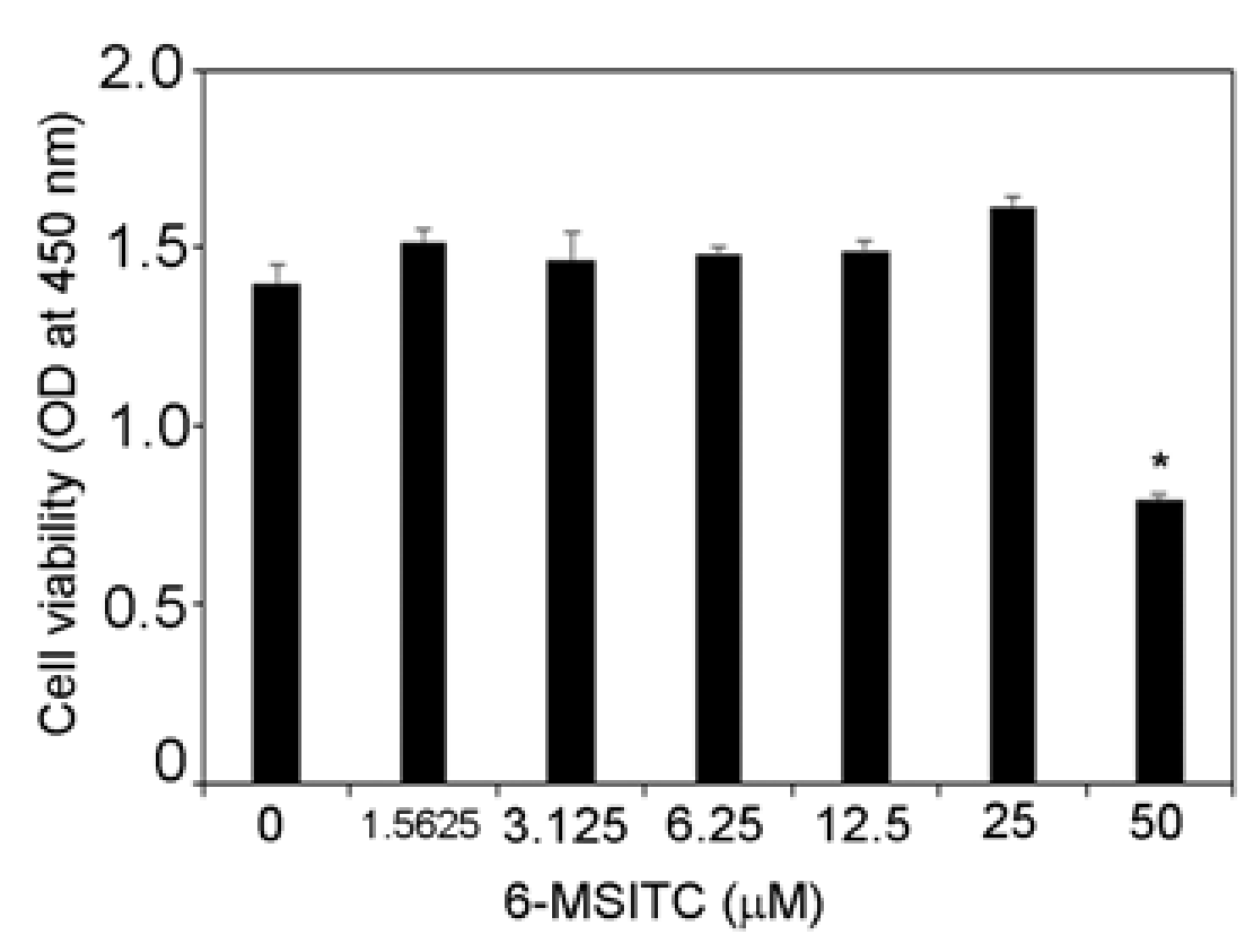
3.1. Effects of 6-MSITC on Cell Viability of TR146 Cells
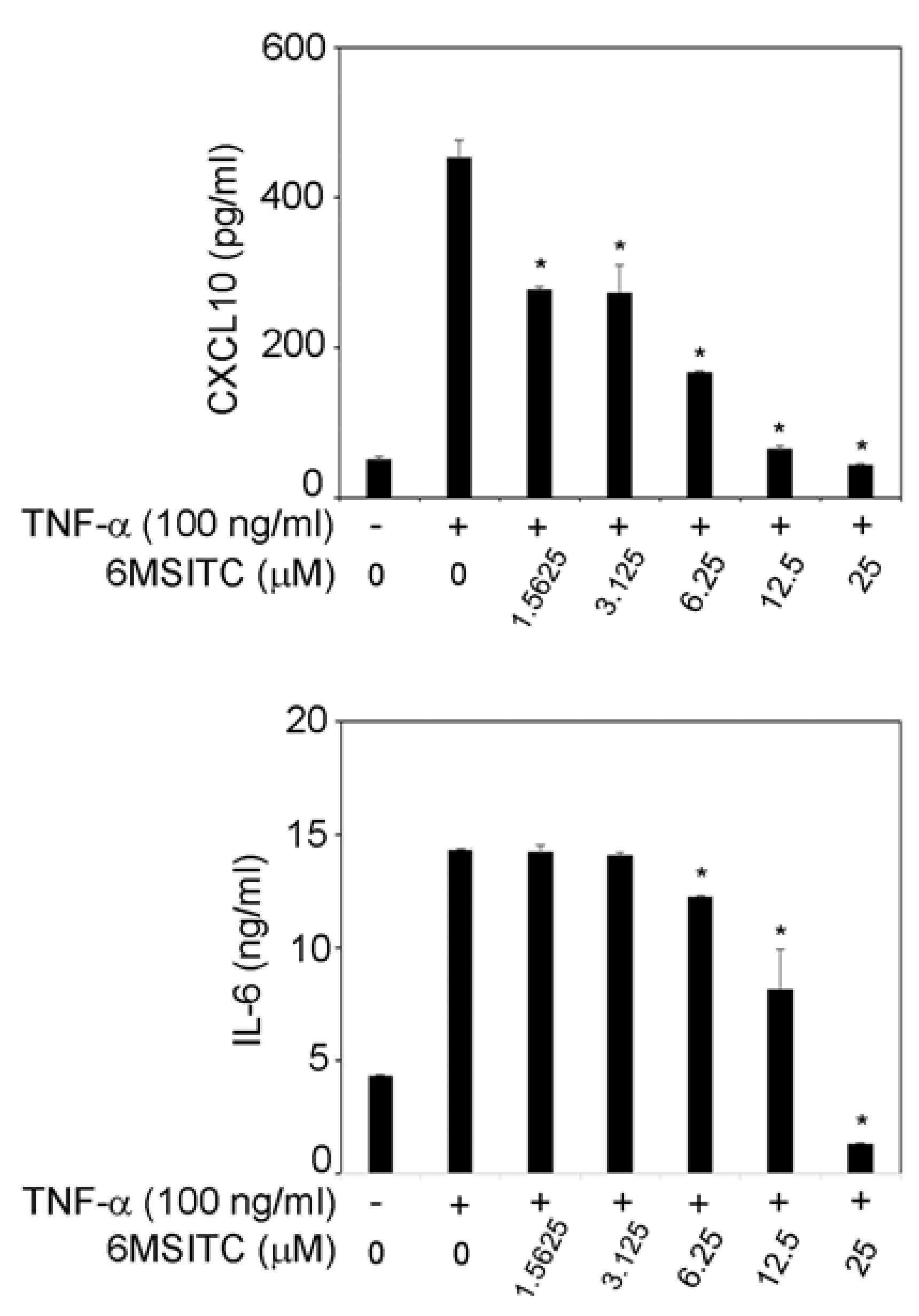
3.2. 6-MSITC Inhibits TNF-α-Induced IL-6 and CXCL10 Production in TR146 Cells
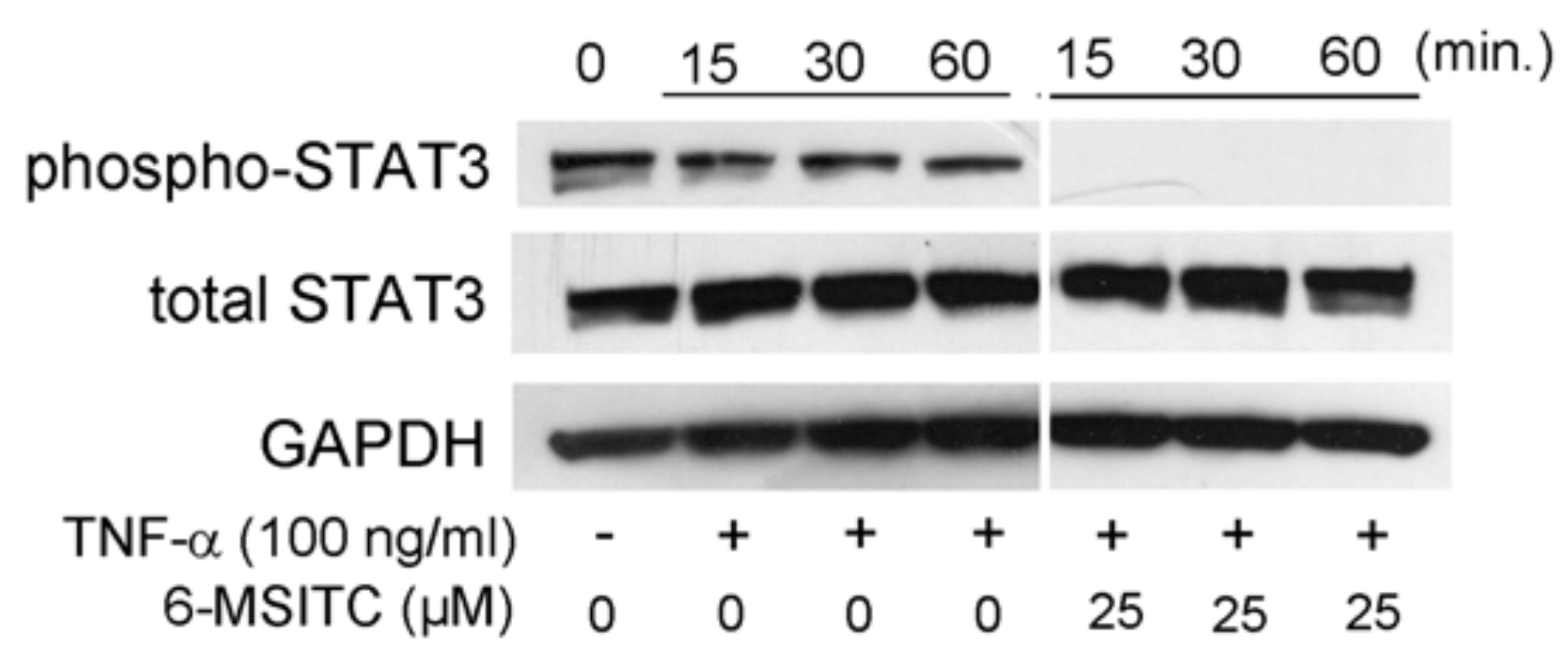
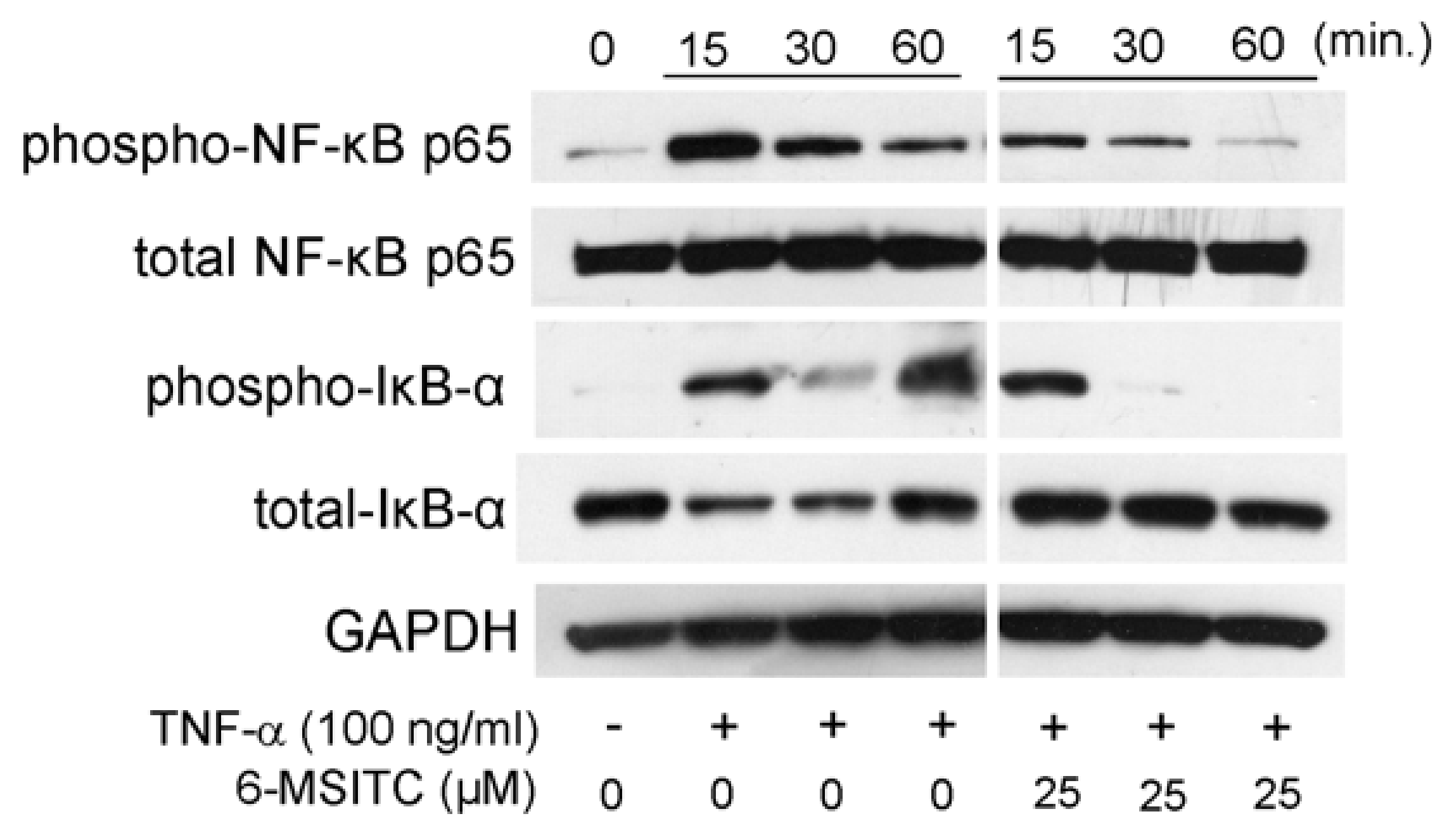
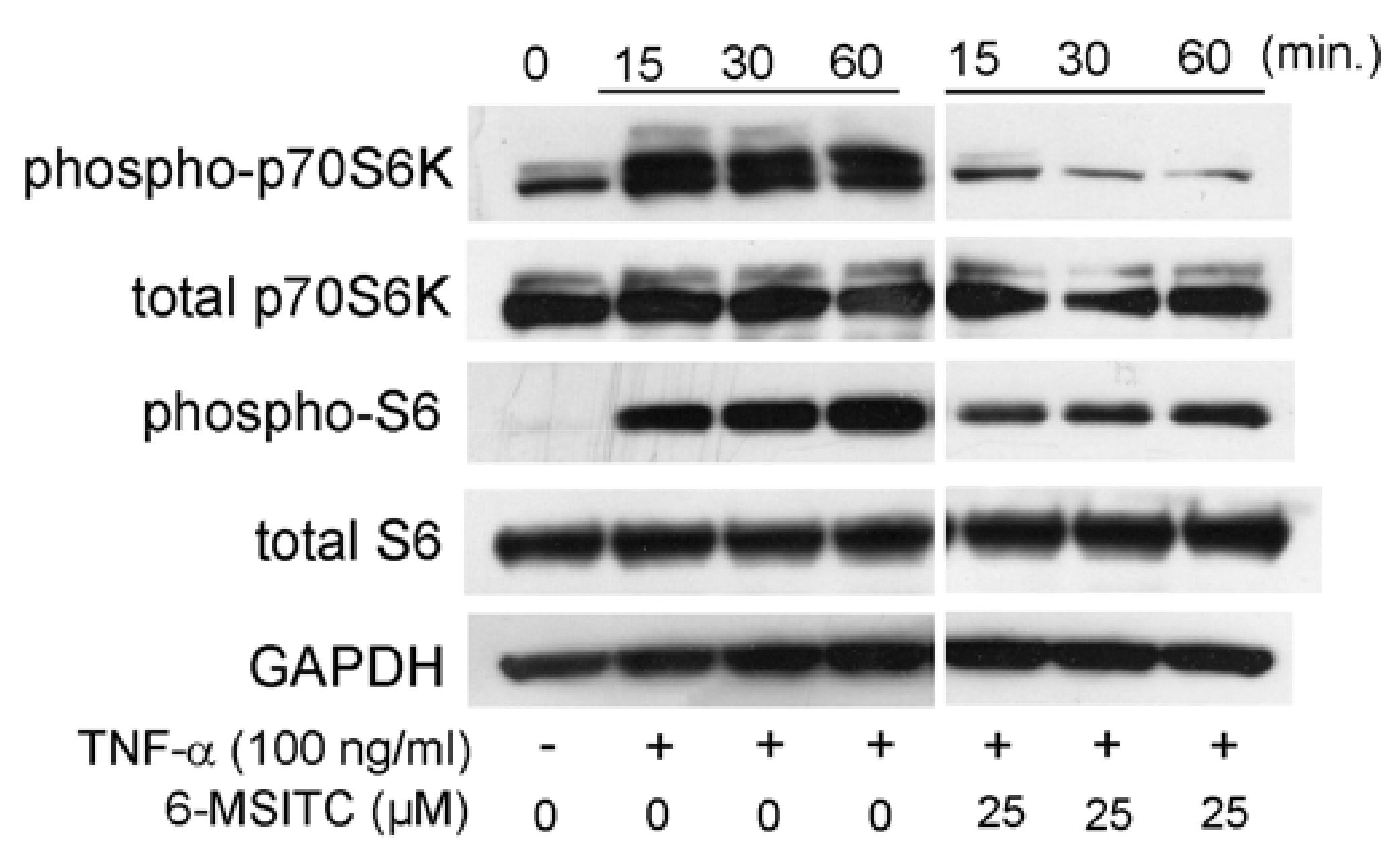
3.3. Effects of STAT3, NF-κB, and p70S6K-S6 Signaling Pathways in TR146 Cells
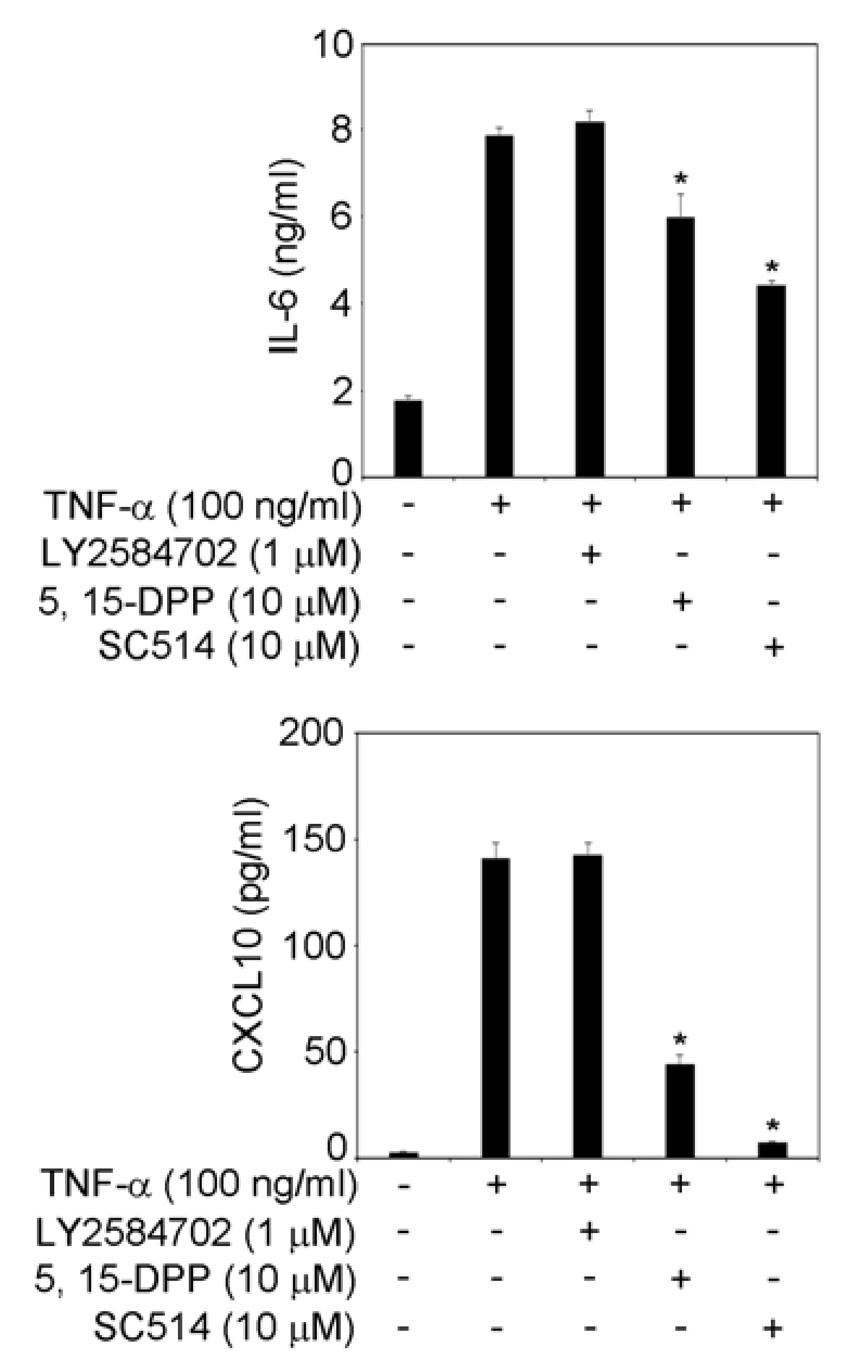
3.4. STAT3 and NF-κB Signaling Pathways Are Involved in IL-6 and CXCL10 Production in TNF-α-Treated TR146 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cai, X.; Ou, Y.; Zhou, Y.; Wang, Y. Resolution of inflammation in periodontitis: A review. Int. J. Clin. Exp. Pathol. 2018, 11, 4283–4295. [Google Scholar] [PubMed]
- Mahuli, S.A.; Zorair, A.M.; Jafer, M.A.; Sultan, A.; Sarode, G.; Baeshen, H.A.; Raj, A.T.; Sarode, S.; Patil, S. Antibiotics for periodontal infections: Biological and clinical perspectives. J. Contemp. Dent. Pract. 2020, 21, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Wu, S.; Sakao, K.; Hou, D.-X. Wasabi 6-(methylsulfinyl)hexyl isothiocyanate induces apoptosis in human colorectal cancer cells through p53-independent mitochondrial dysfunction pathway. BioFactors 2018, 44, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Fuke, Y.; Hishinuma, M.; Namikawa, M.; Oishi, Y.; Matsuzaki, T. Wasabi-derived 6-(methylsulfinyl)hexyl isothiocyanate induces apoptosis in human breast cancer by possible involvement of the NF-κB pathways. Nutr. Cancer 2014, 66, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Uto, T.; Tanigawa, S.; Yamada-Kato, T.; Fujii, M.; Hou, D.X. Microarray-based determination of anti-inflammatory genes targeted by 6-(methylsulfinyl)hexyl isothiocyanate in macrophages. Exp. Ther. Med. 2010, 1, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Akita, N.; Nagai, M.; Hayashi, T.; Suzuki, K. 6-Methylsulfinylhexyl isothiocyanate modulates endothelial cell function and suppresses leukocyte adhesion. J. Nat. Med. 2014, 68, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Kido, R.; Hiroshima, Y.; Kido, J.I.; Ikuta, T.; Sakamoto, E.; Inagaki, Y.; Naruishi, K.; Yumoto, H. Advanced glycation end-products increase lipocalin 2 expression in human oral epithelial cells. J. Periodontal Res. 2020, 55, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Shindo, S.; Ozaki, K.; Matsuo, T. IL-29 enhances CXCL10 production in TNF-α-stimulated human oral epithelial cells. Immunol. Investig. 2017, 46, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Matsuo, T. IL-27 modulates chemokine production in TNF-α-stimulated human oral epithelial cells. Cell. Physiol. Biochem. 2017, 43, 1198–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Atkinson, S.J.; Akbareian, S.E.; Zhou, Z.; Munsterberg, A.; Robinson, S.D.; Bao, Y. Sulforaphane exerts anti-angiogenesis effects against hepatocellular carcinoma through inhibition of STAT3/HIF-1α/VEGF signalling. Sci. Rep. 2017, 7, 12651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawky, N.M.; Segar, L. Sulforaphane inhibits platelet-derived growth factor-induced vascular smooth muscle cell proliferation by targeting mTOR/p70S6kinase signaling independent of Nrf2 activation. Pharmacol. Res. 2017, 119, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Chen, B.; Wang, X.; Wu, L.; Yang, Y.; Cheng, X.; Hu, Z.; Cai, X.; Yang, J.; Sun, X.; et al. Sulforaphane protects against rotenone-induced neurotoxicity in vivo: Involvement of the mTOR, Nrf2, and autophagy pathways. Sci. Rep. 2016, 6, 32206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, T.R.; Gout, I.T. Functions and regulation of the 70kDa ribosomal S6 kinases. Int. J. Biochem. Cell Biol. 2011, 43, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Fujii, M.; Hou, D.X. Inhibition of lipopolysaccharide-induced cyclooxygenase-2 transcription by (6-(methylsulfinyl) hexyl isothiocyanate, a chemopreventive compound from Wasabia japonica (Miq.) Matsumura, in mouse macrophages. Biochem. Pharmacol. 2005, 70, 1772–1784. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Nishimura, A.; Shimoda, K.; Tanaka, T.; Kato, Y.; Shibata, T.; Tanaka, H.; Kurose, H.; Azuma, Y.-T.; Ihara, H.; et al. Redox-dependent internalization of the purinergic P2Y6 receptor limits colitis progression. Sci. Signal. 2022, 15, eabj0644. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimoyama, M.; Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Hosaka, K. 6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells. Curr. Issues Mol. Biol. 2022, 44, 2915-2922. https://doi.org/10.3390/cimb44070201
Shimoyama M, Hosokawa Y, Hosokawa I, Ozaki K, Hosaka K. 6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells. Current Issues in Molecular Biology. 2022; 44(7):2915-2922. https://doi.org/10.3390/cimb44070201
Chicago/Turabian StyleShimoyama, Masahiro, Yoshitaka Hosokawa, Ikuko Hosokawa, Kazumi Ozaki, and Keiichi Hosaka. 2022. "6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells" Current Issues in Molecular Biology 44, no. 7: 2915-2922. https://doi.org/10.3390/cimb44070201
APA StyleShimoyama, M., Hosokawa, Y., Hosokawa, I., Ozaki, K., & Hosaka, K. (2022). 6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells. Current Issues in Molecular Biology, 44(7), 2915-2922. https://doi.org/10.3390/cimb44070201