
Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Stool Samples
2.3. Pathogen Identification
2.4. 16S rRNA Library Preparation and Sequencing
2.5. Bioinformatics and Statistics
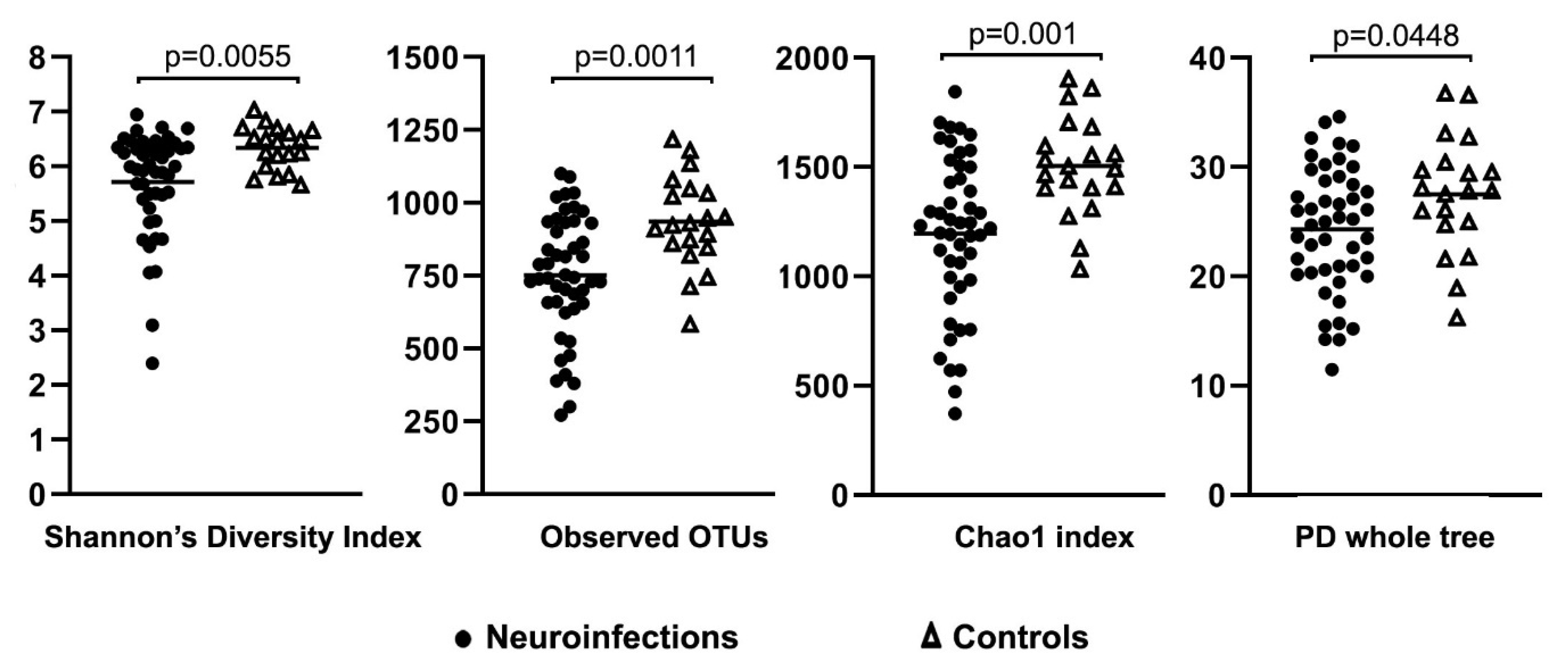
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheperjans, F.; Aho, V.; Pereira, P.A.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Yue, Y.; He, T.; Huang, C.; Qu, B.; Lv, W.; Lai, H.Y. The Association Between the Gut Microbiota and Parkinson’s Disease, a Meta-Analysis. Front. Aging Neurosci. 2021, 13, 636545. [Google Scholar] [CrossRef] [PubMed]
- van Olst, L.; Roks, S.J.M.; Kamermans, A.; Verhaar, B.J.H.; van der Geest, A.M.; Muller, M.; van der Flier, W.M.; de Vries, H.E. Contribution of Gut Microbiota to Immunological Changes in Alzheimer’s Disease. Front. Immunol. 2021, 12, 683068. [Google Scholar] [CrossRef] [PubMed]
- Miyake, S.; Kim, S.; Suda, W.; Oshima, K.; Nakamura, M.; Matsuoka, T.; Chihara, N.; Tomita, A.; Sato, W.; Kim, S.W.; et al. Dysbiosis in the Gut Microbiota of Patients with Multiple Sclerosis, with a Striking Depletion of Species Belonging to Clostridia XIVa and IV Clusters. PLoS ONE 2015, 10, e0137429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantoni, C.; Lin, Q.; Dorsett, Y.; Ghezzi, L.; Liu, Z.; Pan, Y.; Chen, K.; Han, Y.; Li, Z.; Xiao, H.; et al. Alterations of host-gut microbiome interactions in multiple sclerosis. EBioMedicine 2022, 76, 103798. [Google Scholar] [CrossRef]
- McGuinness, A.J.; Davis, J.A.; Dawson, S.L.; Loughman, A.; Collier, F.; O’Hely, M.; Simpson, C.A.; Green, J.; Marx, W.; Hair, C.; et al. A systematic review of gut microbiota composition in observational studies of major depressive disorder, bipolar disorder and schizophrenia. Mol. Psychiatry 2022, 27, 1920–1935. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; Narbad, A.; Rodriguez, J.M. Autism Spectrum Disorder Associated With Gut Microbiota at Immune, Metabolomic, and Neuroactive Level. Front. Neurosci. 2020, 14, 578666. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Zhou, Y.; Wang, D.; Liu, X.; Li, L.; Wang, T.; Zhang, Y.; Jiang, M.; Tang, H.; et al. Gut Microbiota Changes and Their Relationship with Inflammation in Patients with Acute and Chronic Insomnia. Nat. Sci. Sleep 2020, 12, 895–905. [Google Scholar] [CrossRef]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Engen, P.A.; Keshavarzian, A. Circadian Rhythm and the Gut Microbiome. Int. Rev. Neurobiol. 2016, 131, 193–205. [Google Scholar]
- Wang, Y.; Wang, Z.; Wang, Y.; Li, F.; Jia, J.; Song, X.; Qin, S.; Wang, R.; Jin, F.; Kitazato, K.; et al. The Gut-Microglia Connection: Implications for Central Nervous System Diseases. Front. Immunol. 2018, 9, 2325. [Google Scholar] [CrossRef] [Green Version]
- Gubert, C.; Gasparotto, J.; Morais, L.M. Convergent pathways of the gut microbiota-brain axis and neurodegenerative disorders. Gastroenterol. Rep. 2022, 10, goac017. [Google Scholar] [CrossRef] [PubMed]
- Herken, J.; Bang, C.; Ruhlemann, M.C.; Finke, C.; Klag, J.; Franke, A.; Pruss, H. Normal gut microbiome in NMDA receptor encephalitis. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E.; Quan, P.L.; Lipkin, W.I. Viral Encephalitis of Unknown Cause: Current Perspective and Recent Advances. Viruses 2017, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Potharaju, N.R. Incidence Rate of Acute Encephalitis Syndrome without Specific Treatment in India and Nepal. Indian J. Community Med. 2012, 37, 240–251. [Google Scholar] [CrossRef]
- Glaser, C.A.; Gilliam, S.; Schnurr, D.; Forghani, B.; Honarmand, S.; Khetsuriani, N.; Fischer, M.; Cossen, C.K.; Anderson, L.J. In search of encephalitis etiologies: Diagnostic challenges in the California Encephalitis Project, 1998–2000. Clin. Infect. Dis. 2003, 36, 731–742. [Google Scholar] [CrossRef]
- Perlejewski, K.; Bukowska-Osko, I.; Rydzanicz, M.; Pawelczyk, A.; Caraballo Corts, K.; Osuch, S.; Paciorek, M.; Dzieciatkowski, T.; Radkowski, M.; Laskus, T. Next-generation sequencing in the diagnosis of viral encephalitis: Sensitivity and clinical limitations. Sci. Rep. 2020, 10, 16173. [Google Scholar] [CrossRef]
- Glaser, C.A.; Honarmand, S.; Anderson, L.J.; Schnurr, D.P.; Forghani, B.; Cossen, C.K.; Schuster, F.L.; Christie, L.J.; Tureen, J.H. Beyond viruses: Clinical profiles and etiologies associated with encephalitis. Clin. Infect. Dis. 2006, 43, 1565–1577. [Google Scholar] [CrossRef] [Green Version]
- Granerod, J.; Ambrose, H.E.; Davies, N.W.S.; Clewley, J.P.; Walsh, A.L.; Morgan, D.; Cunningham, R.; Zuckerman, M.; Mutton, K.J.; Solomon, T.; et al. Causes of encephalitis and differences in their clinical presentations in England: A multicentre, population-based prospective study. Lancet Infect. Dis. 2010, 10, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 2 January 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronesty, E. Ea-Utils: “Command-Line Tools for Processing Biological Sequencing Data”. 2011. Available online: https://github.com/ExpressionAnalysis/ea-utils (accessed on 2 January 2022).
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Dabdoub, S.M.; Fellows, M.L.; Paropkari, A.D.; Mason, M.R.; Huja, S.S.; Tsigarida, A.A.; Kumar, P.S. PhyloToAST: Bioinformatics tools for species-level analysis and visualization of complex microbial datasets. Sci. Rep. 2016, 6, 29123. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noori, M.S.; Courreges, M.C.; Bergmeier, S.C.; McCall, K.D.; Goetz, D.J. Modulation of LPS-induced inflammatory cytokine production by a novel glycogen synthase kinase-3 inhibitor. Eur. J. Pharmacol. 2020, 883, 173340. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. Roles of Toll-like receptors in innate immune responses. Genes Cells 2001, 6, 733–742. [Google Scholar] [CrossRef]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Lu, R.; Martin, T.A.; Jiang, W.G. The role of claudin-5 in blood-brain barrier (BBB) and brain metastases (review). Mol. Med. Rep. 2014, 9, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Gong, D.; Gong, X.; Wang, L.; Yu, X.; Dong, Q. Involvement of Reduced Microbial Diversity in Inflammatory Bowel Disease. Gastroenterol Res. Pract. 2016, 2016, 6951091. [Google Scholar] [CrossRef] [PubMed]
- Wehedy, E.; Shatat, I.F.; Al Khodor, S. The Human Microbiome in Chronic Kidney Disease: A Double-Edged Sword. Front. Med. 2021, 8, 790783. [Google Scholar] [CrossRef]
- Tan, C.; Ling, Z.; Huang, Y.; Cao, Y.; Liu, Q.; Cai, T.; Yuan, H.; Liu, C.; Li, Y.; Xu, K. Dysbiosis of Intestinal Microbiota Associated With Inflammation Involved in the Progression of Acute Pancreatitis. Pancreas 2015, 44, 868–875. [Google Scholar] [CrossRef]
- Han, Y.; Gong, Z.; Sun, G.; Xu, J.; Qi, C.; Sun, W.; Jiang, H.; Cao, P.; Ju, H. Dysbiosis of Gut Microbiota in Patients with Acute Myocardial Infarction. Front. Microbiol. 2021, 12, 680101. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Noel, S.; Pluznick, J.L.; Hamad, A.R.A.; Rabb, H. Gut Microbiota-Kidney Cross-Talk in Acute Kidney Injury. Semin. Nephrol. 2019, 39, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.; Liu, X.; Li, C.; Chen, C.; Lin, J.; Li, A.; An, D.; Zhou, D.; Hong, Z. Alterations in the human gut microbiome in anti-N-methyl-D-aspartate receptor encephalitis. Ann. Clin. Transl. Neurol. 2019, 6, 1771–1781. [Google Scholar] [CrossRef]
- Mirzaei, R.; Bouzari, B.; Hosseini-Fard, S.R.; Mazaheri, M.; Ahmadyousefi, Y.; Abdi, M.; Jalalifar, S.; Karimitabar, Z.; Teimoori, A.; Keyvani, H.; et al. Role of microbiota-derived short-chain fatty acids in nervous system disorders. Biomed. Pharmacother. 2021, 139, 111661. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Toth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Erny, D.; Hrabe de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Ma, L.; Wang, Z.; Liu, Y.; Long, L.; Ma, X.; Chen, H.; Chen, Z.; Lin, X.; Si, L.; et al. Clinical Features and Gut Microbial Alterations in Anti-leucine-rich Glioma-Inactivated 1 Encephalitis-A Pilot Study. Front. Neurol. 2020, 11, 585977. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Nunez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Shi, X.; Yang, J.; Zhao, Y.; Xue, L.; Xu, L.; Cai, J. Gut microbes in cardiovascular diseases and their potential therapeutic applications. Protein Cell 2021, 12, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D. The Gut Microbiome and Its Role in Obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palleja, A.; Mikkelsen, K.H.; Forslund, S.K.; Kashani, A.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Liang, S.; Feng, Q.; Zhang, C.; et al. Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 2018, 3, 1255–1265. [Google Scholar] [CrossRef]
- Kwon, Y.; Cho, Y.-S.; Lee, Y.-M.; Kim, S.-j.; Bae, J.; Jeong, S.-J. Changes to Gut Microbiota Following Systemic Antibiotic Administration in Infants. Antibiotics 2022, 11, 470. [Google Scholar] [CrossRef]
- Panda, S.; El khader, I.; Casellas, F.; Lopez Vivancos, J.; Garcia Cors, M.; Santiago, A.; Cuenca, S.; Guarner, F.; Manichanh, C. Short-term effect of antibiotics on human gut microbiota. PLoS ONE 2014, 9, e95476. [Google Scholar] [CrossRef]
- Gu, S.L.; Gong, Y.W.; Zhang, J.Y.; Chen, Y.B.; Wu, Z.J.; Xu, Q.M.; Fang, Y.H.; Wang, J.X.; Tang, L.L. Effect of the Short-Term Use of Fluoroquinolone and beta-Lactam Antibiotics on Mouse Gut Microbiota. Infect. Drug Resist. 2020, 13, 4547–4558. [Google Scholar] [CrossRef]
- Hertz, F.B.; Budding, A.E.; van der Lugt-Degen, M.; Savelkoul, P.H.; Lobner-Olesen, A.; Frimodt-Moller, N. Effects of Antibiotics on the Intestinal Microbiota of Mice. Antibiotics 2020, 9, 191. [Google Scholar] [CrossRef]
- Yao, J.; Carter, R.A.; Vuagniaux, G.; Barbier, M.; Rosch, J.W.; Rock, C.O. A Pathogen-Selective Antibiotic Minimizes Disturbance to the Microbiome. Antimicrob. Agents Chemother. 2016, 60, 4264–4273. [Google Scholar] [CrossRef] [Green Version]
- Plassais, J.; Gbikpi-Benissan, G.; Figarol, M.; Scheperjans, F.; Gorochov, G.; Derkinderen, P.; Cervino, A.C.L. Gut microbiome alpha-diversity is not a marker of Parkinson’s disease and multiple sclerosis. Brain Commun. 2021, 3, fcab113. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Ackermann, G.; Khailova, L.; Baird, C.; Heyland, D.; Kozar, R.; Lemieux, M.; Derenski, K.; King, J.; Vis-Kampen, C.; et al. Extreme Dysbiosis of the Microbiome in Critical Illness. mSphere 2016, 1, e00199-16. [Google Scholar] [CrossRef] [Green Version]
- Ravi, A.; Halstead, F.D.; Bamford, A.; Casey, A.; Thomson, N.M.; van Schaik, W.; Snelson, C.; Goulden, R.; Foster-Nyarko, E.; Savva, G.M.; et al. Loss of microbial diversity and pathogen domination of the gut microbiota in critically ill patients. Microb. Genom. 2019, 5, e000293. [Google Scholar] [CrossRef] [PubMed]
- Zaborin, A.; Smith, D.; Garfield, K.; Quensen, J.; Shakhsheer, B.; Kade, M.; Tirrell, M.; Tiedje, J.; Gilbert, J.A.; Zaborina, O.; et al. Membership and behavior of ultra-low-diversity pathogen communities present in the gut of humans during prolonged critical illness. mBio 2014, 5, e01361-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuroinfections | Number (%) | |||
|---|---|---|---|---|
| All Patients | Viral | Bacterial | Undefined | |
| Nº of patients | 47 | 26 (55.32) | 8 (17.02) | 13 (27.66) |
| Male | 23 (48.94) | 13 (50) | 4 (50) | 6 (46.15) |
| Female | 24 (51.06) | 13 (50) | 4 (50) | 7 (53.85) |
| Age, Mean | 43 | 41 | 50 | 44 |
| Symptoms and signs | Number (%) | |||
| All patients | Viral | Bacterial | Undefined | |
| Fever | 31 (65.96) | 19 (73.08) | 6 (75) | 6 (46.15) |
| Headache | 37 (78.72) | 22 (84.62) | 6 (75) | 9 (69.26) |
| Altered mental status | 11 (23.40) | 2 (7.69) | 4 (50) | 5 (38.46) |
| Loss of consciousness | 6 (12.76) | 2 (7.69) | 1 (50) | 3 (23.08) |
| Seizures of epilepsy | 4 (8.51) | 1 (3.85) | 0 | 3 (23.08) |
| Focal neurologic signs | 19 (40.43) | 13 (50) | 6 (75) | 0 |
| CSF analysis (ref. values) | Mean value/standard deviation | |||
| All patients | Viral | Bacterial | Undefined | |
| Total cell count, cells/µL (≤5) | 190/238 | 155/120 | 619/379 | 64/114 |
| % of lymphocytes | 63/30.46 | 69/27 | 20/6 | 50/37 |
| Chlorides, mmol/L (>117) | 121.72/4.15 | 121.61/3.22 | 119.00/5.61 | 123.49/3.91 |
| Protein, g/L (0.15–0.45) | 1.51/1.62 | 1.09/0.89 | 2.51/1.87 | 1.77/2.23 |
| L-Lactic acid, mmol/L (≤2.1) | 2.23/0.99 | 2.15/0.61 | 4.94/1.33 | 1.93/0.77 |
| Glucose, mmol/L (2.4–4.7) | 2.80/0.87 | 2.97/0.35 | 1.64/0.73 | 3.28/1.02 |
| Blood test (ref. values) | Mean value/standard deviation | |||
| All patients | Viral | Bacterial | Undefined | |
| WBC, ×103/µL (–10) | 10.65/4.04 | 9.05/2.72 | 14.74/3.42 | 10.87/4.48 |
| RBC, ×106/µL (4.5–5.9) | 4.56/0.57 | 4.65/0.51 | 3.99/0.75 | 4.63/0.44 |
| Platelet count, ×103/µL (150–450) | 238.48/63.80 | 233.87/52.42 | 204.50/58.14 | 265.80/70.69 |
| CRP, mg/L (<5) | 40.84/63.04 | 13.33/8.55 | 120.37/86.66 | 19.77/18.77 |
| Treatment | Number (%) | |||
| All patients | Viral | Bacterial | Undefined | |
| Antibiotics | 23 (48.94) | 9 (34.62) | 8 (100) | 6 (46.15) |
| Antiviral drugs (Aciclovir) | 15 (31.9) | 10 (38.46) | 0 | 5 (38.46) |
| Antiepileptic Drugs | 7 (14.89) | 2 (7.69) | 1 (12.50) | 4 (30.77) |
| Control group (%) | ||||
| Nº of controls | 20 | |||
| Male | 10 (50) | |||
| Female | 10 (50) | |||
| Age, Mean | 43 | |||
| Taxonomy | Controls (n = 20) vs. | |||
|---|---|---|---|---|
| Neuroinfections (n = 47) | Bacterial (n = 8) | Viral (n = 26) | ||
| Phylum | p__ Bacillota | 0.015 | 0.037 | 0.037 |
| p__ Actinomycetota | 0.037 | |||
| p__ Verrucomicrobiota | 0.030 | |||
| Class | p__ Bacillota;c__Clostridia | 0.026 | 0.013 | 0.026 |
| p__ Actinomycetota;c__Coriobacteriia | 0.013 | |||
| p__ Verrucomicrobiota;c__Verrucomicrobiae | 0.039 | |||
| Order | p__ Bacillota;c__Clostridia;o__Clostridiales | 0.043 | 0.021 | |
| p__ Actinomycetota;c__Coriobacteriia; o__Coriobacteriales | 0.021 | |||
| Family | p__ Bacillota;c__Clostridia;o__Clostridiales;f__Eubacteriaceae | 0.045 | ||
| p__ Verrucomicrobiota;c__Verrucomicrobiae; o__Verrucomicrobiales;f__Verrucomicrobiaceae | 0.045 | |||
| Genus | p__ Bacillota;c__Clostridia;o__Clostridiales;f__Clostridiaceae;g__Clostridium | 0.025 | 0.040 | |
| p__ Bacillota;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Anaerostipes | 0.025 | 0.040 | ||
| p__ Bacillota;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Lachnobacterium | 0.025 | 0.040 | ||
| p__ Bacillota;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Lachnospira | 0.025 | 0.040 | ||
| p__ Bacillota;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Roseburia | 0.025 | 0.040 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grochowska, M.; Laskus, T.; Paciorek, M.; Pollak, A.; Lechowicz, U.; Makowiecki, M.; Horban, A.; Radkowski, M.; Perlejewski, K. Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity. Curr. Issues Mol. Biol. 2022, 44, 2903-2914. https://doi.org/10.3390/cimb44070200
Grochowska M, Laskus T, Paciorek M, Pollak A, Lechowicz U, Makowiecki M, Horban A, Radkowski M, Perlejewski K. Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity. Current Issues in Molecular Biology. 2022; 44(7):2903-2914. https://doi.org/10.3390/cimb44070200
Chicago/Turabian StyleGrochowska, Marta, Tomasz Laskus, Marcin Paciorek, Agnieszka Pollak, Urszula Lechowicz, Michał Makowiecki, Andrzej Horban, Marek Radkowski, and Karol Perlejewski. 2022. "Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity" Current Issues in Molecular Biology 44, no. 7: 2903-2914. https://doi.org/10.3390/cimb44070200
APA StyleGrochowska, M., Laskus, T., Paciorek, M., Pollak, A., Lechowicz, U., Makowiecki, M., Horban, A., Radkowski, M., & Perlejewski, K. (2022). Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity. Current Issues in Molecular Biology, 44(7), 2903-2914. https://doi.org/10.3390/cimb44070200