
Heterogeneous Immunolocalisation of Zinc Transporters ZIP6, ZIP10 and ZIP14 in Human Normo- and Asthenozoospermic Spermatozoa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Preparation of Samples
2.2. Immunofluorescence
2.3. Statistics
3. Results
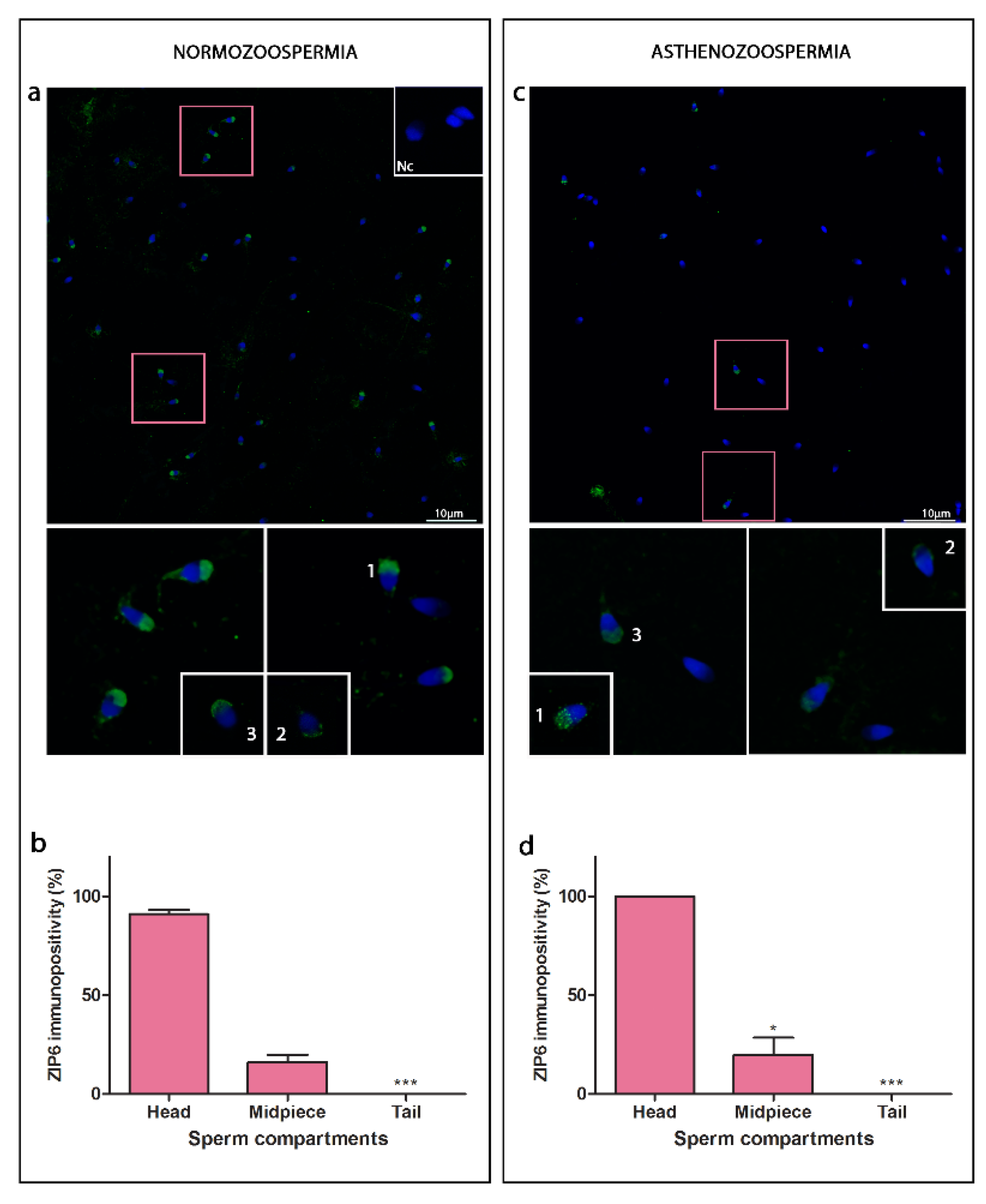
3.1. ZIP6
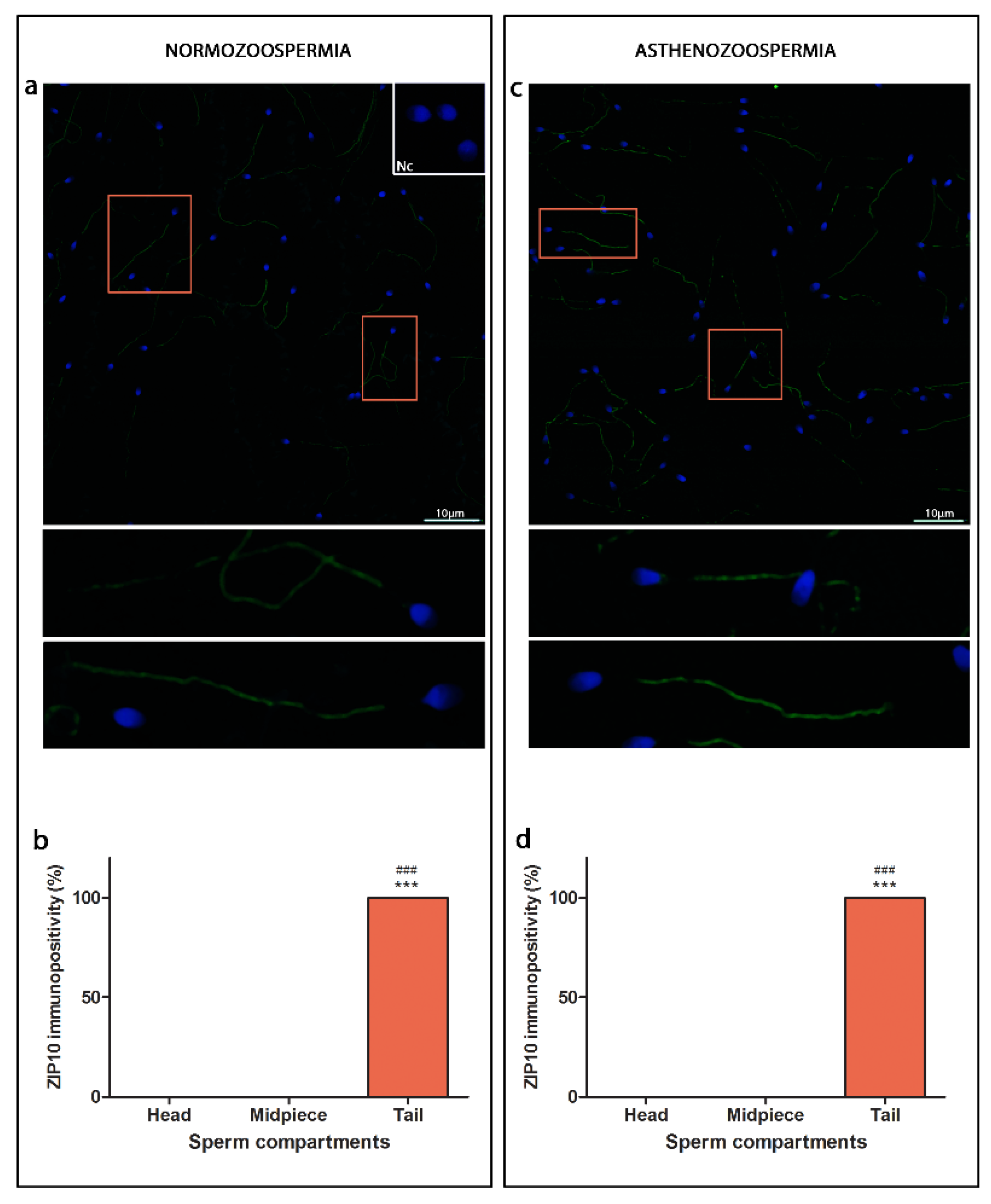
3.2. ZIP10
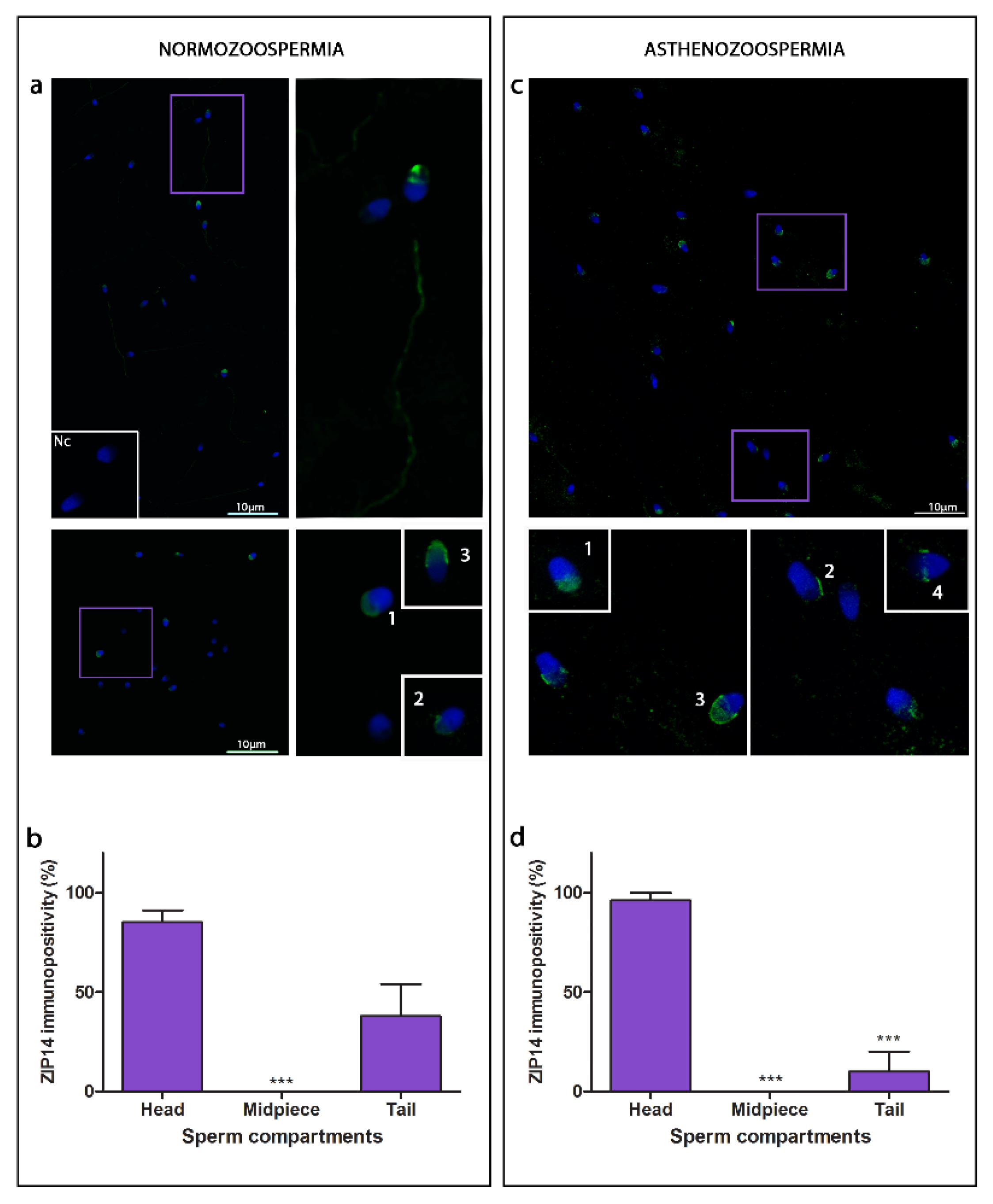
3.3. ZIP14
4. Discussion
4.1. ZIPs Are Heterogeneously Expressed in Sperm Cells; Lower Level of Overall Immunostaining of ZIP6 Is Found in Asthenozoospermia
4.2. ZIPs Show Different Compartmental Localisation, ZIP6 and ZIP14 Are Found in the Head, While ZIP10 Are Located Entirely along the Sperm Cell Tail
4.3. Specific Head Sub-Compartment Localisation of ZIP6 and ZIP14
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kambe, T.; Nishito, Y.; Fukue, K. Chapter 23—Zinc transporters in health and disease. In Molecular, Genetic, and Nutritional Aspects of Major and Trace Minerals; Collins, J.F., Ed.; Academic Press: Boston, MA, USA, 2017; pp. 283–291. ISBN 978-0-12-802168-2. [Google Scholar]
- Kerns, K.; Zigo, M.; Sutovsky, P. Zinc: A necessary ion for mammalian sperm fertilization competency. Int. J. Mol. Sci. 2018, 19, 4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, B.J.; Bradwell, A.R. Identification of the serum binding proteins for iron, zinc, cadmium, nickel, and calcium. Clin. Chem. 1983, 29, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Garolla, A.; Cosci, I.; Menegazzo, M.; Ferigo, M.; Gandin, V.; De Toni, L. Role of zinc trafficking in male fertility: From germ to sperm. Hum. Reprod. 2014, 29, 1134–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vickram, S.; Rohini, K.; Srinivasan, S.; Veenakumari, D.N.; Archana, K.; Anbarasu, K.; Jeyanthi, P.; Thanigaivel, S.; Gulothungan, G.; Rajendiran, N.; et al. Role of zinc (Zn) in human reproduction: A journey from initial spermatogenesis to childbirth. Int. J. Mol. Sci. 2021, 22, 2188. [Google Scholar] [CrossRef] [PubMed]
- Croxford, T.P.; McCormick, N.H.; Kelleher, S.L. Moderate zinc deficiency reduces testicular Zip6 and Zip10 abundance and impairs spermatogenesis in mice. J. Nutr. 2011, 141, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.R. Critical role of zinc as either an antioxidant or a prooxidant in cellular systems. Oxidative Med. Cell. Longev. 2018, 2018, 9156285. [Google Scholar] [CrossRef] [Green Version]
- Fallah, A.; Mohammad-Hasani, A.; Colagar, A.H. Zinc is an essential element for male fertility: A review of Zn roles in men’s health, germination, sperm quality, and fertilization. J. Reprod. Infertil. 2018, 19, 69–81. [Google Scholar] [PubMed]
- Bettger, W.J.; O’Dell, B.L. A critical physiological role of zinc in the structure and function of biomembranes. Life Sci. 1981, 28, 1425–1438. [Google Scholar] [CrossRef]
- Xu, S.; Wu, Y.; Chen, Y.; Lu, W.; Wang, Y.X.; Gao, B.; Zhang, J. Environmental metal exposure, seminal plasma metabolome and semen quality: Evidence from Chinese reproductive-aged men. Sci. Total Environ. 2022, 838, 155860. [Google Scholar] [CrossRef]
- Garolla, A.; Petre, G.C.; Francini-Pesenti, F.; De Toni, L.; Vitagliano, A.; Di Nisio, A.; Grande, G.; Foresta, C. Systematic Review and Critical Analysis on Dietary Supplements for Male Infertility: From a Blend of Ingredients to a Rationale Strategy. Front. Endocrinol. 2022, 12, 824078. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Yamaguchi-Iwai, Y.; Sasaki, R.; Nagao, M. Overview of mammalian zinc transporters. Cell. Mol. Life Sci. 2004, 61, 49–68. [Google Scholar] [CrossRef]
- Zhu, X.; Yu, C.; Wu, W.; Shi, L.; Jiang, C.; Wang, L.; Ding, Z.; Liu, Y. Zinc transporter ZIP12 maintains zinc homeostasis and protects spermatogonia from oxidative stress during spermatogenesis. Reprod. Biol. Endocrinol. 2022, 20, 17. [Google Scholar] [CrossRef]
- Atig, F.; Raffa, M.; Habib, B.A.; Kerkeni, A.; Saad, A.; Ajina, M. Impact of seminal trace element and glutathione levels on semen quality of Tunisian infertile men. BMC Urol. 2012, 12, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoltenberg, M.; Sørensen, M.B.; Danscher, G. Histochemical demonstration of zinc ions in ejaculated human semen. Int. J. Androl. 1997, 20, 229–236. [Google Scholar] [CrossRef] [PubMed]
- WHO. Laboratory Manual for the Examination and Processing of Human Semen. Available online: https://www.who.int/publications-detail-redirect/9789240030787 (accessed on 11 April 2022).
- Protic, I.; Golic, I.; Aleksic, M.; Vidakovic, S.; Korac, B.; Korac, A. Presence of acetylated α-tubulin in human sperm nuclei: A contributor to sperm heterogeneity. Med. Hypotheses 2022, 161, 110800. [Google Scholar] [CrossRef]
- Golić, I.; Aleksić, M.; Lazarević, A.; Bogdanović, M.; Jonić, S.; Korać, A. Methods for studying the localization of mitochondrial complexes III and IV by immunofluorescent and immunogold microscopy. Arch. Biol. Sci. 2016, 68, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Shen, J.; Wang, Y.; Pan, C.; Pang, W.; Diao, H.; Dong, W. Boar seminal plasma exosomes maintain sperm function by infiltrating into the sperm membrane. Oncotarget 2016, 7, 58832–58847. [Google Scholar] [CrossRef] [PubMed]
- Murdica, V.; Giacomini, E.; Alteri, A.; Bartolacci, A.; Cermisoni, G.C.; Zarovni, N.; Papaleo, E.; Montorsi, F.; Salonia, A.; Viganò, P.; et al. Seminal plasma of men with severe asthenozoospermia contain exosomes that affect spermatozoa motility and capacitation. Fertil. Steril. 2019, 111, 897–908.e2. [Google Scholar] [CrossRef] [PubMed]
- Bedwal, R.S.; Bahuguna, A. Zinc, copper and selenium in reproduction. Experientia 1994, 50, 626–640. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, B.J.; Kang, J.; Nam, T.S.; Lim, J.M.; Kim, H.T.; Park, J.K.; Kim, Y.G.; Chae, S.-W.; Kim, U.-H. Ca2+ signaling tools acquired from prostasomes are required for progesterone-induced sperm motility. Sci. Signal. 2011, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Taravati, A.; Tohidi, F. Association between seminal plasma zinc level and asthenozoospermia: A meta-analysis study. Andrologia 2016, 48, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Kvist, U.; Björndahl, L.; Roomans, G.M.; Lindholmer, C. Nuclear zinc in human epididymal and ejaculated spermatozoa. Acta Physiol. Scand. 1985, 125, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc ion flux during mammalian sperm capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef] [Green Version]
- Henkel, R.; Bittner, J.; Weber, R.; Hüther, F.; Miska, W. Relevance of zinc in human sperm flagella and its relation to motility. Fertil. Steril. 1999, 71, 1138–1143. [Google Scholar] [CrossRef]
- Bjorndahl, L.; Kjellberg, S.; Roomans, G.M.; Kvist, U. The human sperm nucleus takes up zinc at ejaculation. Int. J. Androl. 1986, 9, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Mirnamniha, M.; Faroughi, F.; Tahmasbpour, E.; Ebrahimi, P.; Harchegani, A.B. An overview on role of some trace elements in human reproductive health, sperm function and fertilization process. Rev. Environ. Health 2019, 34, 339–348. [Google Scholar] [CrossRef]
- Colleu, D.; Lescoat, D.; Boujard, D.; Le Lannou, D. Human spermatozoal nuclear maturity in normozoospermia and asthenozoospermia. Arch. Androl. 1988, 21, 155–162. [Google Scholar] [CrossRef]
- Kumar, D.; Kalthur, G.; Mascarenhas, C.; Kumar, P.; Adiga, S.K. Ejaculate fractions of asthenozoospermic and teratozoospermic patients have differences in the sperm DNA integrity. Andrologia 2011, 43, 416–421. [Google Scholar] [CrossRef]
- Mortimer, D. The functional anatomy of the human spermatozoon: Relating ultrastructure and function. Mol. Hum. Reprod. 2018, 24, 567–592. [Google Scholar] [CrossRef] [PubMed]
- Björndahl, L.; Kvist, U. Importance of zinc for human sperm head-tail connection. Acta Physiol. Scand. 1982, 116, 51–55. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Head of the Spermatozoa | ||||
|---|---|---|---|---|
| Anterior Head Region (%) | Posterior Head Region (%) | |||
| Acorn-Like Pattern | Crescent-Like Pattern | Acorn/Crescent-Like Pattern | ||
| Normozoospermia | 64.61 ± 6.18 | 19.81 ± 4.09 | 12.68 ± 2.90 | 33.45 ± 7.64 |
| Asthenozoospermia | 71.32 ± 9.95 | 21.54 ± 8.29 | 1.52 ± 1.52 ** | 56.49 ± 14.84 |
| Head of the Spermatozoa | ||||
|---|---|---|---|---|
| Anterior Head Region (%) | Posterior Head Region (%) | |||
| Acorn-Like pattern | Crescent-Like Pattern | Acorn/Crescent like-Pattern | ||
| Normozoospermia | 68.08 ± 2.58 | 18.50 ± 2.91 | 14.66 ± 3.22 | 0.00 |
| Asthenozoospermia | 17.57 ± 6.57 *** | 67.63 ± 9.36 *** | 2.38 ± 2.38 ** | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Protic, I.; Golic, I.; Vidakovic, S.; Korac, B.; Korac, A. Heterogeneous Immunolocalisation of Zinc Transporters ZIP6, ZIP10 and ZIP14 in Human Normo- and Asthenozoospermic Spermatozoa. Curr. Issues Mol. Biol. 2022, 44, 3444-3454. https://doi.org/10.3390/cimb44080237
Protic I, Golic I, Vidakovic S, Korac B, Korac A. Heterogeneous Immunolocalisation of Zinc Transporters ZIP6, ZIP10 and ZIP14 in Human Normo- and Asthenozoospermic Spermatozoa. Current Issues in Molecular Biology. 2022; 44(8):3444-3454. https://doi.org/10.3390/cimb44080237
Chicago/Turabian StyleProtic, Isidora, Igor Golic, Snezana Vidakovic, Bato Korac, and Aleksandra Korac. 2022. "Heterogeneous Immunolocalisation of Zinc Transporters ZIP6, ZIP10 and ZIP14 in Human Normo- and Asthenozoospermic Spermatozoa" Current Issues in Molecular Biology 44, no. 8: 3444-3454. https://doi.org/10.3390/cimb44080237
APA StyleProtic, I., Golic, I., Vidakovic, S., Korac, B., & Korac, A. (2022). Heterogeneous Immunolocalisation of Zinc Transporters ZIP6, ZIP10 and ZIP14 in Human Normo- and Asthenozoospermic Spermatozoa. Current Issues in Molecular Biology, 44(8), 3444-3454. https://doi.org/10.3390/cimb44080237