
Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
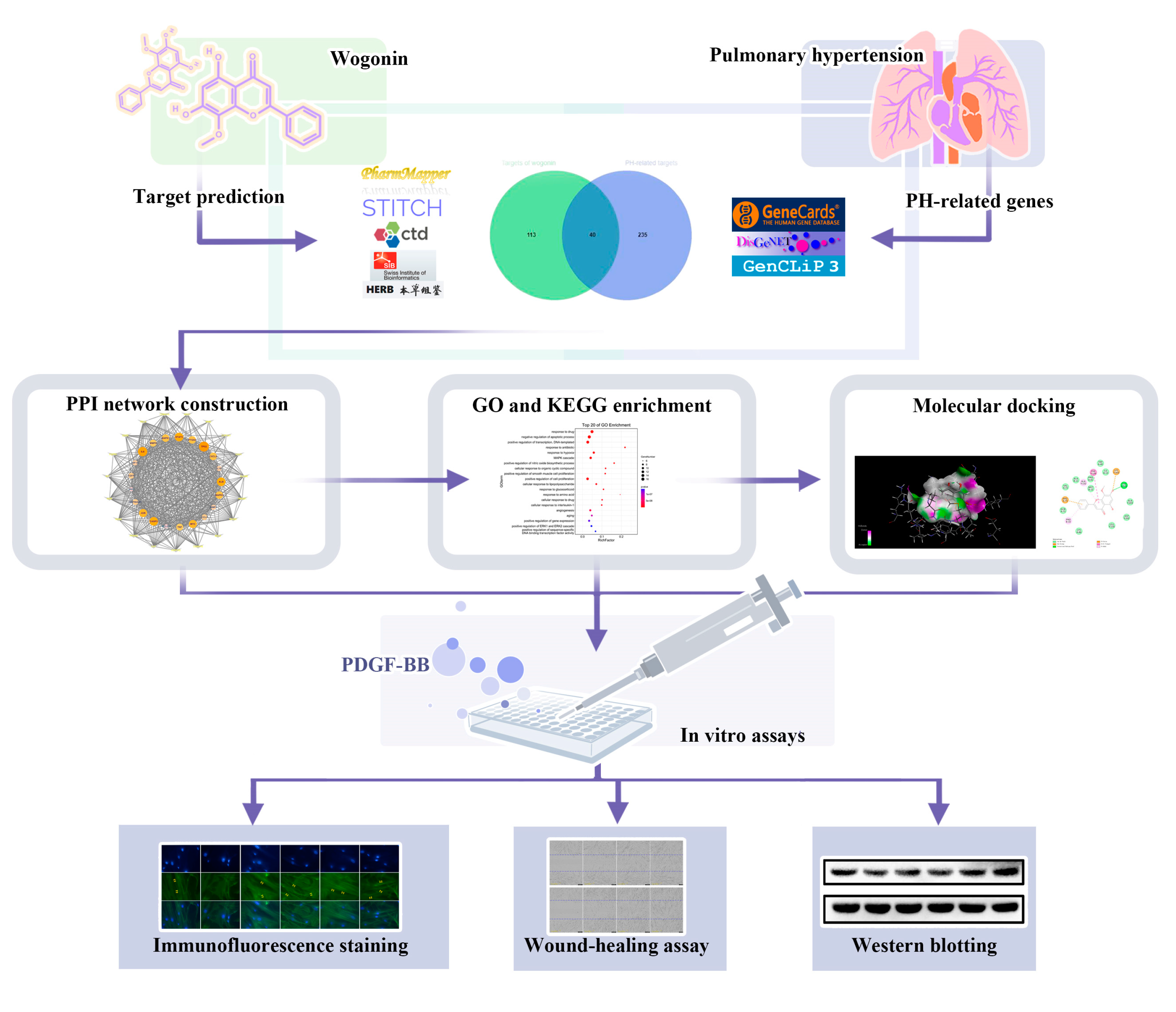
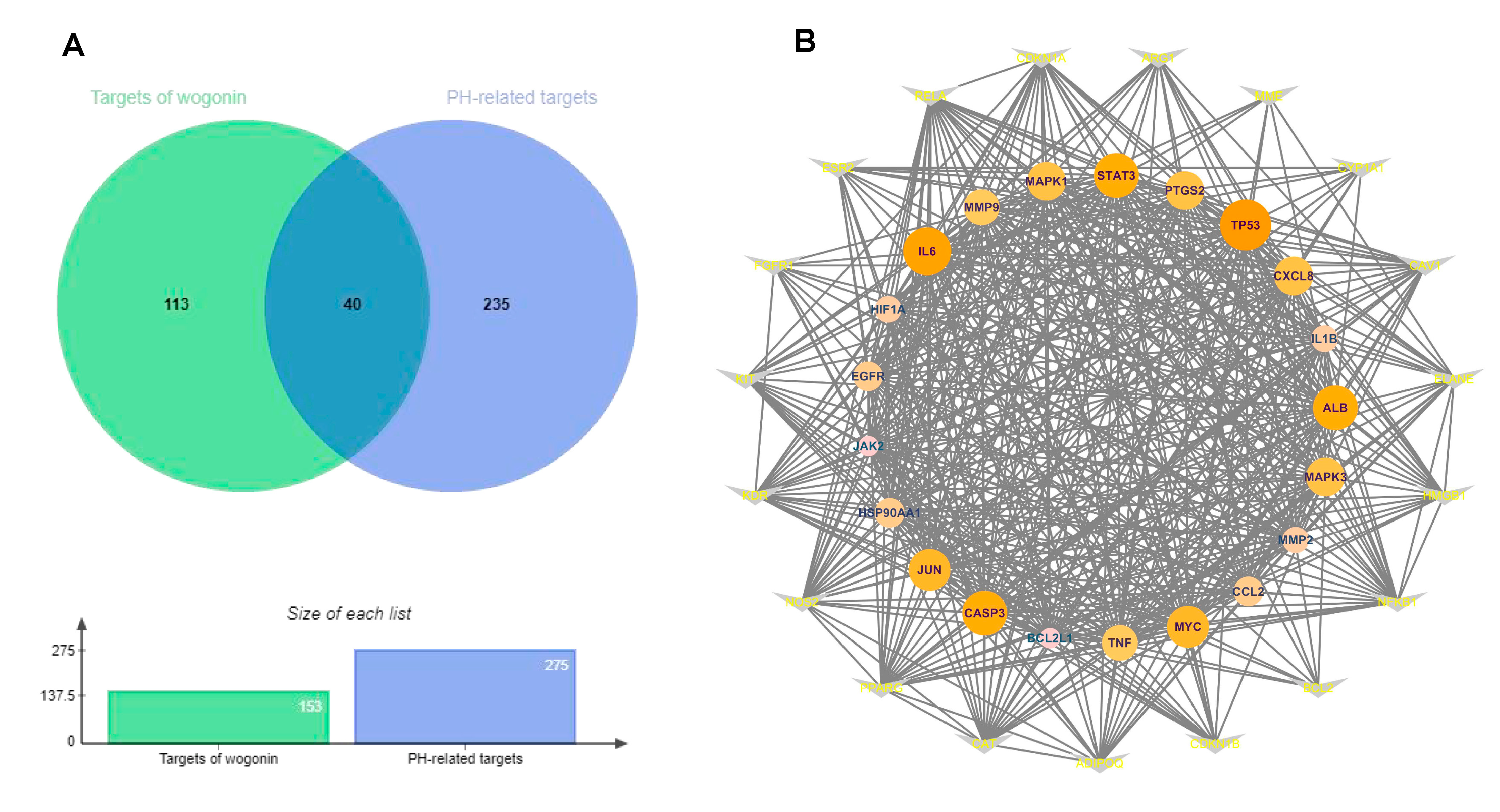
2.2. Collection of Target Genes
2.3. PPI Network Construction
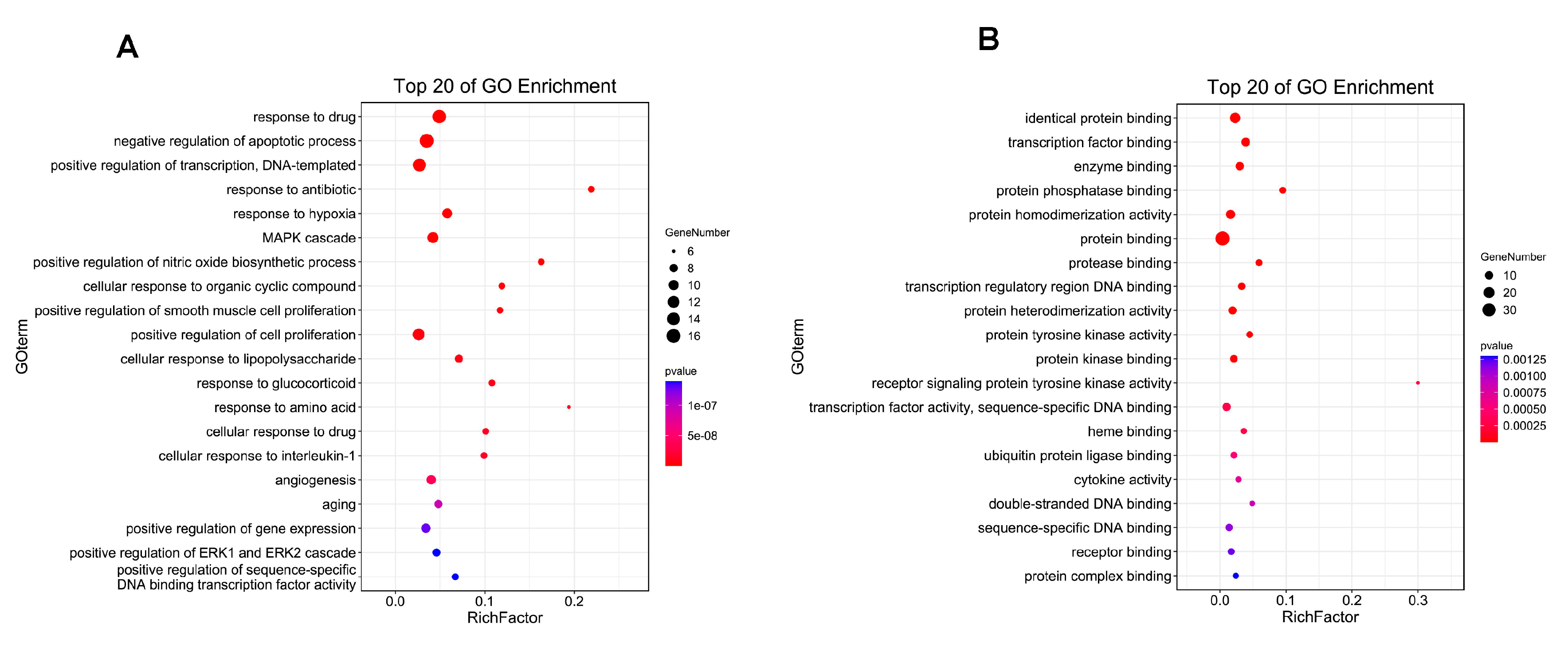
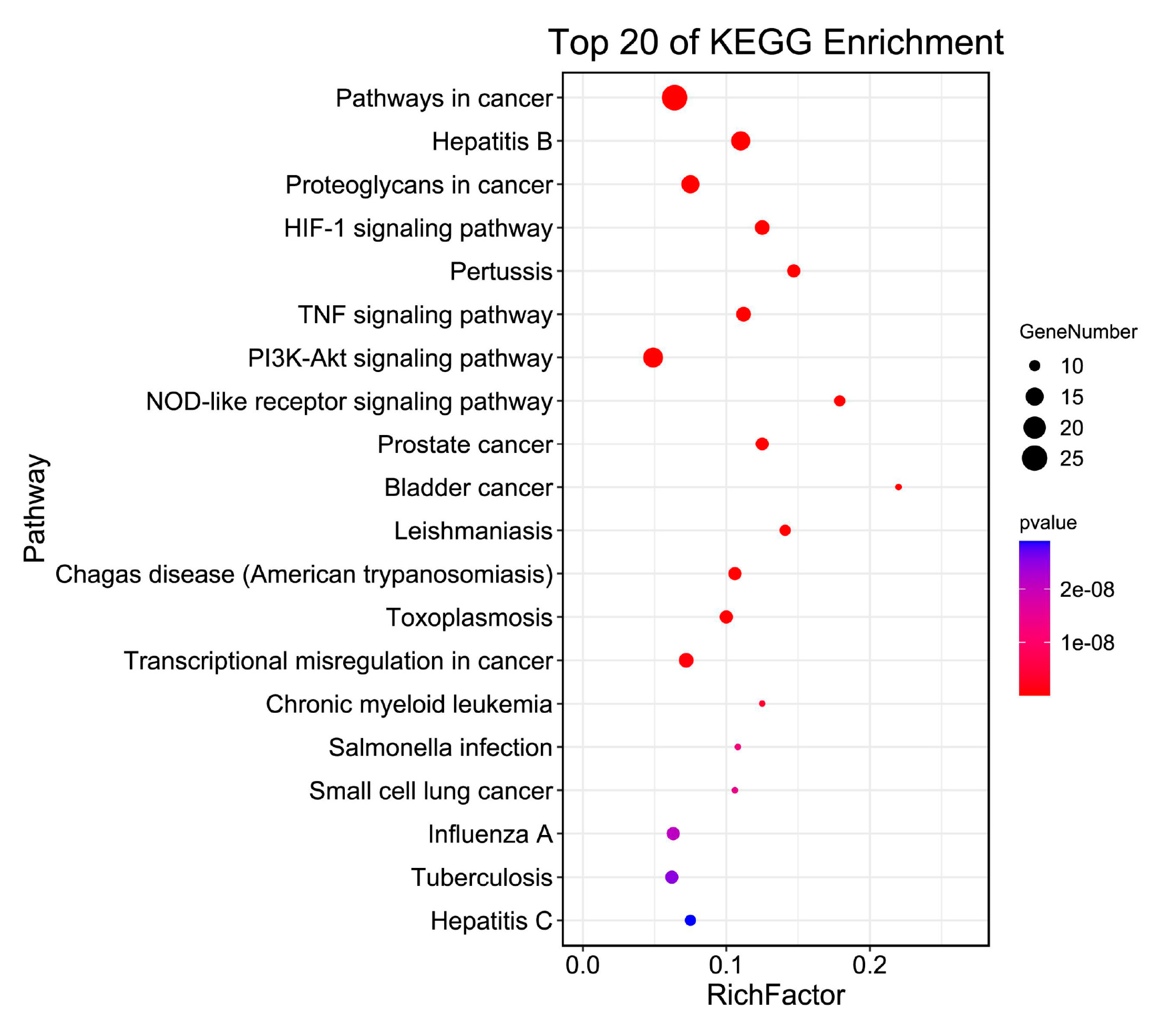
2.4. GO and KEGG Enrichment
2.5. Molecular Docking
2.6. Cell Culture
2.7. Cell Viability Assay
2.8. Immunofluorescence Staining
2.9. Wound-Healing Assay
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
3.1. PPI Analysis of Targets of Wogonin against PH
3.2. Gene Ontology Analysis
3.3. Pathway Enrichment Analysis
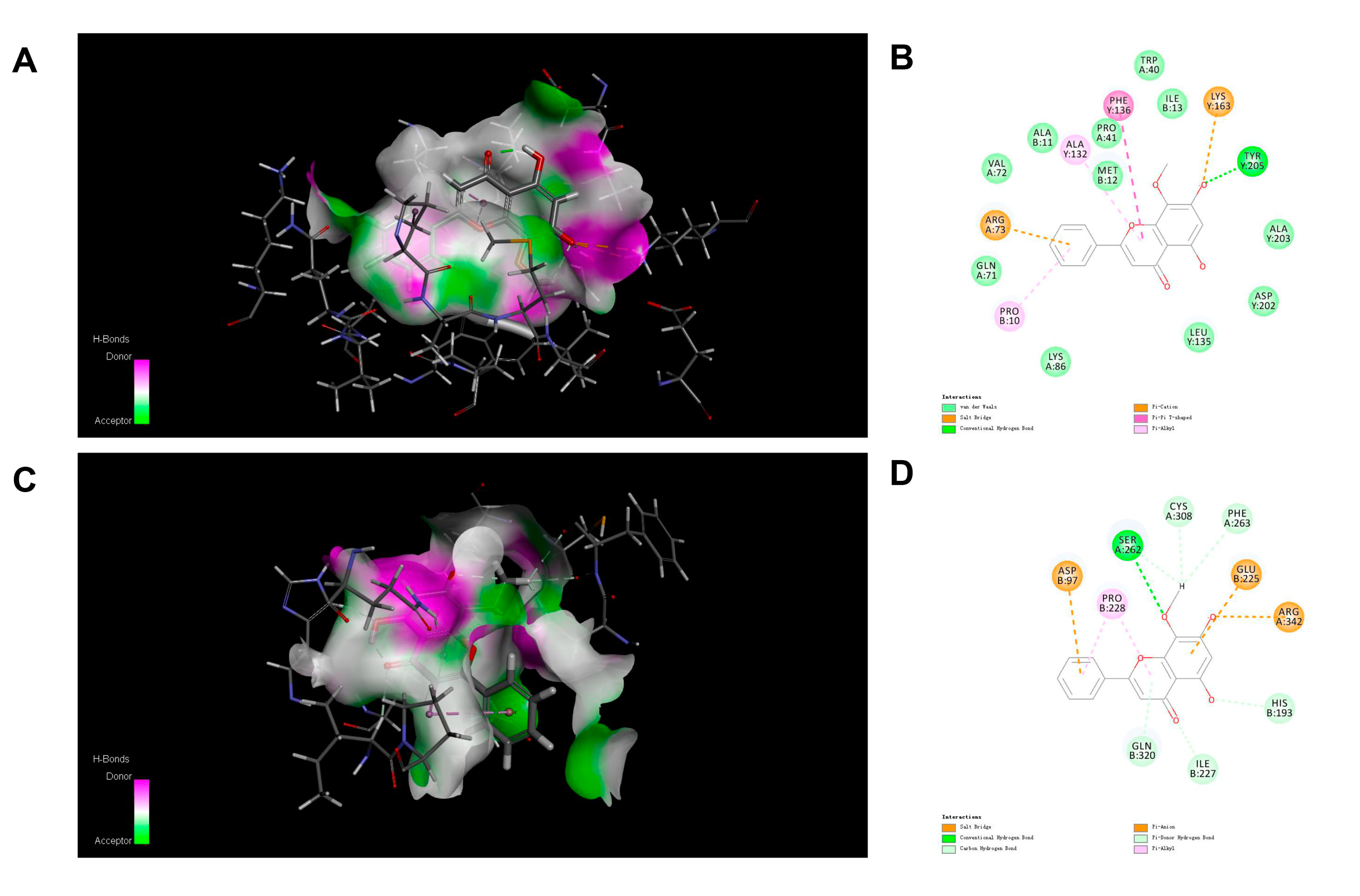
3.4. Molecular Docking
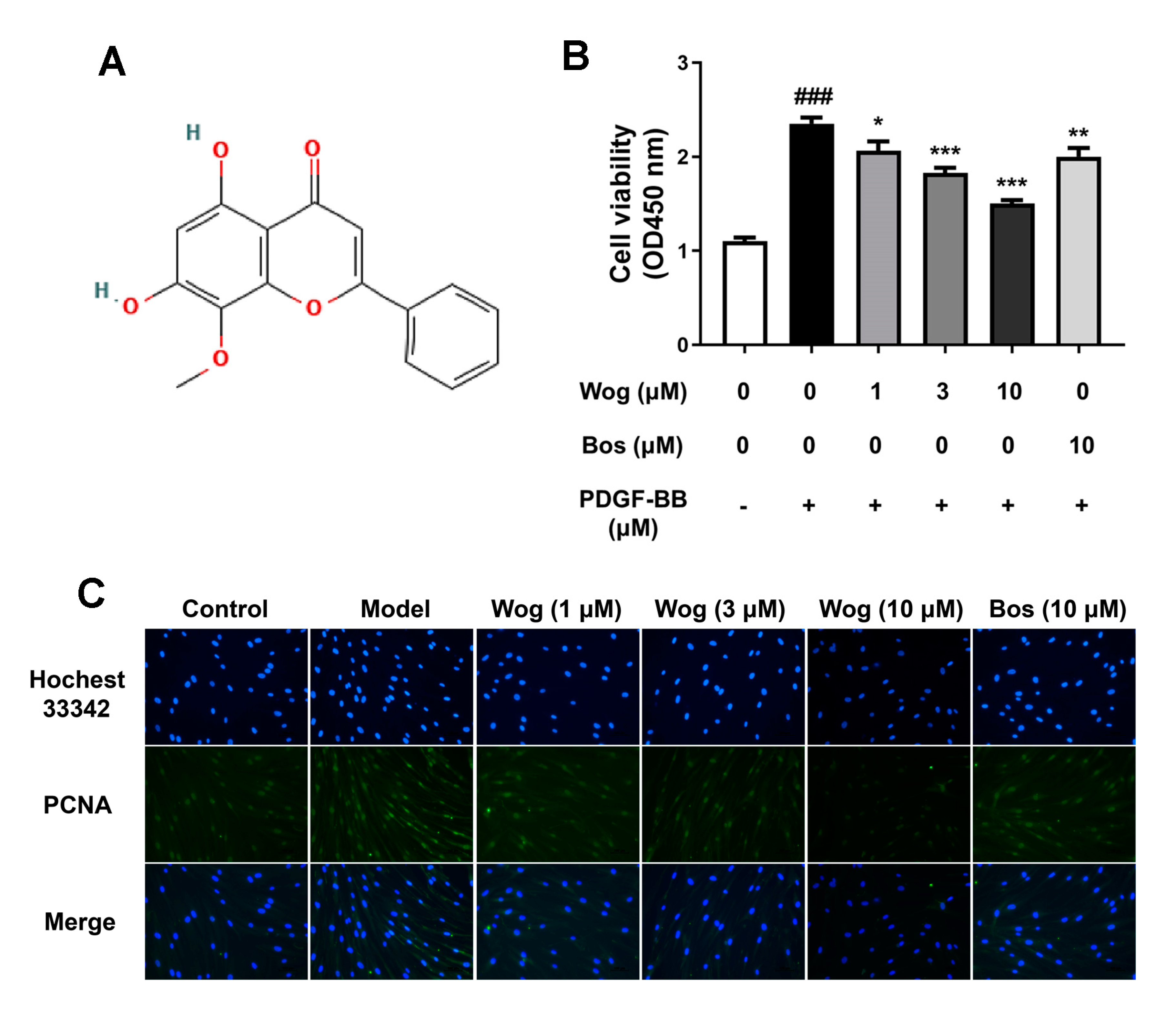
3.5. Wogonin Inhibits PDGF-BB-Mediated Proliferation of PASMCs
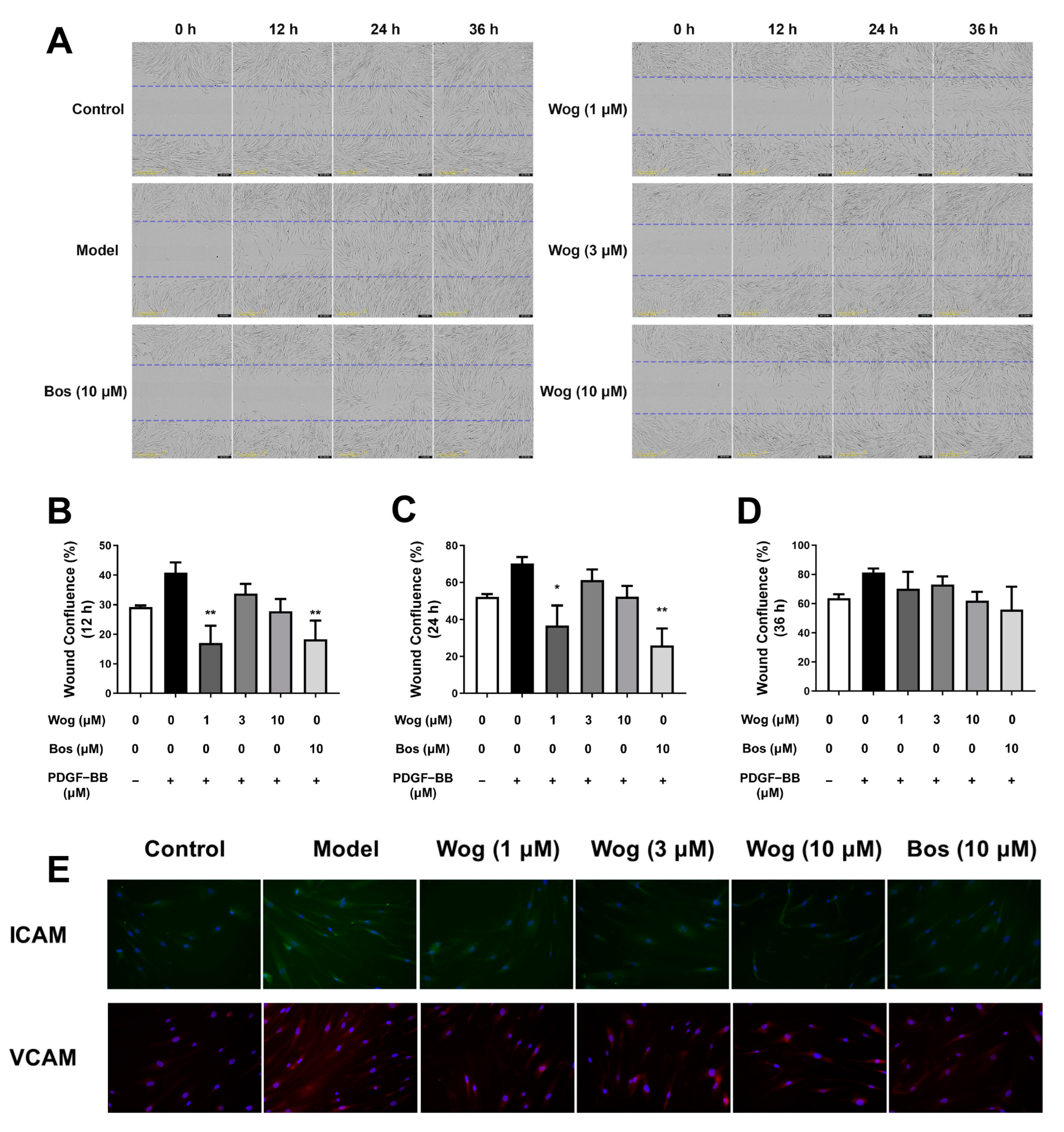
3.6. Wogonin Inhibits Migration of PASMCs
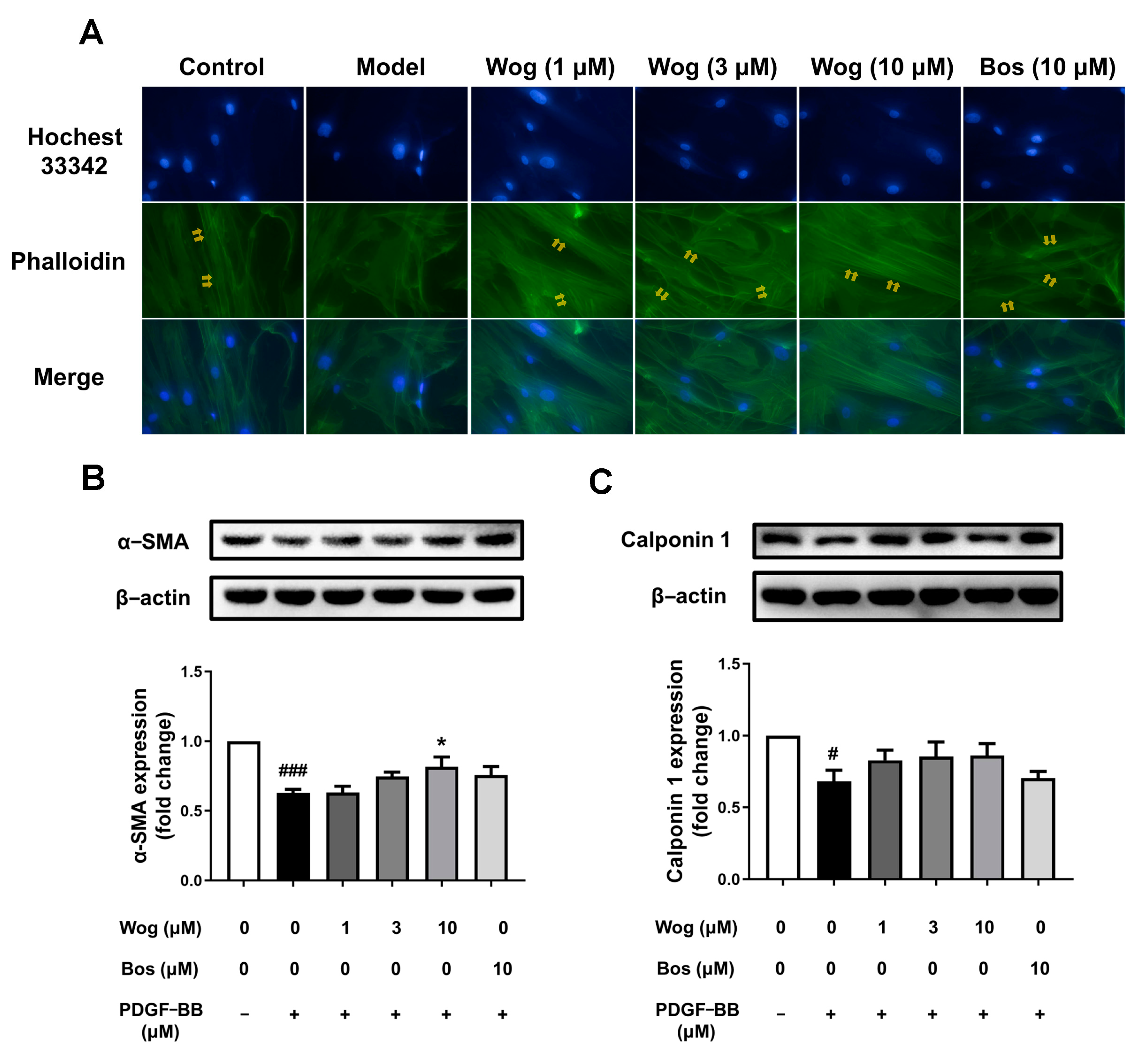
3.7. Effect of Wogonin on Phenotypic Switch of PASMCs
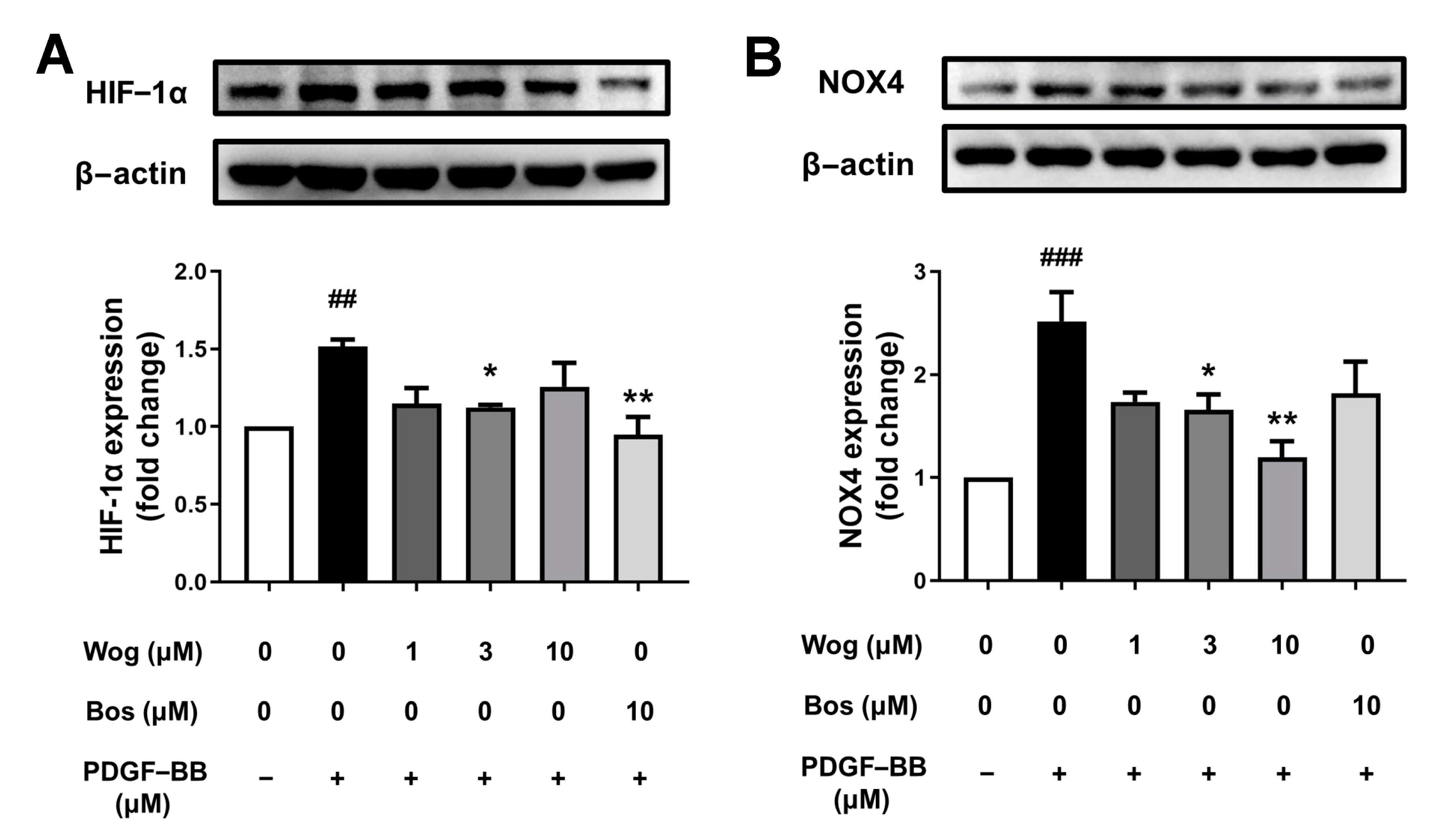
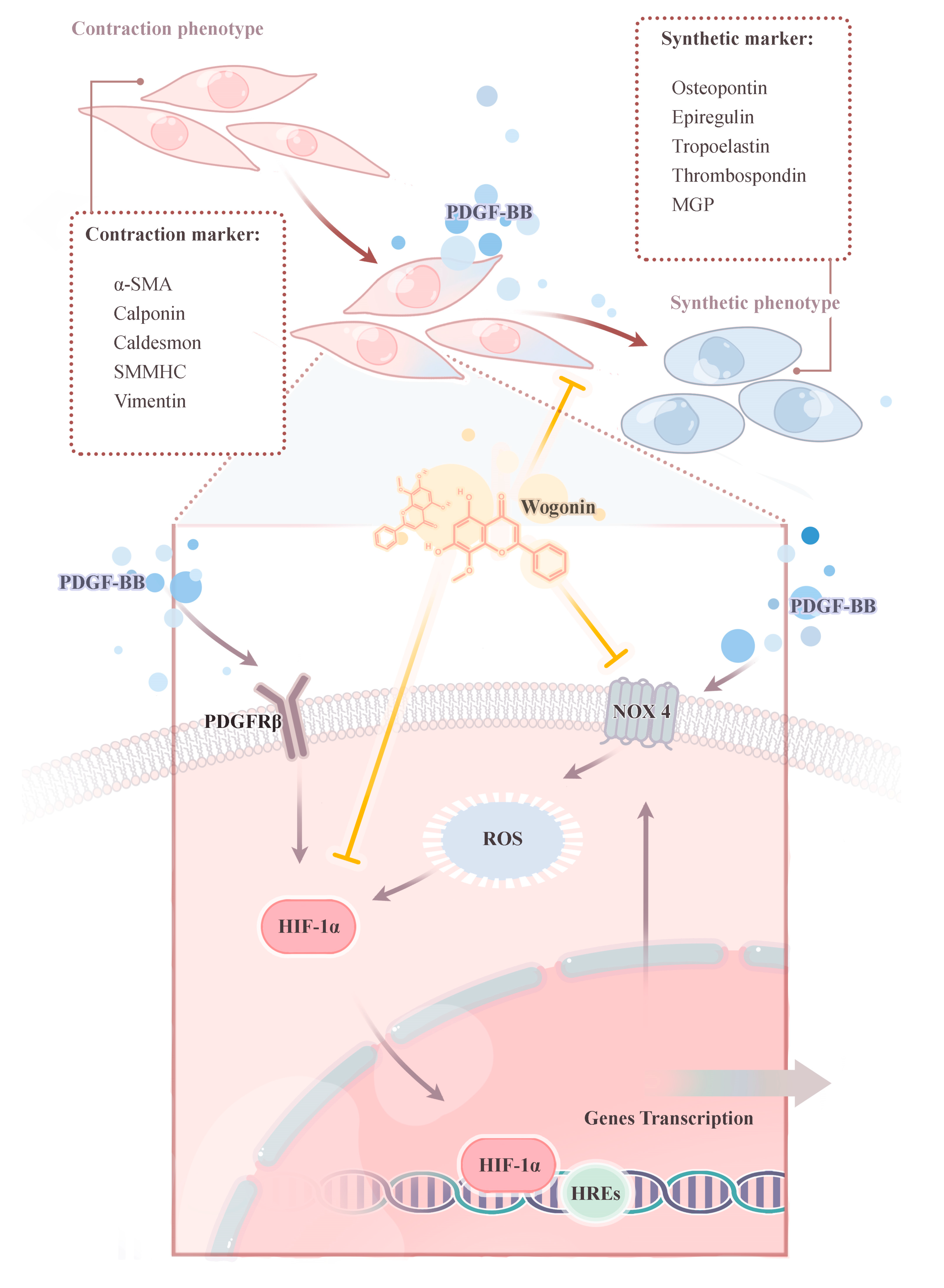
3.8. Effect of Wogonin on HIF-1/NOX4 Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lau, E.M.T.; Giannoulatou, E.; Celermajer, D.S.; Humbert, M. Epidemiology and treatment of pulmonary arterial hypertension. Nat. Rev. Cardiol. 2017, 14, 603–614. [Google Scholar] [CrossRef]
- Archer, S.L.; Weir, E.K.; Wilkins, M.R. Basic science of pulmonary arterial hypertension for clinicians: New concepts and experimental therapies. Circulation 2010, 121, 2045–2066. [Google Scholar] [CrossRef] [PubMed]
- Arora, T.K.; Arora, A.K.; Sachdeva, M.K.; Rajput, S.K.; Sharma, A.K. Pulmonary hypertension: Molecular aspects of current therapeutic intervention and future direction. J. Cell. Physiol. 2018, 233, 3794–3804. [Google Scholar] [CrossRef] [PubMed]
- Bello-Klein, A.; Mancardi, D.; da Rosa Araujo, A.S.; Schenkel, P.C.; de Lima Seolin, B.G.; Turck, P. Role of Redox Homeostasis and Inflammation in the Pathogenesis of Pulmonary Arterial Hypertension. Curr. Med. Chem. 2018, 25, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Freund-Michel, V.; Guibert, C.; Dubois, M.; Courtois, A.; Marthan, R.; Savineau, J.P.; Muller, B. Reactive oxygen species as therapeutic targets in pulmonary hypertension. Ther. Adv. Respir. Dis. 2013, 7, 175–200. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lu, W.; Yang, K.; Duan, X.; Li, M.; Chen, X.; Zhang, J.; Kuang, M.; Liu, S.; Wu, X.; et al. Tetramethylpyrazine: A promising drug for the treatment of pulmonary hypertension. Br. J. Pharmacol. 2020, 177, 2743–2764. [Google Scholar] [CrossRef]
- Maarman, G.J. Natural Antioxidants as Potential Therapy, and a Promising Role for Melatonin Against Pulmonary Hypertension. Adv. Exp. Med. Biol. 2017, 967, 161–178. [Google Scholar]
- Wang, J.; Lu, W.; Wang, W.; Zhang, N.; Wu, H.; Liu, C.; Chen, X.; Chen, Y.; Chen, Y.; Jiang, Q.; et al. Promising therapeutic effects of sodium tanshinone IIA sulfonate towards pulmonary arterial hypertension in patients. J. Thorac. Dis. 2013, 5, 169–172. [Google Scholar]
- Khan, S.; Kamal, M.A. Can Wogonin be Used in Controlling Diabetic Cardiomyopathy? Curr. Pharm. Des. 2019, 25, 2171–2177. [Google Scholar] [CrossRef]
- Qu, J.T.; Zhang, D.X.; Liu, F.; Mao, H.P.; Ma, Y.K.; Yang, Y.; Li, C.X.; Qiu, L.Z.; Geng, X.; Zhang, J.M.; et al. Vasodilatory Effect of Wogonin on the Rat Aorta and Its Mechanism Study. Biol. Pharm. Bull. 2015, 38, 1873–1878. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Zhang, B.; Chu, Z.; Han, B.; Zhang, X.; Huang, P.; Han, J. Wogonin Inhibits Cardiac Hypertrophy by Activating Nrf-2-Mediated Antioxidant Responses. Cardiovasc. Ther. 2021, 2021, 9995342. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Dong, L.; Liu, L.; Guo, J.; Zhao, L.; Zhang, J.; Bu, D.; Liu, X.; Huo, P.; Cao, W.; et al. HERB: A high-throughput experiment- and reference-guided database of traditional Chinese medicine. Nucleic Acids Res. 2021, 49, D1197–D1206. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.31–1.30.33. [Google Scholar] [CrossRef]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Zhao, L.F.; Wang, H.F.; Wen, Y.T.; Jiang, K.K.; Mao, X.M.; Zhou, Z.Y.; Yao, K.T.; Geng, Q.S.; Guo, D.; et al. GenCLiP 3: Mining human genes’ functions and regulatory networks from PubMed based on co-occurrences and natural language processing. Bioinformatics 2020, 36, 1973–1975. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Rani, U.P.; Kesavan, R.; Ganugula, R.; Avaneesh, T.; Kumar, U.P.; Reddy, G.B.; Dixit, M. Ellagic acid inhibits PDGF-BB-induced vascular smooth muscle cell proliferation and prevents atheroma formation in streptozotocin-induced diabetic rats. J. Nutr. Biochem. 2013, 24, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.X.; Jin, Y.; Wang, Z.; Li, M.Y.; Zhang, Z.H.; Wang, J.Y.; Xing, Y.; Ri, M.H.; Jin, C.H.; Xu, G.H.; et al. Curcumol inhibits the expression of programmed cell death-ligand 1 through crosstalk between hypoxia-inducible factor-1alpha and STAT3 (T705) signaling pathways in hepatic cancer. J. Ethnopharmacol. 2020, 257, 112835. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, J.K.; Law, S.M.; Brooks, C.L., 3rd. Flexible CDOCKER: Development and application of a pseudo-explicit structure-based docking method within CHARMM. J. Comput. Chem. 2016, 37, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Robertson, D.H.; Brooks, C.L., 3rd; Vieth, M. Detailed analysis of grid-based molecular docking: A case study of CDOCKER-A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef]
- Kunichika, N.; Landsberg, J.W.; Yu, Y.; Kunichika, H.; Thistlethwaite, P.A.; Rubin, L.J.; Yuan, J.X. Bosentan inhibits transient receptor potential channel expression in pulmonary vascular myocytes. Am. J. Respir. Crit. Care Med. 2004, 170, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Sysol, J.R.; Natarajan, V.; Machado, R.F. PDGF induces SphK1 expression via Egr-1 to promote pulmonary artery smooth muscle cell proliferation. Am. J. Physiol. Cell Physiol. 2016, 310, C983–C992. [Google Scholar] [CrossRef] [Green Version]
- Yuan, T.; Zhang, H.; Chen, Y.; Jiao, X.; Xie, P.; Fang, L.; Du, G. The Protective Effect of DL0805 Derivatives on Pulmonary Artery Cells and the Underlying Mechanisms Study. Curr. Vasc. Pharmacol. 2017, 15, 469–476. [Google Scholar] [CrossRef]
- Dietrich, D.R. Toxicological and pathological applications of proliferating cell nuclear antigen (PCNA), a novel endogenous marker for cell proliferation. Crit. Rev. Toxicol. 1993, 23, 77–109. [Google Scholar] [CrossRef]
- Yusuf-Makagiansar, H.; Anderson, M.E.; Yakovleva, T.V.; Murray, J.S.; Siahaan, T.J. Inhibition of LFA-1/ICAM-1 and VLA-4/VCAM-1 as a therapeutic approach to inflammation and autoimmune diseases. Med. Res. Rev. 2002, 22, 146–167. [Google Scholar]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.C.; Posch, A. The design of a quantitative western blot experiment. Biomed Res Int 2014, 2014, 361590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badran, A.; Nasser, S.A.; Mesmar, J.; El-Yazbi, A.F.; Bitto, A.; Fardoun, M.M.; Baydoun, E.; Eid, A.H. Reactive Oxygen Species: Modulators of Phenotypic Switch of Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2020, 21, 8764. [Google Scholar] [CrossRef]
- Zhang, M.J.; Zhou, Y.; Chen, L.; Wang, Y.Q.; Wang, X.; Pi, Y.; Gao, C.Y.; Li, J.C.; Zhang, L.L. An overview of potential molecular mechanisms involved in VSMC phenotypic modulation. Histochem. Cell Biol. 2016, 145, 119–130. [Google Scholar] [CrossRef]
- Maron, B.A.; Loscalzo, J. Pulmonary hypertension: Pathophysiology and signaling pathways. Handb. Exp. Pharmacol. 2013, 218, 31–58. [Google Scholar]
- Tajsic, T.; Morrell, N.W. Smooth muscle cell hypertrophy, proliferation, migration and apoptosis in pulmonary hypertension. Compr. Physiol. 2011, 1, 295–317. [Google Scholar]
- Wang, S.J.; Zhao, J.K.; Ren, S.; Sun, W.W.; Zhang, W.J.; Zhang, J.N. Wogonin affects proliferation and the energy metabolism of SGC-7901 and A549 cells. Exp. Ther. Med. 2019, 17, 911–918. [Google Scholar] [CrossRef] [Green Version]
- Xin, N.J.; Han, M.; Gao, C.; Fan, T.T.; Shi, W. Wogonin suppresses proliferation and invasion of skin epithelioid carcinoma cells through Notch1. Cell. Mol. Biol. 2020, 66, 29–33. [Google Scholar] [CrossRef]
- Wang, C.; Cui, C. Inhibition of Lung Cancer Proliferation by Wogonin is Associated with Activation of Apoptosis and Generation of Reactive Oxygen Species. Balkan. Med. J. 2019, 37, 29–33. [Google Scholar] [CrossRef]
- Li, M.; Zhu, H.; Hu, X.; Gao, F.; Hu, X.; Cui, Y.; Wei, X.; Xie, C.; Lv, G.; Zhao, Y.; et al. TMEM98, a novel secretory protein, promotes endothelial cell adhesion as well as vascular smooth muscle cell proliferation and migration. Can. J. Physiol. Pharmacol. 2021, 99, 536–548. [Google Scholar] [CrossRef]
- Xing, J.; Peng, K.; Cao, W.; Lian, X.; Wang, Q.; Wang, X. Effects of total flavonoids from Dracocephalum moldavica on the proliferation, migration, and adhesion molecule expression of rat vascular smooth muscle cells induced by TNF-α. Pharm. Biol. 2013, 51, 74–83. [Google Scholar] [CrossRef]
- Zhao, Y.; Yao, J.; Wu, X.P.; Zhao, L.; Zhou, Y.X.; Zhang, Y.; You, Q.D.; Guo, Q.L.; Lu, N. Wogonin suppresses human alveolar adenocarcinoma cell A549 migration in inflammatory microenvironment by modulating the IL-6/STAT3 signaling pathway. Mol. Carcinog. 2015, 54 (Suppl. S1), E81–E93. [Google Scholar] [CrossRef]
- Zhao, K.; Wei, L.; Hui, H.; Dai, Q.; You, Q.D.; Guo, Q.L.; Lu, N. Wogonin suppresses melanoma cell B16-F10 invasion and migration by inhibiting Ras-medicated pathways. PLoS ONE 2014, 9, e106458. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhou, Y.; Zhou, M.; Huang, Y.; Li, Z.; You, Q.; Lu, N.; Guo, Q. Wogonin influences vascular permeability via Wnt/β-catenin pathway. Mol. Carcinog. 2015, 54, 501–512. [Google Scholar] [CrossRef]
- Zhou, M.; Song, X.; Huang, Y.; Wei, L.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Wogonin inhibits H2O2-induced angiogenesis via suppressing PI3K/Akt/NF-κB signaling pathway. Vascul. Pharmacol. 2014, 60, 110–119. [Google Scholar] [CrossRef]
- Piao, H.Z.; Choi, I.Y.; Park, J.S.; Kim, H.S.; Cheong, J.H.; Son, K.H.; Jeon, S.J.; Ko, K.H.; Kim, W.K. Wogonin inhibits microglial cell migration via suppression of nuclear factor-kappa B activity. Int. Immunopharmacol. 2008, 8, 1658–1662. [Google Scholar] [CrossRef]
- Dunham-Snary, K.J.; Wu, D.; Sykes, E.A.; Thakrar, A.; Parlow, L.R.; Mewburn, J.D.; Parlow, J.L.; Archer, S.L. Hypoxic Pulmonary Vasoconstriction: From Molecular Mechanisms to Medicine. Chest 2017, 151, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, L.A.; Laurie, S.S. HIF and pulmonary vascular responses to hypoxia. J. Appl. Physiol. 2014, 116, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Roos, T.U.; Heiss, E.H.; Schwaiberger, A.V.; Schachner, D.; Sroka, I.M.; Oberan, T.; Vollmar, A.M.; Dirsch, V.M. Caffeic acid phenethyl ester inhibits PDGF-induced proliferation of vascular smooth muscle cells via activation of p38 MAPK, HIF-1α, and heme oxygenase-1. J. Nat. Prod. 2011, 74, 352–356. [Google Scholar] [CrossRef]
- Xiao, Y.; Peng, H.; Hong, C.; Chen, Z.; Deng, X.; Wang, A.; Yang, F.; Yang, L.; Chen, C.; Qin, X. PDGF Promotes the Warburg Effect in Pulmonary Arterial Smooth Muscle Cells via Activation of the PI3K/AKT/mTOR/HIF-1α Signaling Pathway. Cell. Physiol. Biochem. 2017, 42, 1603–1613. [Google Scholar] [CrossRef]
- Khalmuratova, R.; Lee, M.; Mo, J.H.; Jung, Y.; Park, J.W.; Shin, H.W. Wogonin attenuates nasal polyp formation by inducing eosinophil apoptosis through HIF-1α and survivin suppression. Sci. Rep. 2018, 8, 6201. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Yao, J.; Wang, F.; Zhou, M.; Zhou, Y.; Wang, H.; Wei, L.; Zhao, L.; Li, Z.; Lu, N.; et al. Wogonin inhibits tumor angiogenesis via degradation of HIF-1α protein. Toxicol. Appl. Pharmacol. 2013, 271, 144–155. [Google Scholar] [CrossRef]
- Diebold, I.; Petry, A.; Hess, J.; Gorlach, A. The NADPH oxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1. Mol. Biol. Cell 2010, 21, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Wang, H.R.; Hsieh, L.M. Antiproliferative effect of baicalein, a flavonoid from a Chinese herb, on vascular smooth muscle cell. Eur. J. Pharmacol. 1994, 251, 91–93. [Google Scholar] [CrossRef]
- Wu, X.; Zhi, F.; Lun, W.; Deng, Q.; Zhang, W. Baicalin inhibits PDGF-BB-induced hepatic stellate cell proliferation, apoptosis, invasion, migration and activation via the miR-3595/ACSL4 axis. Int. J. Mol. Med. 2018, 41, 1992–2002. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.H.; Wen, J.K.; Miao, S.B.; Jia, Z.; Hu, H.J.; Sun, R.H.; Wu, Y.; Han, M. Baicalin inhibits PDGF-BB-stimulated vascular smooth muscle cell proliferation through suppressing PDGFRbeta-ERK signaling and increase in p27 accumulation and prevents injury-induced neointimal hyperplasia. Cell Res. 2010, 20, 1252–1262. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Target | -CDCOKER ENERGY |
|---|---|---|
| Wogonin | PDGFR | 35.9576 |
| HIF-1α | 20.8013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Zeng, Z.; Wang, X.; Yuan, T.; Wang, C.; Liu, D.; Guo, J.; Chen, Y. Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation. Curr. Issues Mol. Biol. 2023, 45, 555-570. https://doi.org/10.3390/cimb45010037
Cui L, Zeng Z, Wang X, Yuan T, Wang C, Liu D, Guo J, Chen Y. Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation. Current Issues in Molecular Biology. 2023; 45(1):555-570. https://doi.org/10.3390/cimb45010037
Chicago/Turabian StyleCui, Lidan, Zuomei Zeng, Xinyue Wang, Tianyi Yuan, Can Wang, Dianlong Liu, Jian Guo, and Yucai Chen. 2023. "Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation" Current Issues in Molecular Biology 45, no. 1: 555-570. https://doi.org/10.3390/cimb45010037
APA StyleCui, L., Zeng, Z., Wang, X., Yuan, T., Wang, C., Liu, D., Guo, J., & Chen, Y. (2023). Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation. Current Issues in Molecular Biology, 45(1), 555-570. https://doi.org/10.3390/cimb45010037