
Lavender Essential Oil Modulates Hepatic Cholesterol Metabolism in HepG2 Cells

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
2.1. LEO Yield and Composition
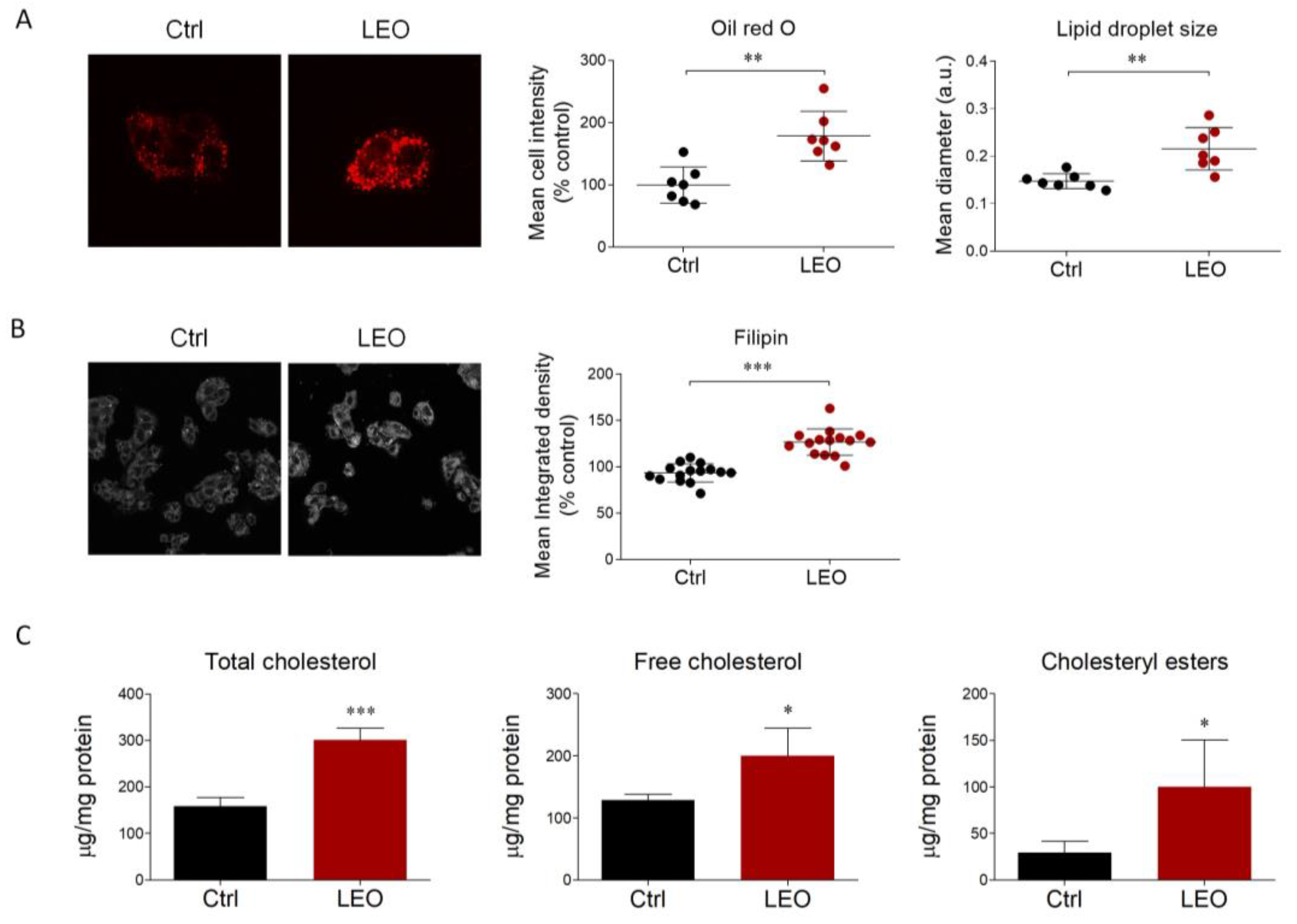
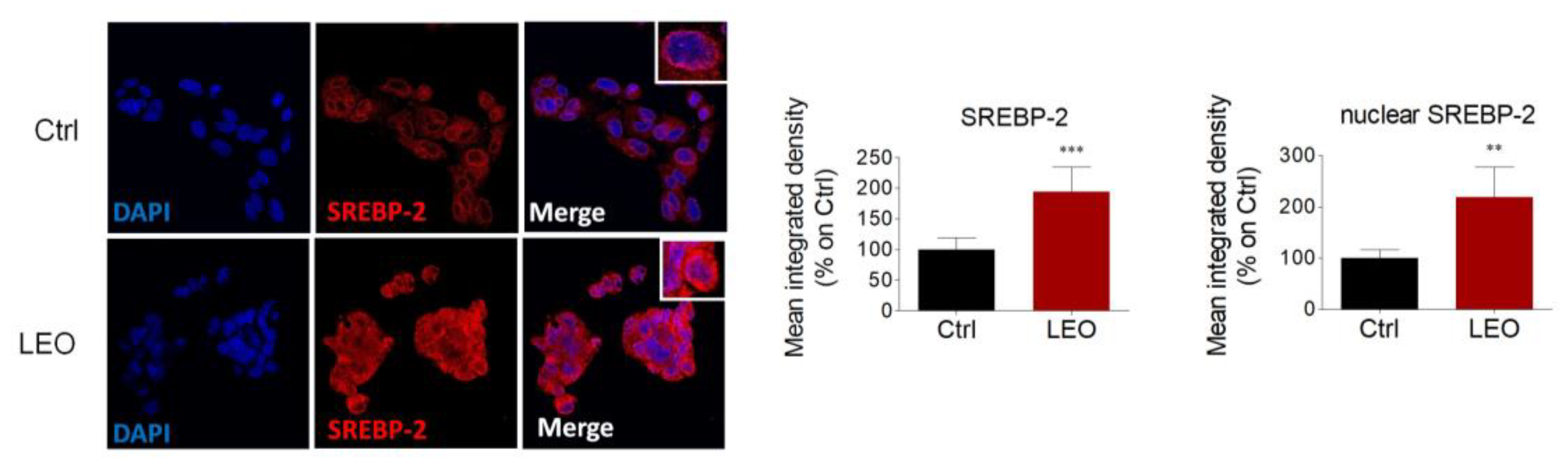
2.2. LEO Increases Intracellular Neutral Lipids and Cholesterol Content in HepG2 Cells
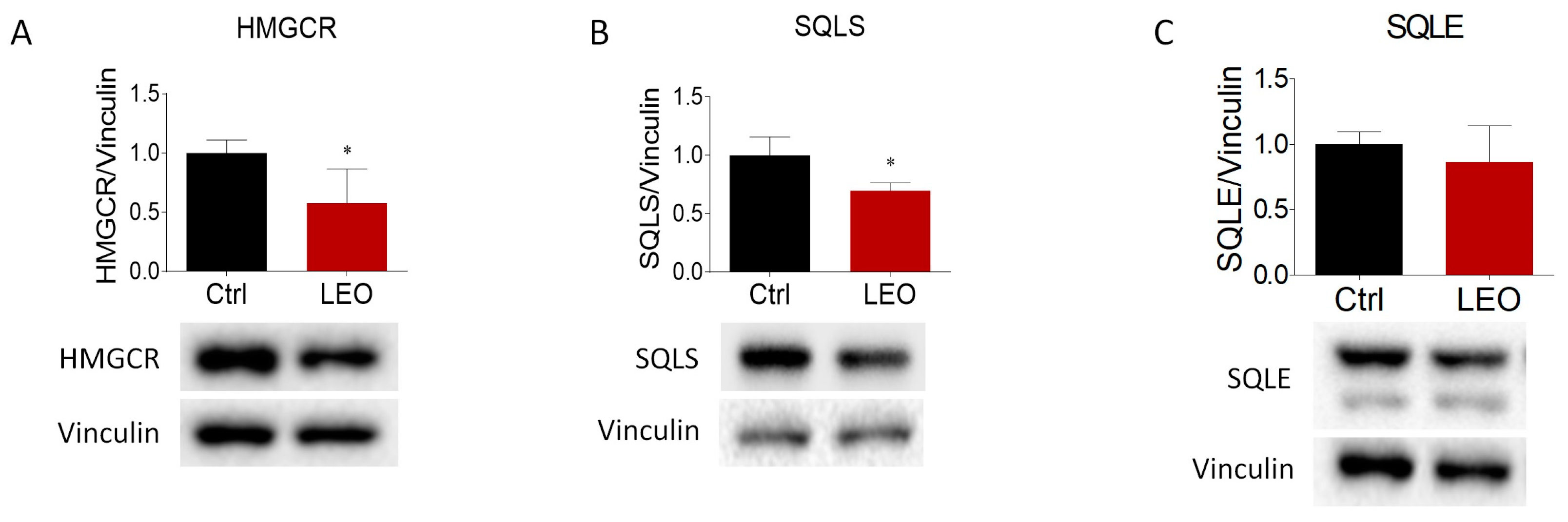
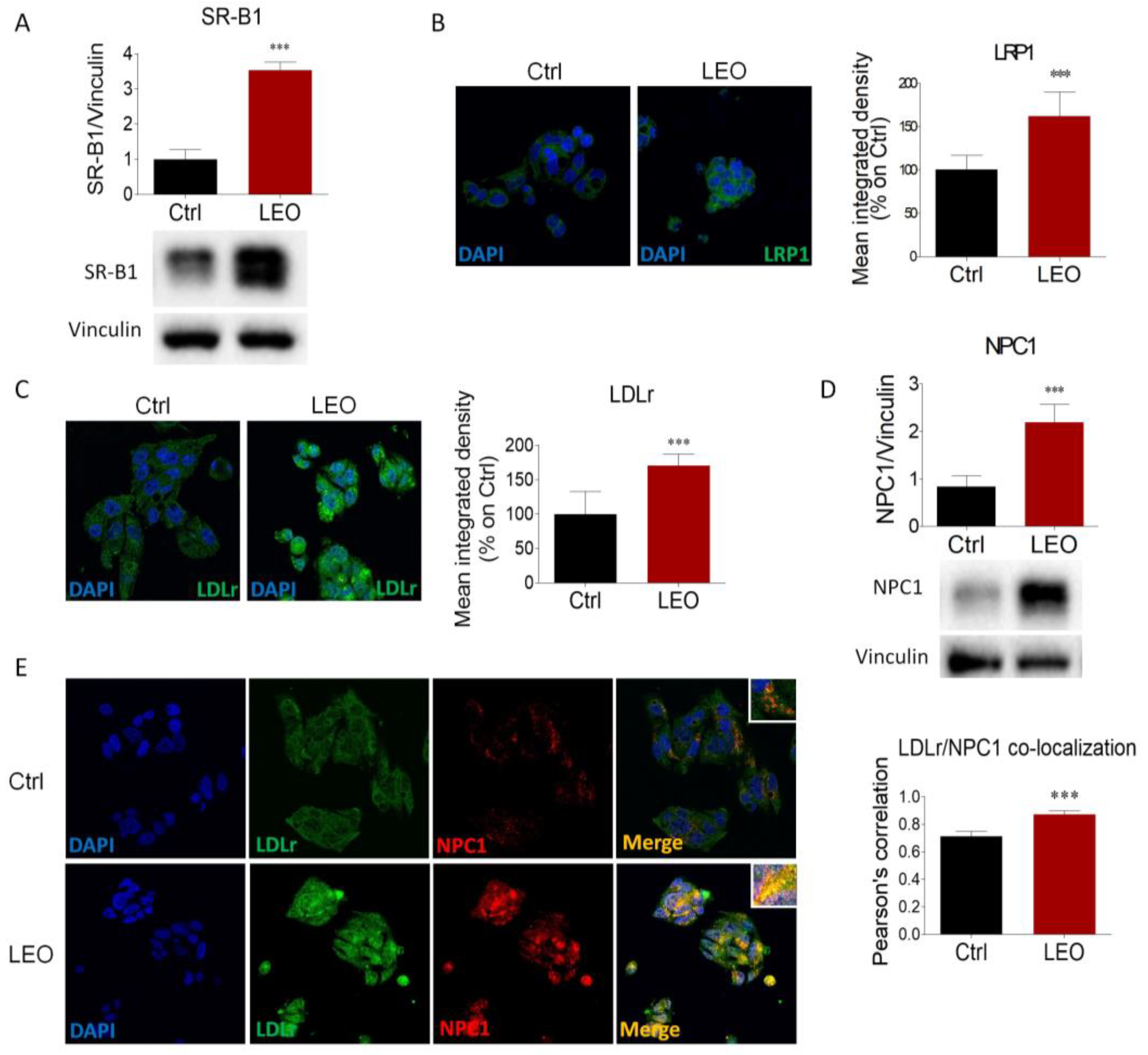
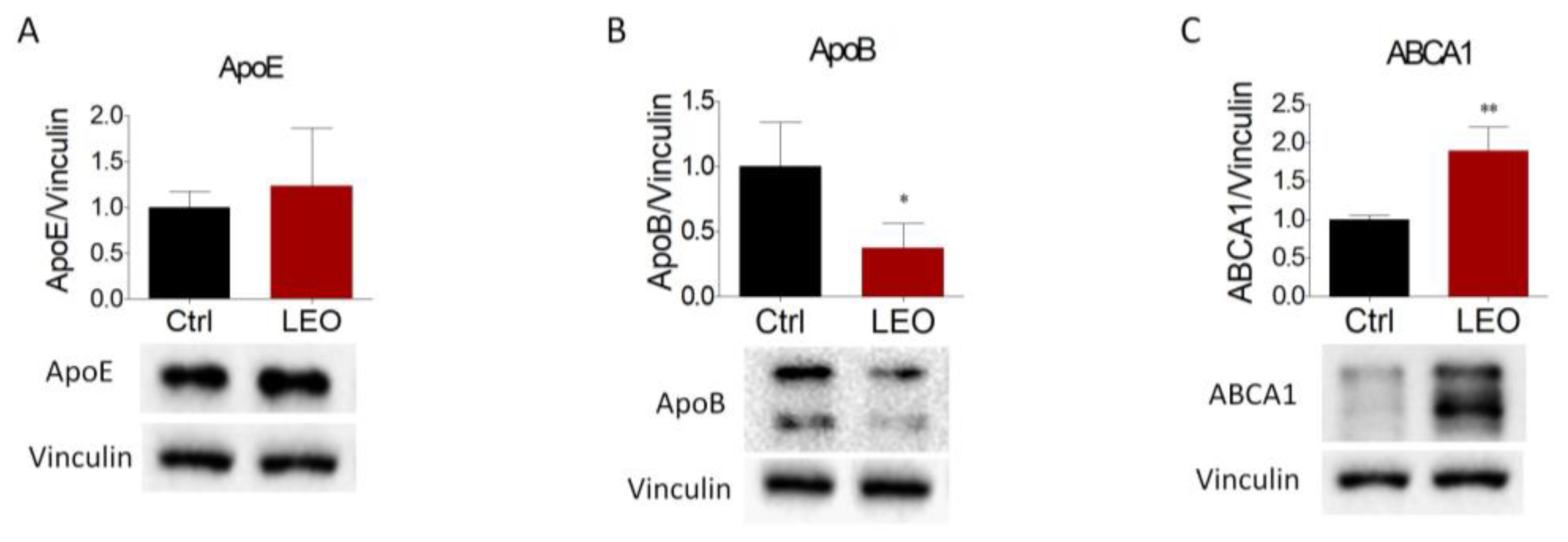
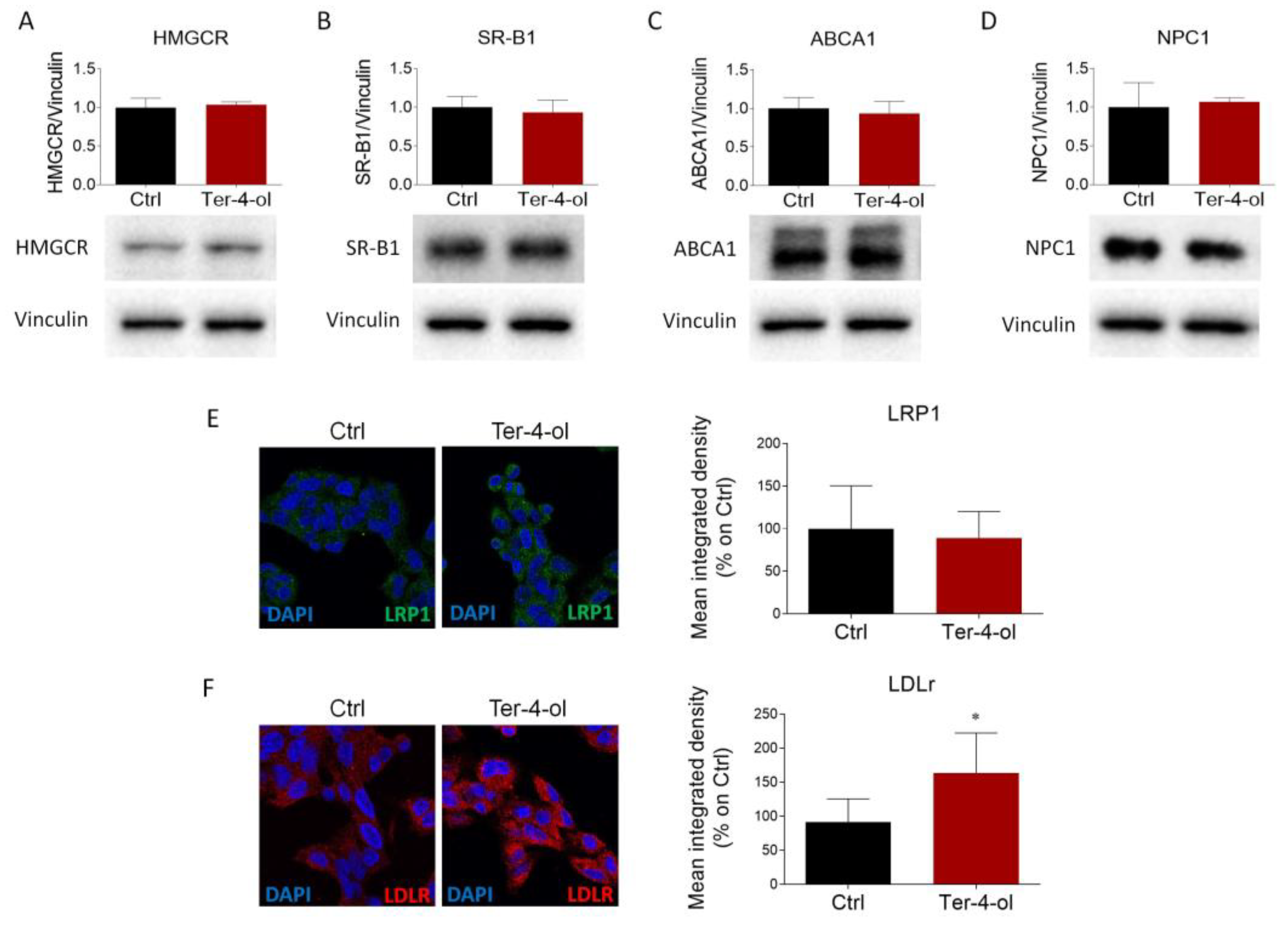
2.3. LEO Modulates the Expression of Proteins and Enzymes That Control Cholesterol Metabolism
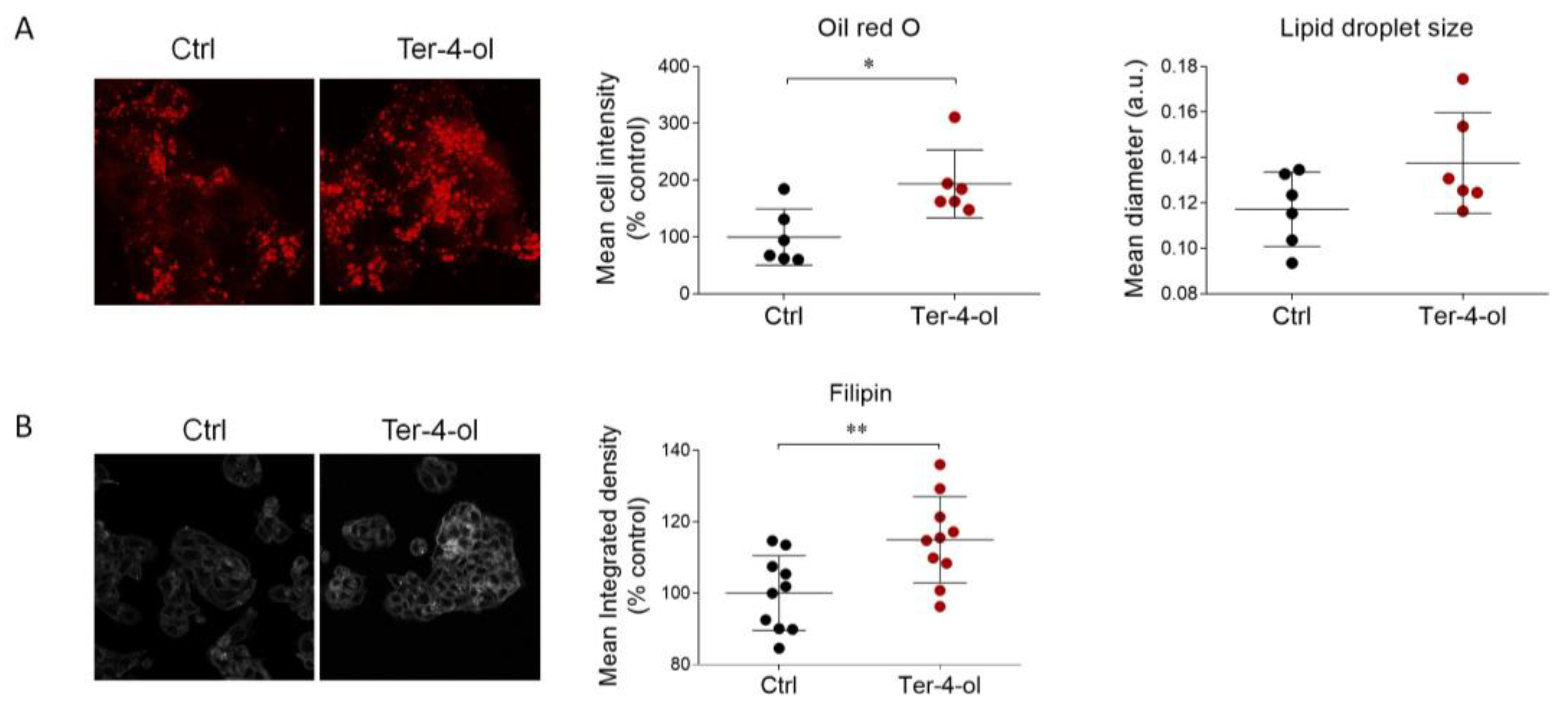
2.4. Terpinen-4-ol Partially Reproduces the Effects Induced by LEO on Cholesterol Metabolism
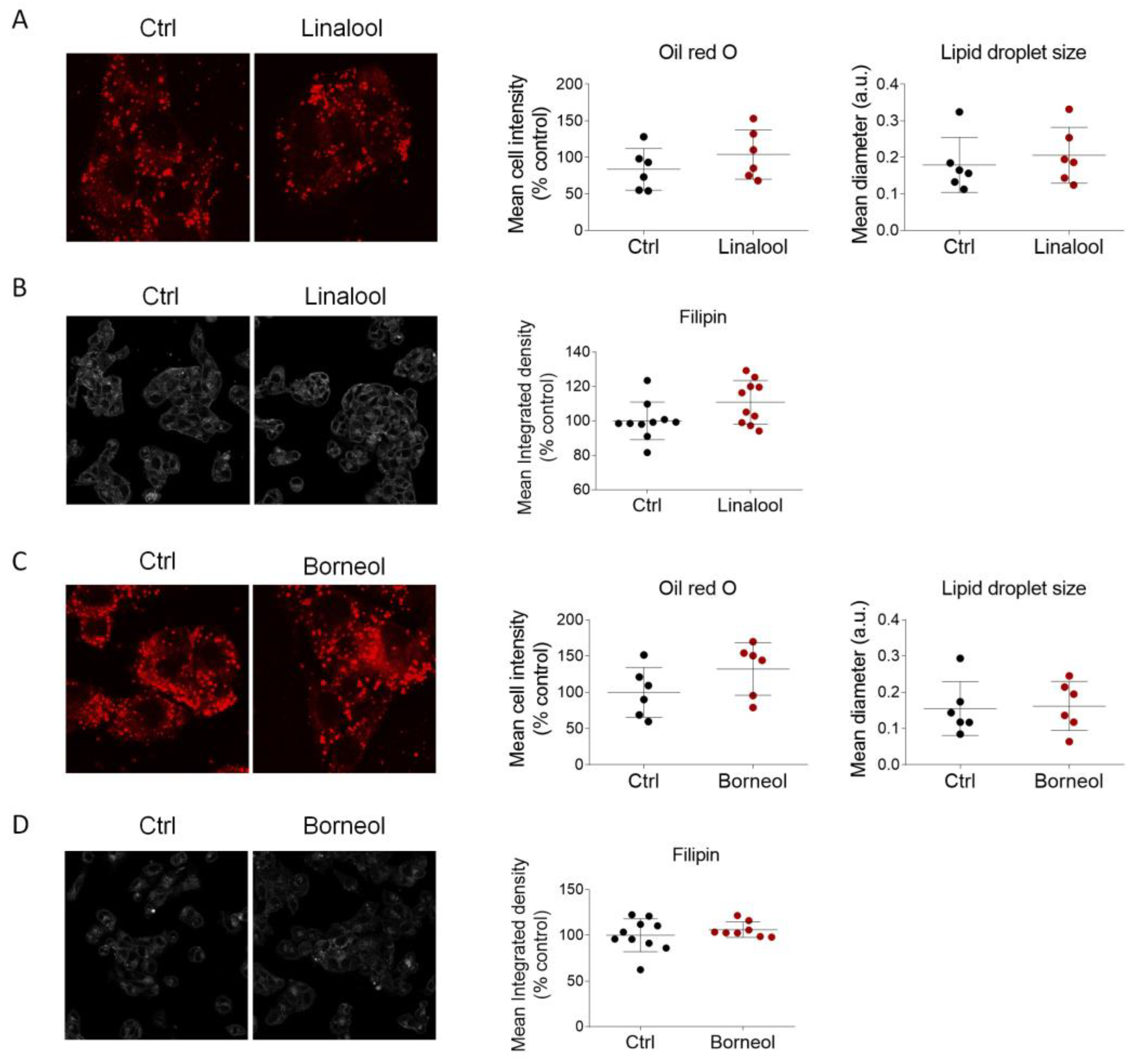
2.5. Linalool and Borneol Do Not Influence Cholesterol Metabolism in HepG2 Cells
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of the Essential Oil
4.3. GC-FID/GC-MS Analysis
4.4. Identification of Essential Oil Components
4.5. Cells Cultures
4.6. Oil Red O Staining and Quantification
4.7. Filipin Staining and Quantification
4.8. Cholesterol Quantification
4.9. Lysate Preparation and Western Blot Analysis
4.10. Immunofluorescence Staining
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colardo, M.; Martella, N.; Pensabene, D.; Siteni, S.; di Bartolomeo, S.; Pallottini, V.; Segatto, M. Neurotrophins as Key Regulators of Cell Metabolism: Implications for Cholesterol Homeostasis. Int. J. Mol. Sci. 2021, 22, 5692. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, J.R. Enzymes Involved in the Formation and Transformation of Steroid Hormones in the Fetal and Placental Compartments. J. Steroid. Biochem. Mol. Biol. 2005, 97, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Nowaczyk, M.J.M.; Irons, M.B. Smith-Lemli-Opitz Syndrome: Phenotype, Natural History, and Epidemiology. Am. J. Med. Genet. C Semin. Med. Genet. 2012, 160, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Pallottini, V.; Pfrieger, F.W. Understanding and Treating Niemann–Pick Type c Disease: Models Matter. Int. J. Mol. Sci. 2020, 21, 8979. [Google Scholar] [CrossRef]
- Trapani, L.; Segatto, M.; Pallottini, V. Regulation and Deregulation of Cholesterol Homeostasis: The Liver as a Metabolic “Power Station”. World J. Hepatol. 2012, 4, 184–190. [Google Scholar] [CrossRef]
- Silvente-Poirot, S.; Poirot, M. Cholesterol and Cancer, in the Balance. Science 2014, 343, 1445–1446. [Google Scholar] [CrossRef]
- Trapani, L.; Segatto, M.; Ascenzi, P.; Pallottini, V. Potential Role of Nonstatin Cholesterol Lowering Agents. IUBMB Life 2011, 63, 964–971. [Google Scholar] [CrossRef]
- Lecis C; Segatto M Cholesterol Homeostasis Imbalance and Brain Functioning: Neurological Disorders and Behavioral Consequences. J. Neurol. Neurol. Disord. 2014, 1, 101. [CrossRef]
- Brown, M.S.; Goldstein, J.L. The SREBP Pathway: Regulation Review of Cholesterol Metabolism by Proteolysis of a Membrane-Bound Transcription Factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Tansey, T.; Shechter, I. Squalene Synthase: Structure and Regulation. Prog. Nucleic Acid Res. Mol. Biol. 2001, 65, 157–195. [Google Scholar] [CrossRef]
- Brown, M.S.; Radhakrishnan, A.; Goldstein, J.L. Retrospective on Cholesterol Homeostasis: The Central Role of Scap. Annu. Rev. Biochem. 2018, 87, 783–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, G.A.; Yu, L.; Li, W.P.; Gerard, R.; Tuma, P.L.; Cohen, J.C.; Hobbs, H.H. ABCG5 and ABCG8 Are Obligate Heterodimers for Protein Trafficking and Biliary Cholesterol Excretion. J. Biol. Chem. 2003, 278, 48275–48282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Gouda, K.; Chisaki, I.; Ochiai, M.; Itagaki, S.; Iseki, K. Regulation Mechanism of ABCA1 Expression by Statins in Hepatocytes. Eur. J. Pharmacol. 2011, 662, 9–14. [Google Scholar] [CrossRef]
- Bahr, T.; Butler, G.; Rock, C.; Welburn, K.; Allred, K.; Rodriguez, D. Cholesterol-Lowering Activity of Natural Mono- and Sesquiterpenoid Compounds in Essential Oils: A Review and Investigation of Mechanisms Using in Silico Protein–Ligand Docking. Phytother. Res. 2021, 35, 4215–4245. [Google Scholar] [CrossRef]
- Rabiei, Z.; Rafieian-Kopaei, M.; Mokhtari, S.; Shahrani, M. Effect of Dietary Ethanolic Extract of Lavandula officinalis on Serum Lipids Profile in Rats. Iran. J. Pharm. Res. 2014, 13, 1295–1301. [Google Scholar]
- Cho, S.Y.; Jun, H.J.; Lee, J.H.; Jia, Y.; Kim, K.H.; Lee, S.J. Linalool Reduces the Expression of 3-Hydroxy-3-Methylglutaryl CoA Reductase via Sterol Regulatory Element Binding Protein-2- and Ubiquitin-Dependent Mechanisms. FEBS Lett. 2011, 585, 3289–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javitt, N.B. Hep G2 Cells as a Resource for Metabolic Studies: Lipoprotein, Cholesterol, and Bile Acids. FASEB J. 1990, 4, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Caprari, C.; Fantasma, F.; Divino, F.; Bucci, A.; Iorizzi, M.; Naclerio, G.; Ranalli, G.; Saviano, G. Chemical Profile, in Vitro Biological Activity and Comparison of Essential Oils from Fresh and Dried Flowers of Lavandula angustifolia L. Molecules 2021, 26, 5317. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/mass Spectrometry, 4th ed.; Allured Publishing Co.: Carol Stream, IL, USA, 2007; pp. 60188–62787. [Google Scholar]
- Mehlem, A.; Hagberg, C.E.; Muhl, L.; Eriksson, U.; Falkevall, A. Imaging of Neutral Lipids by Oil Red O for Analyzing the Metabolic Status in Health and Disease. Nat. Protoc. 2013, 8, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.-J.; Azhar, S.; Kraemer, F.B. SR-B1: A Unique Multifunctional Receptor for Cholesterol Influx and Efflux. Annu. Rev. Physiol. 2018, 80, 95–116. [Google Scholar] [CrossRef]
- Qin, Y.; Ting, F.; Kim, M.J.; Strelnikov, J.; Harmon, J.; Gao, F.; Dose, A.; Teng, B.B.; Alipour, M.A.; Yao, Z.; et al. Phosphatidylinositol-(4,5)-Bisphosphate Regulates Plasma Cholesterol through LDL (Low-Density Lipoprotein) Receptor Lysosomal Degradation. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.L.; Ginsberg, H.N. Hepatic Synthesis of Lipoproteins and Apolipoproteins. Semin. Liver Dis. 1992, 12, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Tonini, C.; Colardo, M.; Colella, B.; di Bartolomeo, S.; Berardinelli, F.; Caretti, G.; Pallottini, V.; Segatto, M. Inhibition of Bromodomain and Extraterminal Domain (BET) Proteins by JQ1 Unravels a Novel Epigenetic Modulation to Control Lipid Homeostasis. Int. J. Mol. Sci. 2020, 21, 1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 25. Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J. Terpinen-4-Ol, the Main Component of the Essential Oil of Melaleuca alternifolia (Tea Tree Oil), Suppresses Inflam.mmatory Mediator Production by Activated Human Monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Ma, X.; Jiang, M.; Cheng, Z.; Datsomor, O.; Zhao, G.; Zhan, K. The Role of Tea Tree Oil in Alleviating Palmitic Acid-Induced Lipid Accumulation in Bovine Hepatocytes. Front. Vet. Sci. 2022, 8, 814840. [Google Scholar] [CrossRef] [PubMed]
- Jun, H.J.; Lee, J.H.; Kim, J.; Jia, Y.; Kim, K.H.; Hwang, K.Y.; Yun, E.J.; Do, K.R.; Lee, S.J. Linalool Is a PPARα Ligand That Reduces Plasma TG Levels and Rewires the Hepatic Transcriptome and Plasma Metabolome. J. Lipid Res. 2014, 55, 1098–1110. [Google Scholar] [CrossRef] [Green Version]
- Madhuri, K.; Naik, P.R. Ameliorative effect of borneol, a natural bicyclic monoterpene against hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced diabetic Wistar rats. Biomed. Pharmacother. 2017, 96, 336–347. [Google Scholar] [CrossRef]
- Mooberry, L.K.; Sabnis, N.A.; Panchoo, M.; Nagarajan, B.; Lacko, A.G. Targeting the SR-B1 Receptor as a Gateway for Cancer Therapy and Imaging. Front. Pharmacol. 2016, 7, 466. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, H.B.; Yieh, L.; Osborne, T.F. Cooperation by Sterol Regulatory Element-Binding Protein and Sp1 in Sterol Regulation of Low Density Lipoprotein Receptor Gene. J. Biol. Chem. 1995, 270, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.H.; Kang, S.B.; Park, J.S.; Lee, B.W.; Kang, E.S.; Ahn, C.W.; Lee, H.C.; Cha, B.S. Up-Regulation of Hepatic Low-Density Lipoprotein Receptor-Related Protein 1: A Possible Novel Mechanism of Antiatherogenic Activity of Hydroxymethylglutaryl-Coenzyme A Reductase Inhibitor: Atorvastatin and Hepatic LRP1 Expression. Metabolism 2011, 60, 930–940. [Google Scholar] [CrossRef]
- DeBose-Boyd, R.A. Feedback Regulation of Cholesterol Synthesis: Sterol-Accelerated Ubiquitination and Degradation of HMG CoA Reductase. Cell Res. 2008, 18, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Nathan, J.A. Squalene and Cholesterol in the Balance at the ER Membrane. Proc. Natl. Acad. Sci. USA 2020, 117, 8228–8230. [Google Scholar] [CrossRef] [PubMed]
- Heidasch, R.; Avci, D.; Lüchtenborg, C.; Kale, D.; Beard, H.; Mentrup, T.; Barniol-Xicota, M.; Schröder, B.; Verhelst, S.; Brügger, B.; et al. Intramembrane Protease SPP Defines a Cholesterol-Regulated Switch of the Mevalonate Pathway. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wong, J.; Quinn, C.M.; Brown, A.J. SREBP-2 Positively Regulates Transcription of the Cholesterol Efflux Gene, ABCA1, by Generating Oxysterol Ligands for LXR. Biochem. J. 2006, 400, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.J.; Woo Park, K.; Heon Kim, K.; Kim, C.T.; Pill Baek, J.; Bang, K.H.; Choi, Y.M.; Lee, S.J. Asian Plantain (Plantago asiatica) Essential Oils Suppress 3-Hydroxy-3-Methylglutaryl-Co-Enzyme A Reductase Expression in Vitro and in Vivo and Show Hypocholesterolaemic Properties in Mice. Br. J. Nutr. 2008, 99, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council of Europe. European Pharmacopoeia, 5th ed.; Council of Europe: Strasbourg, France, 2004; Volume I, p. 217. [Google Scholar]
- 38. NIST/EPA/NIH; Mass Spectral Library. National Institute of Standard and Technology: Gaithersburg, MD, USA, 2005.
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Kovats, E. Gas Chromatographic Characterization of Organic Substances in the Retention Index System. In Advances in Chromatography; Giddings, J.C., Keller, R.A., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1965; Volume I, pp. 229–247. [Google Scholar]
- van den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Grob, R.L.; Kaiser, M.A. Qualitative and quantitative analysis by gas chromatography. In Modern Practice of Gas Chromatography; Grob, B., Ed.; John Wiley & Sons: New York, NY, USA, 2004; ISBN 0471229830. [Google Scholar]
- Göritz, C.; Thiebaut, R.; Tessier, L.-H.; Nieweg, K.; Moehle, C.; Buard, I.; Dupont, J.-L.; Schurgers, L.J.; Schmitz, G.; Pfrieger, F.W. Glia-induced neuronal differentiation by transcriptional regulation. Glia 2007, 55, 1108–1122. [Google Scholar] [CrossRef]
- Segatto, M.; Fittipaldi, R.; Pin, F.; Sartori, R.; Ko, K.D.; Zare, H.; Fenizia, C.; Zanchettin, G.; Pierobon, E.S.; Hatakeyama, S.; et al. Epigenetic targeting of bromodomain protein BRD4 counteracts cancer cachexia and prolongs survival. Nat. Commun. 2017, 8, 1707. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Exp. RI | Ref. RI | LEO Area % ± SD | Abbr. |
|---|---|---|---|---|
| 1,8-Cineole | 1034 | 1031 | 9.00 ± 0.29 | BMO |
| Linalool oxide cis | 1075 | 1072 | 1.68 ± 0.04 | AMO |
| Linalool oxide trans | 1089 | 1086 | 1.49 ± 0.02 | AMO |
| Linalool | 1105 | 1096 | 36.03 ± 0.10 | AMO |
| Camphor | 1148 | 1146 | 6.80 ± 0.07 | BMO |
| Borneol | 1170 | 1169 | 19.35 ± 0.12 | BMO |
| Terpinen-4-ol | 1182 | 1177 | 6.81 ± 0.02 | BMO |
| Exil butirate | 1196 | 1192 | 1.17 ± 0.02 | OT |
| Linalyl acetate | 1262 | 1257 | 2.75 ± 0.02 | AMO |
| β-Farnesene | 1462 | 1456 | 1.50 ± 0.01 | AS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martella, N.; Colardo, M.; Sergio, W.; Petraroia, M.; Varone, M.; Pensabene, D.; Russo, M.; Di Bartolomeo, S.; Ranalli, G.; Saviano, G.; et al. Lavender Essential Oil Modulates Hepatic Cholesterol Metabolism in HepG2 Cells. Curr. Issues Mol. Biol. 2023, 45, 364-378. https://doi.org/10.3390/cimb45010026
Martella N, Colardo M, Sergio W, Petraroia M, Varone M, Pensabene D, Russo M, Di Bartolomeo S, Ranalli G, Saviano G, et al. Lavender Essential Oil Modulates Hepatic Cholesterol Metabolism in HepG2 Cells. Current Issues in Molecular Biology. 2023; 45(1):364-378. https://doi.org/10.3390/cimb45010026
Chicago/Turabian StyleMartella, Noemi, Mayra Colardo, William Sergio, Michele Petraroia, Michela Varone, Daniele Pensabene, Miriam Russo, Sabrina Di Bartolomeo, Giancarlo Ranalli, Gabriella Saviano, and et al. 2023. "Lavender Essential Oil Modulates Hepatic Cholesterol Metabolism in HepG2 Cells" Current Issues in Molecular Biology 45, no. 1: 364-378. https://doi.org/10.3390/cimb45010026
APA StyleMartella, N., Colardo, M., Sergio, W., Petraroia, M., Varone, M., Pensabene, D., Russo, M., Di Bartolomeo, S., Ranalli, G., Saviano, G., & Segatto, M. (2023). Lavender Essential Oil Modulates Hepatic Cholesterol Metabolism in HepG2 Cells. Current Issues in Molecular Biology, 45(1), 364-378. https://doi.org/10.3390/cimb45010026