
Cooperative Binding of SRSF3 to Structured 3’ss-α Exon RNA during α Exon Inclusion in the ZO-1 mRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Protein Purification
2.2. Primers, Constructs, In Vitro Transcription, and RNase Probing
2.3. Transfections and In Vivo Splicing Assays
2.4. UV Crosslinking and EMSA
2.5. Western Blot
3. Results
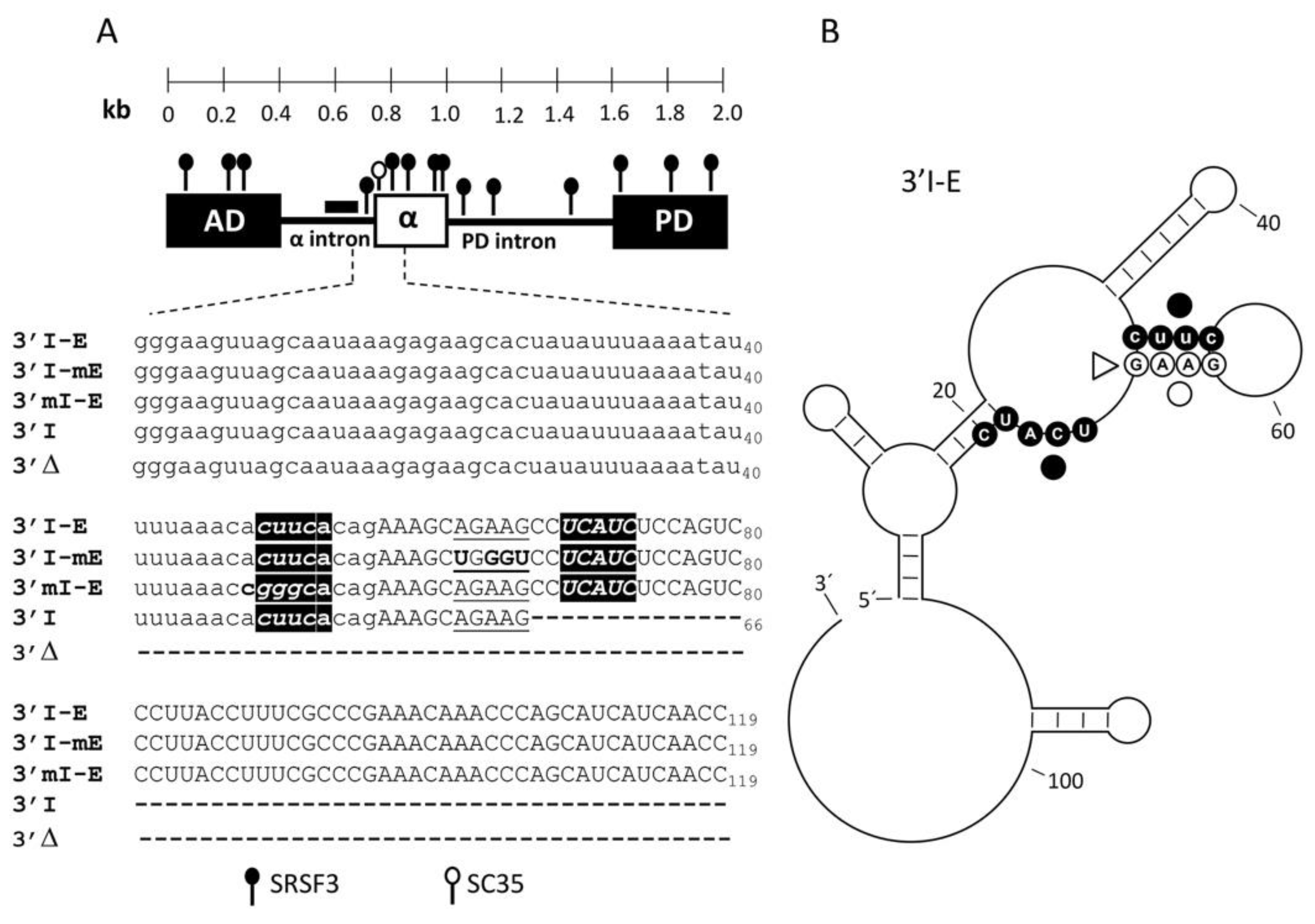
3.1. The Exon α Architecture
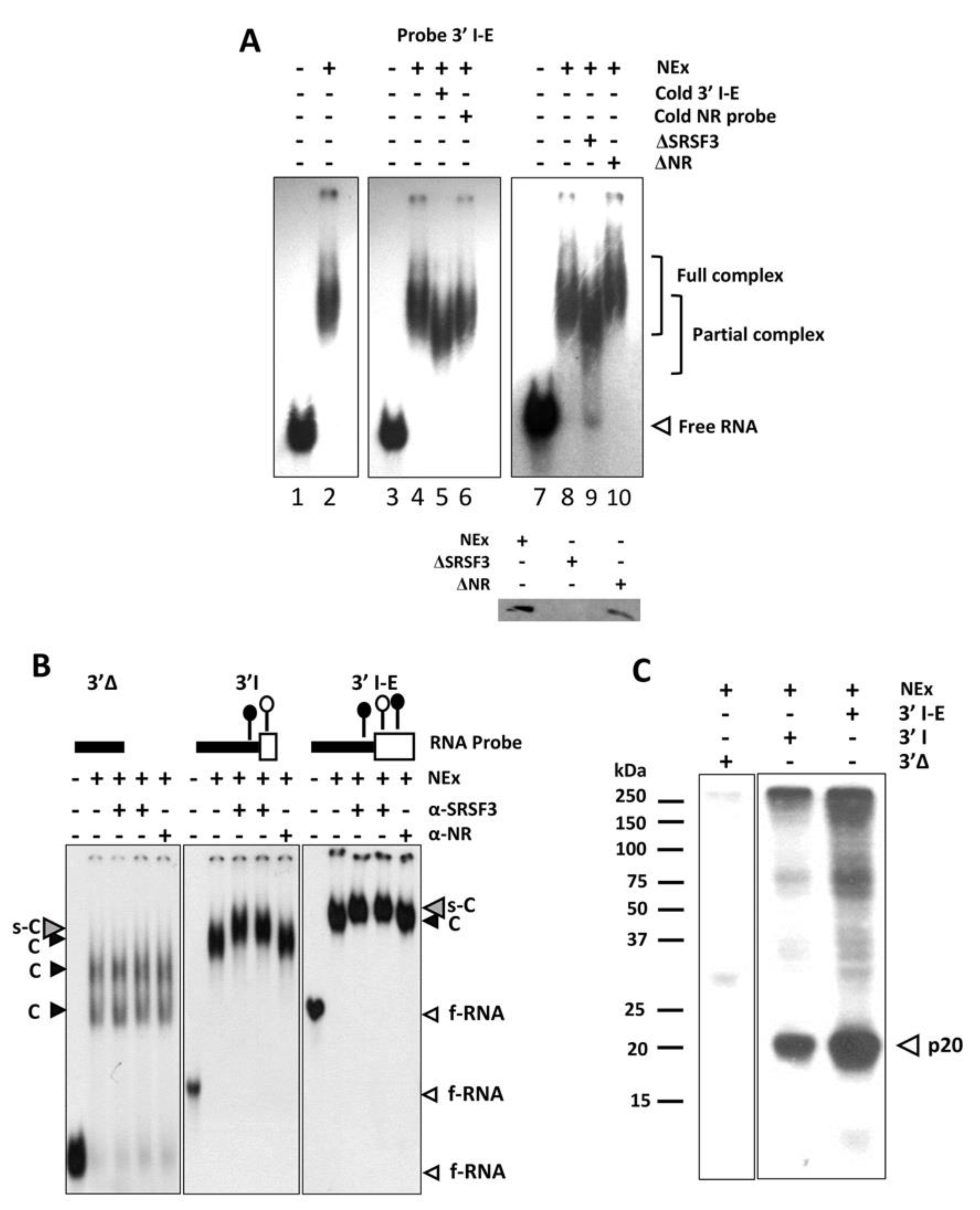
3.2. The 3’ss in Front of the α Exon Is Recognized by SRSF3 In Vitro
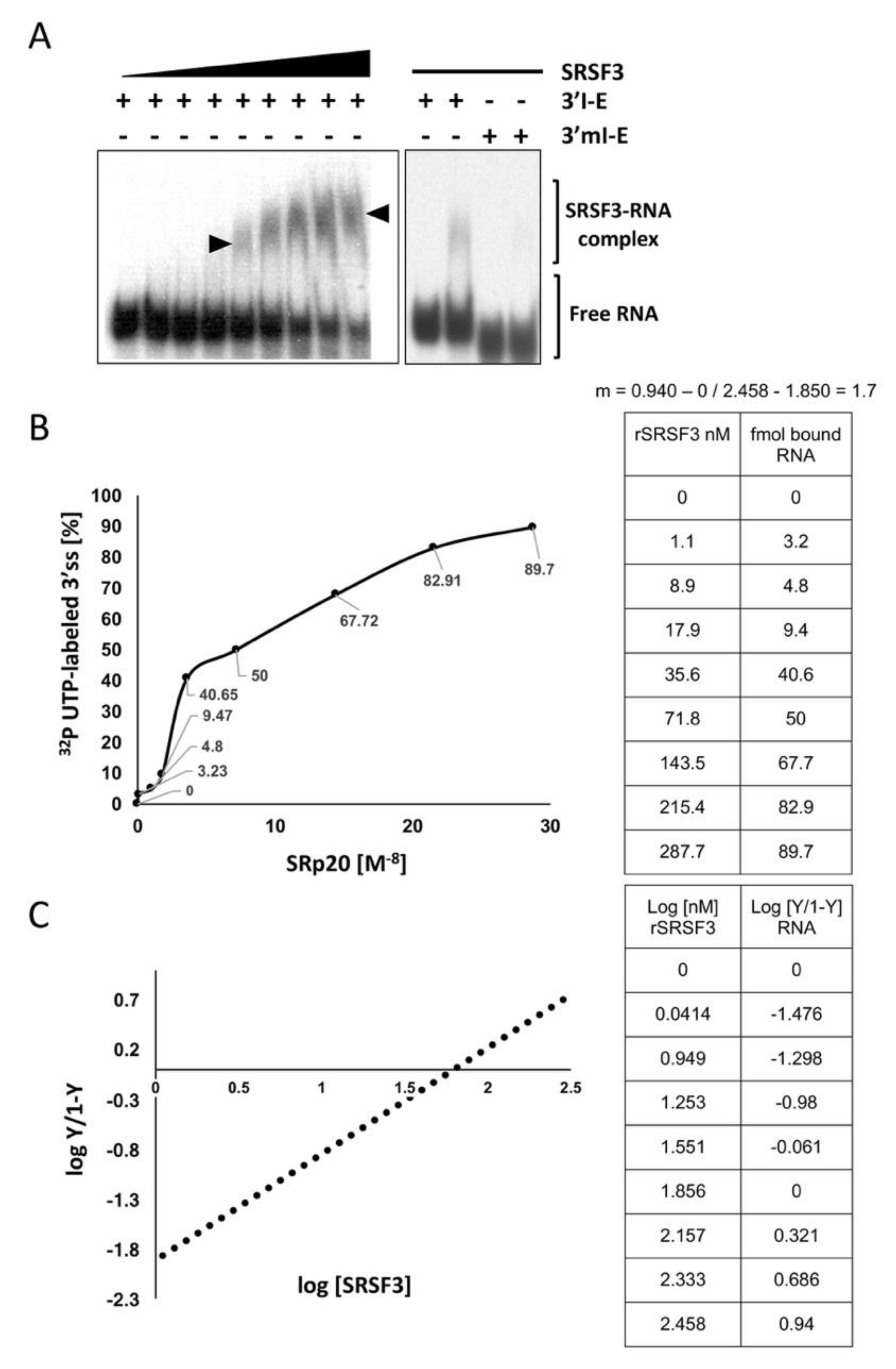
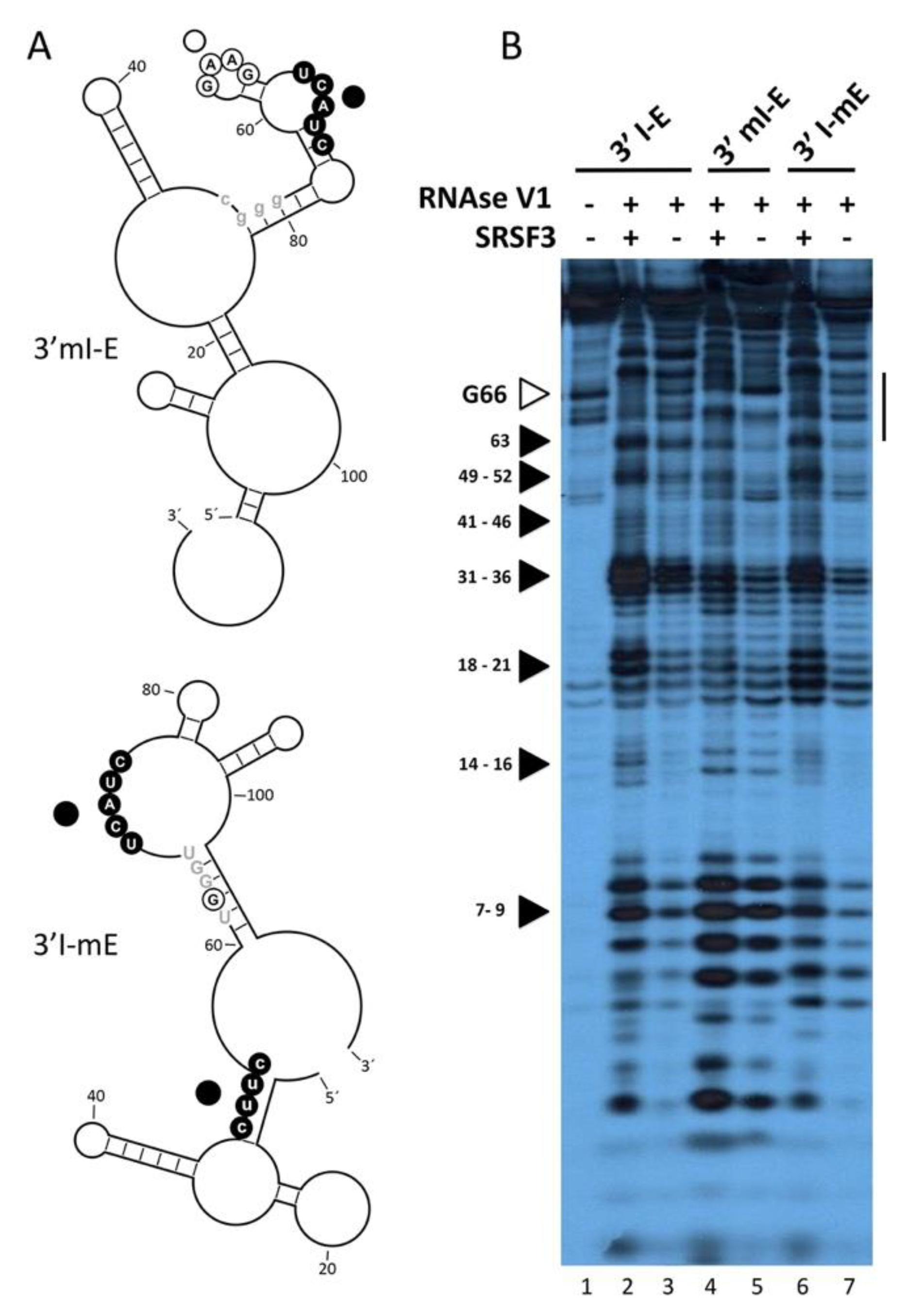
3.3. SRSF3 Cooperatively Binds around the 3’ss in a Structure-Dependent Way
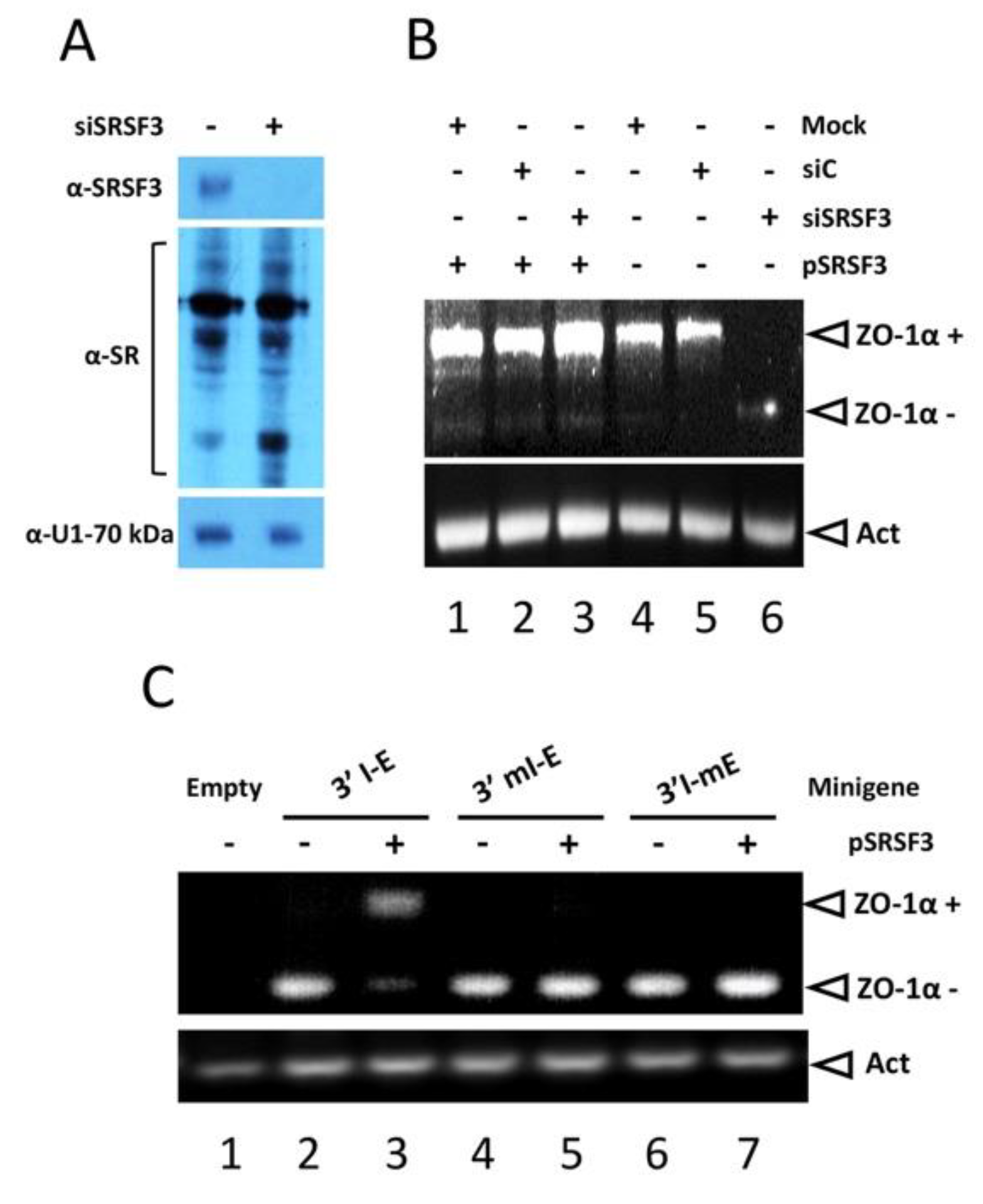
3.4. SRSF3 Regulates the Splicing of the α Exon In Vivo via Its Intronic Binding Site
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.; Hertel, K.J. Evolution of SR protein and hnRNP splicing regulatory factors. Wiley Interdiscip. Rev. RNA 2012, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shepard, P.J.; Hertel, K.J. The SR protein family. Genome Biol. 2009, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Corbo, C.; Orru, S.; Salvatore, F. SRp20: An overview of its role in human diseases. Biochem. Biophys. Res. Commun. 2013, 436, 1–5. [Google Scholar] [CrossRef]
- Galiana-Arnoux, D.; Lejeune, F.; Gesnel, M.C.; Stevenin, J.; Breathnach, R.; Del Gatto-Konczak, F. The CD44 alternative v9 exon contains a splicing enhancer responsive to the SR proteins 9G8, ASF/SF2, and SRp20. J. Biol. Chem. 2003, 278, 32943–32953. [Google Scholar] [CrossRef] [Green Version]
- de la Mata, M.; Kornblihtt, A.R. RNA polymerase II C-terminal domain mediates regulation of alternative splicing by SRp20. Nat. Struct. Mol. Biol. 2006, 13, 973–980. [Google Scholar] [CrossRef]
- Cavaloc, Y.; Bourgeois, C.F.; Kister, L.; Stevenin, J. The splicing factors 9G8 and SRp20 transactivate splicing through different and specific enhancers. RNA 1999, 5, 468–483. [Google Scholar] [CrossRef] [Green Version]
- Hargous, Y.; Hautbergue, G.M.; Tintaru, A.M.; Skrisovska, L.; Golovanov, A.P.; Stevenin, J.; Lian, L.Y.; Wilson, S.A.; Allain, F.H. Molecular basis of RNA recognition and TAP binding by the SR proteins SRp20 and 9G8. EMBO J. 2006, 25, 5126–5137. [Google Scholar] [CrossRef]
- Anko, M.L.; Muller-McNicoll, M.; Brandl, H.; Curk, T.; Gorup, C.; Henry, I.; Ule, J.; Neugebauer, K.M. The RNA-binding landscapes of two SR proteins reveal unique functions and binding to diverse RNA classes. Genome Biol. 2012, 13, R17. [Google Scholar] [CrossRef] [Green Version]
- Schaal, T.D.; Maniatis, T. Selection and characterization of pre-mRNA splicing enhancers: Identification of novel SR protein-specific enhancer sequences. Mol. Cell. Biol. 1999, 19, 1705–1719. [Google Scholar] [CrossRef] [Green Version]
- Schaal, T.D.; Maniatis, T. Multiple distinct splicing enhancers in the protein-coding sequences of a constitutively spliced pre-mRNA. Mol. Cell. Biol. 1999, 19, 261–273. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Arslan, A.D.; Pool, M.D.; Ho, T.T.; Darcy, K.M.; Coon, J.S.; Beck, W.T. Knockdown of splicing factor SRp20 causes apoptosis in ovarian cancer cells and its expression is associated with malignancy of epithelial ovarian cancer. Oncogene 2011, 30, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.; Garner, B.; Halliday, G.M.; Kwok, J.B. Srp20 regulates TrkB pre-mRNA splicing to generate TrkB-Shc transcripts with implications for Alzheimer’s disease. J. Neurochem. 2012, 123, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Willott, E.; Balda, M.S.; Heintzelman, M.; Jameson, B.; Anderson, J.M. Localization and differential expression of two isoforms of the tight junction protein ZO-1. Am. J. Physiol. 1992, 262, C1119–C1124. [Google Scholar] [CrossRef]
- Martinez-Contreras, R.; Galindo, J.M.; Aguilar-Rojas, A.; Valdes, J. Two exonic elements in the flanking constitutive exons control the alternative splicing of the alpha exon of the ZO-1 pre-mRNA. Biochim. Biophys. Acta 2003, 1630, 71–83. [Google Scholar] [CrossRef]
- Dignam, J.D. Preparation of extracts from higher eukaryotes. Methods Enzymol. 1990, 182, 194–203. [Google Scholar] [CrossRef]
- Zahler, A.M.; Lane, W.S.; Stolk, J.A.; Roth, M.B. SR proteins: A conserved family of pre-mRNA splicing factors. Genes Dev. 1992, 6, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Urrutia, E.; Valdes, J.; Bonilla-Moreno, R.; Martinez-Salazar, M.; Martinez-Garcia, M.; Berumen, J.; Villegas-Sepulveda, N. A few nucleotide polymorphisms are sufficient to recruit nuclear factors differentially to the intron 1 of HPV-16 intratypic variants. Virus Res. 2012, 166, 43–53. [Google Scholar] [CrossRef]
- Black, D.L.; Chan, R.; Min, H.; Wang, J.; Bell, L. The electrophoretic mobility shift assay for RNA binding proteins. In RNA: Protein Interactions (A Practical Approach); The Prtactical Approach Series; Smith, C.W.J., Hames, B.D., Eds.; Oxford University Press: New York, NY, USA, 1998. [Google Scholar]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jumaa, H.; Guenet, J.L.; Nielsen, P.J. Regulated expression and RNA processing of transcripts from the Srp20 splicing factor gene during the cell cycle. Mol. Cell. Biol. 1997, 17, 3116–3124. [Google Scholar] [CrossRef] [Green Version]
- Jumaa, H.; Nielsen, P.J. The splicing factor SRp20 modifies splicing of its own mRNA and ASF/SF2 antagonizes this regulation. EMBO J. 1997, 16, 5077–5085. [Google Scholar] [CrossRef]
- Shukla, S.; Dirksen, W.P.; Joyce, K.M.; Le Guiner-Blanvillain, C.; Breathnach, R.; Fisher, S.A. TIA proteins are necessary but not sufficient for the tissue-specific splicing of the myosin phosphatase targeting subunit 1. J. Biol. Chem. 2004, 279, 13668–13676. [Google Scholar] [CrossRef] [Green Version]
- ten Dam, G.B.; Wieringa, B.; Poels, L.G. Alternative splicing of CD45 pre-mRNA is uniquely obedient to conditions in lymphoid cells. Biochim. Biophys. Acta 1999, 1446, 317–333. [Google Scholar] [CrossRef]
- Huang, Y.; Steitz, J.A. Splicing factors SRp20 and 9G8 promote the nucleocytoplasmic export of mRNA. Mol. Cell. 2001, 7, 899–905. [Google Scholar] [CrossRef]
- Sen, S.; Talukdar, I.; Webster, N.J. SRp20 and CUG-BP1 modulate insulin receptor exon 11 alternative splicing. Mol. Cell. Biol. 2009, 29, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Chebli, K.; Gattoni, R.; Schmitt, P.; Hildwein, G.; Stevenin, J. The 216-nucleotide intron of the E1A pre-mRNA contains a hairpin structure that permits utilization of unusually distant branch acceptors. Mol. Cell. Biol. 1989, 9, 4852–4861. [Google Scholar] [CrossRef] [Green Version]
- Clouet d’Orval, B.; d’Aubenton Carafa, Y.; Sirand-Pugnet, P.; Gallego, M.; Brody, E.; Marie, J. RNA secondary structure repression of a muscle-specific exon in HeLa cell nuclear extracts. Science 1991, 252, 1823–1828. [Google Scholar] [CrossRef]
- Estes, P.A.; Cooke, N.E.; Liebhaber, S.A. A native RNA secondary structure controls alternative splice-site selection and generates two human growth hormone isoforms. J. Biol. Chem. 1992, 267, 14902–14908. [Google Scholar] [CrossRef]
- Jacquenet, S.; Ropers, D.; Bilodeau, P.S.; Damier, L.; Mougin, A.; Stoltzfus, C.M.; Branlant, C. Conserved stem-loop structures in the HIV-1 RNA region containing the A3 3′ splice site and its cis-regulatory element: Possible involvement in RNA splicing. Nucleic Acids Res. 2001, 29, 464–478. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Hoffman, B.E.; Lis, J.T. A specific RNA hairpin loop structure binds the RNA recognition motifs of the Drosophila SR protein B52. Mol. Cell. Biol. 1997, 17, 2649–2657. [Google Scholar] [CrossRef] [Green Version]
- Damgaard, C.K.; Tange, T.O.; Kjems, J. hnRNP A1 controls HIV-1 mRNA splicing through cooperative binding to intron and exon splicing silencers in the context of a conserved secondary structure. RNA 2002, 8, 1401–1415. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Green, M.R. A pathway of sequential arginine-serine-rich domain-splicing signal interactions during mammalian spliceosome assembly. Mol. Cell 2004, 16, 363–373. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Luis, T.A.; Ortuño-Pineda, C.; Galindo-Rosales, J.M.; Saucedo-Cárdenas, O.; Valdés, J. Cooperative Binding of SRSF3 to Structured 3’ss-α Exon RNA during α Exon Inclusion in the ZO-1 mRNA. Curr. Issues Mol. Biol. 2023, 45, 593-603. https://doi.org/10.3390/cimb45010039
Ruiz-Luis TA, Ortuño-Pineda C, Galindo-Rosales JM, Saucedo-Cárdenas O, Valdés J. Cooperative Binding of SRSF3 to Structured 3’ss-α Exon RNA during α Exon Inclusion in the ZO-1 mRNA. Current Issues in Molecular Biology. 2023; 45(1):593-603. https://doi.org/10.3390/cimb45010039
Chicago/Turabian StyleRuiz-Luis, Tea Anastasia, Carlos Ortuño-Pineda, José Manuel Galindo-Rosales, Odila Saucedo-Cárdenas, and Jesús Valdés. 2023. "Cooperative Binding of SRSF3 to Structured 3’ss-α Exon RNA during α Exon Inclusion in the ZO-1 mRNA" Current Issues in Molecular Biology 45, no. 1: 593-603. https://doi.org/10.3390/cimb45010039
APA StyleRuiz-Luis, T. A., Ortuño-Pineda, C., Galindo-Rosales, J. M., Saucedo-Cárdenas, O., & Valdés, J. (2023). Cooperative Binding of SRSF3 to Structured 3’ss-α Exon RNA during α Exon Inclusion in the ZO-1 mRNA. Current Issues in Molecular Biology, 45(1), 593-603. https://doi.org/10.3390/cimb45010039