
The Sphingolipid-Signaling Pathway as a Modulator of Infection by SARS-CoV-2




{kind=link}
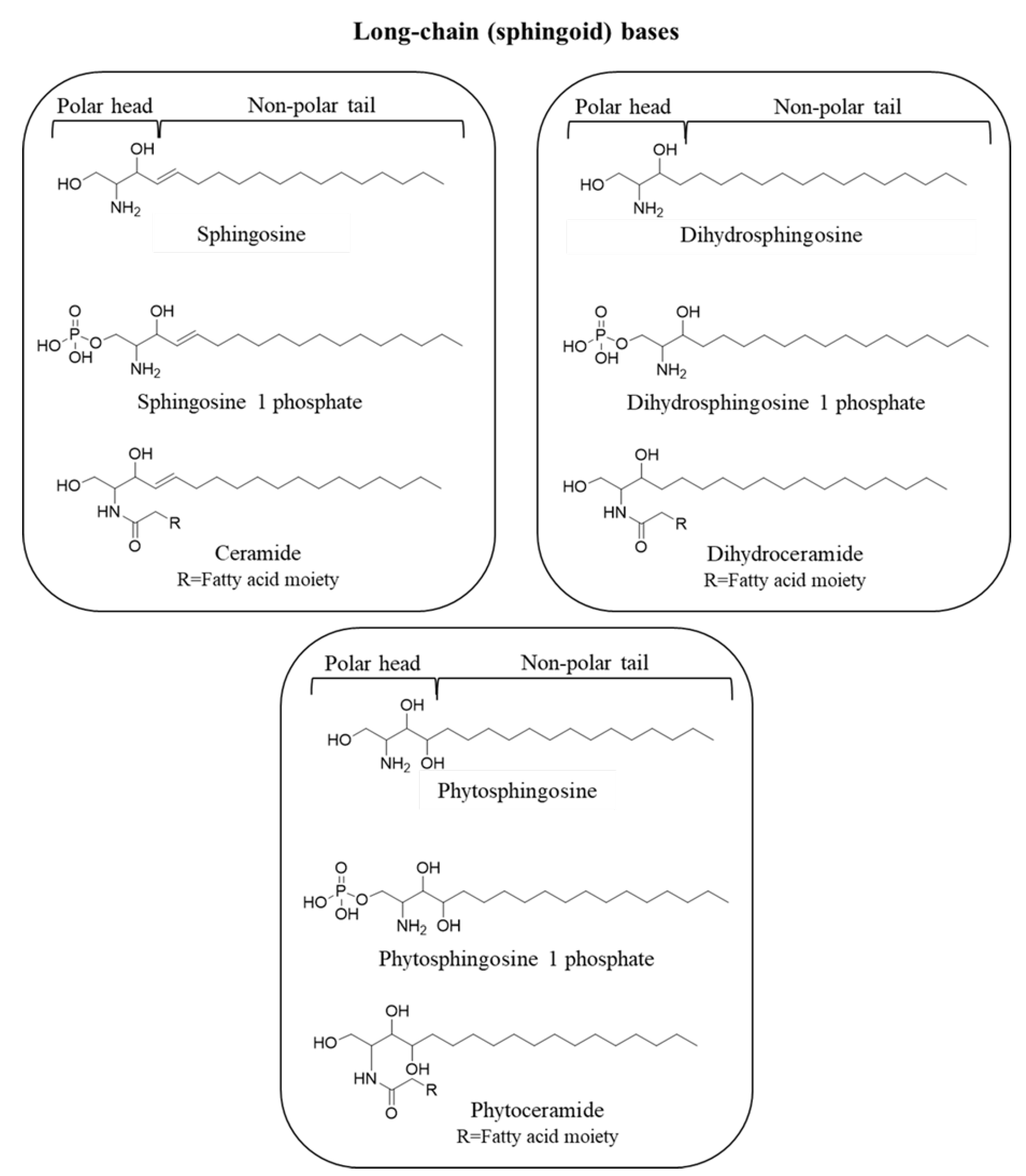{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. SARS-CoV-2 Life Cycle
2.1. Virus Binding and Entry
2.2. Viral Replication
2.3. Viral Release
3. Sphingolipids: Metabolism and Function
3.1. De Novo Pathway and Ceramide Transport to the Golgi
3.2. Sphingomyelinase Pathway
3.3. Salvage/Recycling Pathway
4. Sphingolipids in SARS-CoV-2 Life Cycle
4.1. Viral Entry
4.2. Viral Replication
4.3. Viral Release
5. Role of Sphingolipids in COVID-19 Severity and Complications
6. Sphingolipids as Potential Therapeutic Targets in SARS-CoV-2 Infection
- GT-11: this is a cyclopropene Cer molecule, where the C4–C5 double bond of Cer is replaced by a cyclopropene unit. GT-11 and its analogues act as DES1 inhibitors and have been studied mainly in cancer models, where they increase autophagy and apoptosis and inhibit tumor growth in mice models [101,102,103]. Interestingly, GT-11 is also able to inhibit flavivirus infection (e.g., West Nile virus) in a dose-dependent manner through DES inhibition [104].
- XM462: this molecule is a dihydroceramide analogue, where the C5 methylene group of the sphinganine moiety is replaced by a sulfur atom. XM462 and its analogues may inhibit both DES and ASAH1, although with different efficacy according to the characteristics of the N-acyl group [105]. Similar to another DES inhibitor (XM461), this compound can modulate autophagy and reduce amyloid secretion in primary neurons of a transgenic Alzheimer’s Disease model [106].
- SKI II: Sphingosine kinase inhibitor II, which interferes with SphK1 and SphK2, as well as DES1 [107], has been tested in the field of viral diseases, where it has been shown to reduce the virus replication of measles in both epithelial and lymphoid cell lines and affect mTOR Complex 1 (mTORC1) downstream signaling, likely through multiple mechanisms (e.g., reduction of rpS6 protein expression as well as its phosphorylation) [108,109];
- PF-543: this is a novel potent and selective inhibitor of SphK1 that has been studied in breast and colon cancers (IC50 = 2 nM; Ki = 3.6 nM: >100-fold selectivity for Sphk1 over Sphk2) [110].
7. Conclusions
- different acyl chain lengths not only characterize Cers structurally but also influence their physiological effects, which may be beneficial or detrimental depending on their molecular structure;
- Cers characterized by the same acyl chain length may play different physiological functions and pathophysiological roles according to their localization in the cellular microenvironment;
- Cers can interact with other drugs, as shown in the cancer field, and these interactions need to be carefully taken into account [114].
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| COVID-19 | Coronavirus disease 2019 |
| COVID-ARDS | COVID-19—associated acute respiratory distress syndrome |
| Cer | Ceramides |
| SL | Sphingolipid |
| ASM | Acid sphingomyelinase |
| S1P | Sphingosine-1-phosphate |
| S1PR1 | Sphingosine-1-phosphate receptor 1 |
| INF-α | Interferon alpha |
| ORF1a; ORF1b | Open reading frames 1a and 1b |
| S | Spike protein |
| E | Envelope protein |
| M | Membrane protein |
| N | Nucleocapsid protein |
| ACE2 | Angiotensin-converting enzyme 2 |
| RDB | receptor-binding domain |
| NSPs | non-structural proteins |
| RTC | replication-transcription complex |
| RdRp | RNA-dependent RNA polymerase |
| ER | endoplasmic reticulum |
| ERGIC | ER-to-Golgi intermediate compartment |
| Sph | sphingosine |
| CerS | ceramide synthase |
| DHCer | dihydroceramide |
| GLS | glycosphingolipids |
| SM | sphingomyelins |
| SPT | serine palmitoyltransferase |
| DES1 | dihydroceramide desaturase-1 |
| CERT | Cer transport protein |
| SMS | sphingomyelin synthase |
| DAG | diacylglycerol |
| CerK | Cer kinase |
| C1P | Cer-1-phosphate |
| SMases | sphingomyelinases |
| CDases | ceramidases |
| ASAH1 | acid CDase |
| ASAH2 | neutral CDase |
| ACER 1-2-3 | Alkaline CDases |
| SphK1-2 | sphingosine kinases 1 and 2 |
| HSV-1 | Herpes simplex virus |
| GCS | glucosylceramide synthase |
| SERINC5 | Serine Incorporator 5 |
| RSV | resveratrol |
| mTORC1 | mTOR Complex 1 |
References
- World Health Organization. Coronavirus Disease (COVID-19). Situation Reports. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 13 February 2022).
- Gaggini, M.; Pingitore, A.; Vassalle, C. Plasma Ceramides Pathophysiology, Measurements, Challenges, and Opportunities. Metabolites 2021, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Carpinteiro, A.; Edwards, M.J.; Hoffmann, M.; Kochs, G.; Gripp, B.; Weigang, S.; Adams, C.; Carpinteiro, E.; Gulbins, A.; Keitsch, S.; et al. Pharmacological Inhibition of Acid Sphingomyelinase Prevents Uptake of SARS-CoV-2 by Epithelial Cells. Cell Rep. Med. 2020, 1, 100142. [Google Scholar] [CrossRef] [PubMed]
- Carpinteiro, A.; Gripp, B.; Hoffmann, M.; Pöhlmann, S.; Hoertel, N.; Edwards, M.J.; Kamler, M.; Kornhuber, J.; Becker, K.A.; Gulbins, E. Inhibition of acid sphingomyelinase by ambroxol prevents SARS-CoV-2 entry into epithelial cells. J. Biol. Chem. 2021, 296, 100701. [Google Scholar] [CrossRef]
- Vitner, E.B.; Achdout, H.; Avraham, R.; Politi, B.; Cherry, L.; Tamir, H.; Yahalom-Ronen, Y.; Paran, N.; Melamed, S.; Erez, N.; et al. Glucosylceramide synthase inhibitors prevent replication of SARS-CoV-2 and influenza virus. J. Biol. Chem. 2021, 296, 100470. [Google Scholar] [CrossRef]
- Torretta, E.; Garziano, M.; Poliseno, M.; Capitanio, D.; Biasin, M.; Santantonio, T.A.; Clerici, M.; Lo Caputo, S.; Trabattoni, D.; Gelfi, C. Severity of COVID-19 Patients Predicted by Serum Sphingolipids Signature. Int. J. Mol. Sci. 2021, 22, 10198. [Google Scholar] [CrossRef]
- Marfia, G.; Navone, S.; Guarnaccia, L.; Campanella, R.; Mondoni, M.; Locatelli, M.; Barassi, A.; Fontana, L.; Palumbo, F.; Garzia, E.; et al. Decreased serum level of sphingosine-1-phosphate: A novel predictor of clinical severity in COVID-19. EMBO Mol. Med. 2021, 13, e13424. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Batiha, G.E.; Al-Gareeb, A.I.; Al-Harcan, N.A.H.; Welson, N.N. Receptor-dependent effects of sphingosine-1-phosphate (S1P) in COVID-19: The black side of the moon. Mol. Cell. Biochem. 2023, 478, 2271–2279. [Google Scholar] [CrossRef]
- Pizzato, M.; Baraldi, C.; Boscato Sopetto, G.; Finozzi, D.; Gentile, C.; Gentile, M.D.; Marconi, R.; Paladino, D.; Raoss, A.; Riedmiller, I.; et al. SARS-CoV-2 and the Host Cell: A Tale of Interactions. Front. Virol. 2022, 1, 81538801. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Sternberg, A.; Naujokat, C. Structural features of coronavirus SARS-CoV-2 spike protein: Targets for vaccination. Life Sci. 2020, 257, 118056. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Barua, A.; Grot, N.; Plawski, A. The basis of mink susceptibility to SARS-CoV-2 infection. J. Appl. Genet. 2022, 63, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Eastman, R.T.; Roth, J.S.; Brimacombe, K.R.; Simeonov, A.; Shen, M.; Patnaik, S.; Hall, M.D. Remdesivir: A Review of Its Discovery and Development Leading to Emergency Use Authorization for Treatment of COVID-19. ACS Cent. Sci. 2020, 6, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Bar-On, Y.M.; Gleizer, S.; Bernshtein, B.; Flamholz, A.; Phillips, R.; Milo, R. The total number and mass of SARS-CoV-2 virions. Proc. Natl. Acad. Sci. USA 2021, 118, e2024815118. [Google Scholar] [CrossRef]
- Meacci, E.; Pierucci, F.; Garcia-Gil, M. Skeletal Muscle and COVID-19: The Potential Involvement of Bioactive Sphingolipids. Biomedicines 2022, 10, 1068. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Gaggini, M.; Fenizia, S.; Vassalle, C. Sphingolipid Levels and Signaling via Resveratrol and Antioxidant Actions in Cardiometabolic Risk and Disease. Antioxidants 2023, 12, 1102. [Google Scholar] [CrossRef]
- Törnquist, K.; Asghar, M.Y.; Srinivasan, V.; Korhonen, L.; Lindholm, D. Sphingolipids as Modulators of SARS-CoV-2 Infection. Front. Cell Dev. Biol. 2021, 9, 689854. [Google Scholar] [CrossRef]
- Futerman, A.H.; Hannun, Y.A. The complex life of simple sphingolipids. EMBO Rep. 2004, 5, 777–782. [Google Scholar] [CrossRef]
- Ryan, E.; Nguyen, C.Q.N.; Shiea, C.; Reid, G.E. Detailed Structural Characterization of Sphingolipids via 193 nm Ultraviolet Photodissociation and Ultra High Resolution Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2017, 28, 1406–1419. [Google Scholar] [CrossRef]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Rechberger, G.N. Sphingoid Bases. In Encyclopedia of Lipidomics; Wenk, M.R., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–2. [Google Scholar] [CrossRef]
- Hirabayashi, Y.; Igarashi, Y.; Merrill, A.H. Sphingolipids Synthesis, Transport and Cellular Signaling. In Sphingolipid Biology; Hirabayashi, Y., Igarashi, Y., Merrill, A.H., Eds.; Springer: Tokyo, Japan, 2006; pp. 3–22. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The sphingolipid salvage pathway in ceramide metabolism and signaling. Cell. Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Kraveka, J.M.; Hannun, Y.A. Bioactive Sphingolipids: An Overview on Ceramide, Ceramide 1-Phosphate Dihydroceramide, Sphingosine, Sphingosine 1-Phosphate. In Handbook of Neurochemistry and Molecular Neurobiology: Neural Lipids; Lajtha, A., Tettamanti, G., Goracci, G., Eds.; Springer: Boston, MA, USA, 2009; pp. 373–383. [Google Scholar] [CrossRef]
- Quinville, B.M.; Deschenes, N.M.; Ryckman, A.E.; Walia, J.S. A Comprehensive Review: Sphingolipid Metabolism and Implications of Disruption in Sphingolipid Homeostasis. Int. J. Mol. Sci. 2021, 22, 5793. [Google Scholar] [CrossRef] [PubMed]
- Tan-Chen, S.; Guitton, J.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism and Signaling in Skeletal Muscle: From Physiology to Physiopathology. Front. Endocrinol. 2020, 11, 491. [Google Scholar] [CrossRef]
- Ferreira, N.S.; Engelsby, H.; Neess, D.; Kelly, S.L.; Volpert, G.; Merrill, A.H.; Futerman, A.H.; Færgeman, N.J. Regulation of very-long acyl chain ceramide synthesis by acyl-CoA-binding protein. J. Biol. Chem. 2017, 292, 7588–7597. [Google Scholar] [CrossRef]
- Hanada, K.; Kumagai, K.; Yasuda, S.; Miura, Y.; Kawano, M.; Fukasawa, M.; Nishijima, M. Molecular machinery for non-vesicular trafficking of ceramide. Nature 2003, 426, 803–809. [Google Scholar] [CrossRef]
- Huitema, K.; van den Dikkenberg, J.; Brouwers, J.F.; Holthuis, J.C. Identification of a family of animal sphingomyelin synthases. EMBO J. 2004, 23, 33–44. [Google Scholar] [CrossRef]
- Diaz Escarcega, R.; McCullough, L.D.; Tsvetkov, A.S. The Functional Role of Sphingosine Kinase 2. Front. Mol. Biosci. 2021, 8, 683767. [Google Scholar] [CrossRef]
- Di Pietro, P.; Izzo, C.; Abate, A.C.; Iesu, P.; Rusciano, M.R.; Venturini, E.; Visco, V.; Sommella, E.; Ciccarelli, M.; Carrizzo, A.; et al. The Dark Side of Sphingolipids: Searching for Potential Cardiovascular Biomarkers. Biomolecules 2023, 13, 168. [Google Scholar] [CrossRef]
- Hammerschmidt, P.; Brüning, J.C. Contribution of specific ceramides to obesity-associated metabolic diseases. Cell. Mol. Life Sci. 2022, 79, 395. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Peng, Q.; Huang, Y. Potential therapeutic targets for atherosclerosis in sphingolipid metabolism. Clin. Sci. 2019, 133, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, Y.M.; Al Aidaros, A.; Valappil, A.; Ali, B.R.; Akawi, N. Role of Ceramides in the Molecular Pathogenesis and Potential Therapeutic Strategies of Cardiometabolic Diseases: What we Know so Far. Front. Cell Dev. Biol. 2021, 9, 816301. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, N.; Becker, K.A. Ceramide and Related Molecules in Viral Infections. Int. J. Mol. Sci. 2021, 22, 5676. [Google Scholar] [CrossRef] [PubMed]
- Teichgräber, V.; Ulrich, M.; Endlich, N.; Riethmüller, J.; Wilker, B.; De Oliveira-Munding, C.C.; van Heeckeren, A.M.; Barr, M.L.; von Kürthy, G.; Schmid, K.W.; et al. Ceramide accumulation mediates inflammation, cell death and infection susceptibility in cystic fibrosis. Nat. Med. 2008, 14, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Oskeritzian, C.A.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate in allergic responses, asthma and anaphylaxis. Pharmacol. Ther. 2007, 115, 390–399. [Google Scholar] [CrossRef]
- Tringali, C.; Giussani, P. Ceramide and Sphingosine-1-Phosphate in Neurodegenerative Disorders and Their Potential Involvement in Therapy. Int. J. Mol. Sci. 2022, 23, 7806. [Google Scholar] [CrossRef]
- Kornhuber, J.; Hoertel, N.; Gulbins, E. The acid sphingomyelinase/ceramide system in COVID-19. Mol. Psychiatry 2022, 27, 307–314. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, D.X.; Tam, J.P. Lipid rafts are involved in SARS-CoV entry into Vero E6 cells. Biochem. Biophys. Res. Commun. 2008, 369, 344–349. [Google Scholar] [CrossRef]
- Teissier, E.; Pécheur, E.I. Lipids as modulators of membrane fusion mediated by viral fusion proteins. Eur. Biophys. J. 2007, 36, 887–899. [Google Scholar] [CrossRef]
- Dissanayake, T.K.; Yan, B.; Ng, A.C.; Zhao, H.; Chan, G.; Yip, C.C.; Sze, K.H.; To, K.K. Differential role of sphingomyelin in influenza virus, rhinovirus and SARS-CoV-2 infection of Calu-3 cells. J. Gen. Virol. 2021, 102, 001593. [Google Scholar] [CrossRef]
- Moolamalla, S.T.R.; Balasubramanian, R.; Chauhan, R.; Priyakumar, U.D.; Vinod, P.K. Host metabolic reprogramming in response to SARS-CoV-2 infection: A systems biology approach. Microb. Pathog. 2021, 158, 105114. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Ventura, A.E.; Varela, A.R.; Silva, L.C. Tackling the biophysical properties of sphingolipids to decipher their biological roles. Biol. Chem. 2015, 396, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.J.; Becker, K.A.; Gripp, B.; Hoffmann, M.; Keitsch, S.; Wilker, B.; Soddemann, M.; Gulbins, A.; Carpinteiro, E.; Patel, S.H.; et al. Sphingosine prevents binding of SARS-CoV-2 spike to its cellular receptor ACE2. J. Biol. Chem. 2020, 295, 15174–15182. [Google Scholar] [CrossRef] [PubMed]
- Vitner, E.B.; Avraham, R.; Politi, B.; Melamed, S.; Israely, T. Elevation in sphingolipid upon SARS-CoV-2 infection: Possible implications for COVID-19 pathology. Life Sci. Alliance 2022, 5, e202101168. [Google Scholar] [CrossRef]
- Lang, J.; Bohn, P.; Bhat, H.; Jastrow, H.; Walkenfort, B.; Cansiz, F.; Fink, J.; Bauer, M.; Olszewski, D.; Ramos-Nascimento, A.; et al. Acid ceramidase of macrophages traps herpes simplex virus in multivesicular bodies and protects from severe disease. Nat. Commun. 2020, 11, 1338. [Google Scholar] [CrossRef]
- Geiger, N.; Kersting, L.; Schlegel, J.; Stelz, L.; Fähr, S.; Diesendorf, V.; Roll, V.; Sostmann, M.; König, E.M.; Reinhard, S.; et al. The Acid Ceramidase Is a SARS-CoV-2 Host Factor. Cells 2022, 11, 2532. [Google Scholar] [CrossRef]
- Yan, B.; Yuan, S.; Cao, J.; Fung, K.; Lai, P.M.; Yin, F.; Sze, K.H.; Qin, Z.; Xie, Y.; Ye, Z.W.; et al. Phosphatidic acid phosphatase 1 impairs SARS-CoV-2 replication by affecting the glycerophospholipid metabolism pathway. Int. J. Biol. Sci. 2022, 18, 4744–4755. [Google Scholar] [CrossRef]
- Menck, K.; Sönmezer, C.; Worst, T.S.; Schulz, M.; Dihazi, G.H.; Streit, F.; Erdmann, G.; Kling, S.; Boutros, M.; Binder, C.; et al. Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J. Extracell. Vesicles 2017, 6, 1378056. [Google Scholar] [CrossRef]
- Mesquita, F.S.; Abrami, L.; Sergeeva, O.; Turelli, P.; Qing, E.; Kunz, B.; Raclot, C.; Paz Montoya, J.; Abriata, L.A.; Gallagher, T.; et al. S-acylation controls SARS-CoV-2 membrane lipid organization and enhances infectivity. Dev. Cell 2021, 56, 2790–2807.e8. [Google Scholar] [CrossRef]
- Timilsina, U.; Umthong, S.; Ivey, E.B.; Waxman, B.; Stavrou, S. SARS-CoV-2 ORF7a potently inhibits the antiviral effect of the host factor SERINC5. Nat. Commun. 2022, 13, 2935. [Google Scholar] [CrossRef] [PubMed]
- Göggel, R.; Winoto-Morbach, S.; Vielhaber, G.; Imai, Y.; Lindner, K.; Brade, L.; Brade, H.; Ehlers, S.; Slutsky, A.S.; Schütze, S.; et al. PAF-mediated pulmonary edema: A new role for acid sphingomyelinase and ceramide. Nat. Med. 2004, 10, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Cartier, A.; Hla, T. Sphingosine 1-phosphate: Lipid signaling in pathology and therapy. Science 2019, 366, eaar5551. [Google Scholar] [CrossRef] [PubMed]
- Petrache, I.; Pujadas, E.; Ganju, A.; Serban, K.A.; Borowiec, A.; Babbs, B.; Bronova, I.A.; Egersdorf, N.; Hume, P.S.; Goel, K.; et al. Marked elevations in lung and plasma ceramide in COVID-19 linked to microvascular injury. JCI Insight 2023, 8, e156104. [Google Scholar] [CrossRef] [PubMed]
- Havulinna, A.S.; Sysi-Aho, M.; Hilvo, M.; Kauhanen, D.; Hurme, R.; Ekroos, K.; Salomaa, V.; Laaksonen, R. Circulating Ceramides Predict Cardiovascular Outcomes in the Population-Based FINRISK 2002 Cohort. Arter. Thromb. Vasc. Biol. 2016, 36, 2424–2430. [Google Scholar] [CrossRef]
- Anroedh, S.; Hilvo, M.; Akkerhuis, K.M.; Kauhanen, D.; Koistinen, K.; Oemrawsingh, R.; Serruys, P.; van Geuns, R.J.; Boersma, E.; Laaksonen, R.; et al. Plasma concentrations of molecular lipid species predict long-term clinical outcome in coronary artery disease patients. J. Lipid Res. 2018, 59, 1729–1737. [Google Scholar] [CrossRef]
- Michelucci, E.; Rocchiccioli, S.; Gaggini, M.; Ndreu, R.; Berti, S.; Vassalle, C. Ceramides and Cardiovascular Risk Factors, Inflammatory Parameters and Left Ventricular Function in AMI Patients. Biomedicines 2022, 10, 429. [Google Scholar] [CrossRef]
- Marathe, S.; Schissel, S.L.; Yellin, M.J.; Beatini, N.; Mintzer, R.; Williams, K.J.; Tabas, I. Human vascular endothelial cells are a rich and regulatable source of secretory sphingomyelinase. Implications for early atherogenesis and ceramide-mediated cell signaling. J. Biol. Chem. 1998, 273, 4081–4088. [Google Scholar] [CrossRef]
- Schissel, S.L.; Keesler, G.A.; Schuchman, E.H.; Williams, K.J.; Tabas, I. The cellular trafficking and zinc dependence of secretory and lysosomal sphingomyelinase, two products of the acid sphingomyelinase gene. J. Biol. Chem. 1998, 273, 18250–18259. [Google Scholar] [CrossRef]
- Janneh, A.H.; Kassir, M.F.; Dwyer, C.J.; Chakraborty, P.; Pierce, J.S.; Flume, P.A.; Li, H.; Nadig, S.N.; Mehrotra, S.; Ogretmen, B. Alterations of lipid metabolism provide serologic biomarkers for the detection of asymptomatic versus symptomatic COVID-19 patients. Sci. Rep. 2021, 11, 14232. [Google Scholar] [CrossRef]
- Song, J.-W.; Lam, S.M.; Fan, X.; Cao, W.-J.; Wang, S.-Y.; Tian, H.; Chua, G.H.; Zhang, C.; Meng, F.-P.; Xu, Z.; et al. Omics-Driven Systems Interrogation of Metabolic Dysregulation in COVID-19 Pathogenesis. Cell Metab. 2020, 32, 188–202.e5. [Google Scholar] [CrossRef] [PubMed]
- Khodadoust, M.M. Inferring a causal relationship between ceramide levels and COVID-19 respiratory distress. Sci. Rep. 2021, 11, 20866. [Google Scholar] [CrossRef] [PubMed]
- Ghidoni, R.; Caretti, A.; Signorelli, P. Role of Sphingolipids in the Pathobiology of Lung Inflammation. Mediat. Inflamm. 2015, 2015, 487508. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.W.; Ooi, E.M.; Watts, G.F.; Chan, D.C.; Weir, J.M.; Meikle, P.J.; Barrett, P.H. Dose-dependent effects of rosuvastatin on the plasma sphingolipidome and phospholipidome in the metabolic syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E2335–E2340. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Wen, X.Q.; Ren, N.; Yang, L.; Gao, B. Lipidomic profile in patients with a very high risk of atherosclerotic cardiovascular disease on PCSK9 inhibitor therapy. Rev. Cardiovasc. Med. 2021, 22, 461–467. [Google Scholar] [CrossRef]
- Palacios-Rápalo, S.N.; De Jesús-González, L.A.; Cordero-Rivera, C.D.; Farfan-Morales, C.N.; Osuna-Ramos, J.F.; Martínez-Mier, G.; Quistián-Galván, J.; Muñoz-Pérez, A.; Bernal-Dolores, V.; Del Ángel, R.M.; et al. Cholesterol-Rich Lipid Rafts as Platforms for SARS-CoV-2 Entry. Front. Immunol. 2021, 12, 796855. [Google Scholar] [CrossRef]
- Sorice, M.; Misasi, R.; Riitano, G.; Manganelli, V.; Martellucci, S.; Longo, A.; Garofalo, T.; Mattei, V. Targeting Lipid Rafts as a Strategy Against Coronavirus. Front. Cell Dev. Biol. 2020, 8, 618296. [Google Scholar] [CrossRef]
- Mazzone, A.; Clemente, A.; Chiappino, D.; Berti, S.; Vassalle, C. Double Face of Statins at the Crossroad of Coronary Atherosclerotic Plaque and Aortic Valve Calcification? JACC Cardiovasc. Imaging 2018, 11, 1930–1931. [Google Scholar] [CrossRef]
- Vahedian-Azimi, A.; Mohammadi, S.M.; Heidari Beni, F.; Banach, M.; Guest, P.C.; Jamialahmadi, T.; Sahebkar, A. Improved COVID-19 ICU admission and mortality outcomes following treatment with statins: A systematic review and meta-analysis. Arch. Med. Sci. 2021, 17, 579–595. [Google Scholar] [CrossRef]
- Lao, U.S.; Law, C.F.; Baptista-Hon, D.T.; Tomlinson, B. Systematic Review and Meta-Analysis of Statin Use and Mortality, Intensive Care Unit Admission and Requirement for Mechanical Ventilation in COVID-19 Patients. J. Clin. Med. 2022, 11, 5454. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, G.; Han, D. Statins in hospitalized COVID-19 patients: A systematic review and meta-analysis of randomized controlled trials. J. Med. Virol. 2023, 95, e28823. [Google Scholar] [CrossRef] [PubMed]
- Vuorio, A.; Kovanen, P.T. PCSK9 inhibitors for COVID-19: An opportunity to enhance the antiviral action of interferon in patients with hypercholesterolaemia. J. Intern. Med. 2021, 289, 749–751. [Google Scholar] [CrossRef] [PubMed]
- Navarese, E.P.; Podhajski, P.; Gurbel, P.A.; Grzelakowska, K.; Ruscio, E.; Tantry, U.; Magielski, P.; Kubica, A.; Niezgoda, P.; Adamski, P.; et al. PCSK9 Inhibition During the Inflammatory Stage of SARS-CoV-2 Infection. J. Am. Coll. Cardiol. 2023, 81, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Domi, E.; Hoxha, M.; Kolovani, E.; Tricarico, D.; Zappacosta, B. The Importance of Nutraceuticals in COVID-19: What’s the Role of Resveratrol? Molecules 2022, 27, 2376. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Ye, Q.; Duan, X.; Xiang, T. Network Pharmacology Reveals That Resveratrol Can Alleviate COVID-19-Related Hyperinflammation. Dis. Markers 2021, 2021, 4129993. [Google Scholar] [CrossRef] [PubMed]
- Nixon, G.F. Sphingolipids in inflammation: Pathological implications and potential therapeutic targets. Br. J. Pharmacol. 2009, 158, 982–993. [Google Scholar] [CrossRef]
- Chiba, K. Discovery of fingolimod based on the chemical modification of a natural product from the fungus, Isaria sinclairii. J. Antibiot. 2020, 73, 666–678. [Google Scholar] [CrossRef]
- Foerch, C.; Friedauer, L.; Bauer, B.; Wolf, T.; Adam, E.H. Severe COVID-19 infection in a patient with multiple sclerosis treated with fingolimod. Mult. Scler. Relat. Disord. 2020, 42, 102180. [Google Scholar] [CrossRef]
- Gomez-Mayordomo, V.; Montero-Escribano, P.; Matías-Guiu, J.A.; González-García, N.; Porta-Etessam, J.; Matías-Guiu, J. Clinical exacerbation of SARS-CoV2 infection after fingolimod withdrawal. J. Med. Virol. 2021, 93, 546–549. [Google Scholar] [CrossRef]
- Huwiler, A.; Zangemeister-Wittke, U. The sphingosine 1-phosphate receptor modulator fingolimod as a therapeutic agent: Recent findings and new perspectives. Pharmacol. Ther. 2018, 185, 34–49. [Google Scholar] [CrossRef]
- McGowan, E.M.; Haddadi, N.; Nassif, N.T.; Lin, Y. Targeting the SphK-S1P-SIPR Pathway as a Potential Therapeutic Approach for COVID-19. Int. J. Mol. Sci. 2020, 21, 7189. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Maines, L.W.; Keller, S.N.; Katz Ben-Yair, V.; Fathi, R.; Plasse, T.F.; Levitt, M.L. Recent Progress in the Development of Opaganib for the Treatment of Covid-19. Drug Des. Dev. Ther. 2022, 16, 2199–2211. [Google Scholar] [CrossRef] [PubMed]
- Orienti, I.; Francescangeli, F.; De Angelis, M.L.; Fecchi, K.; Bongiorno-Borbone, L.; Signore, M.; Peschiaroli, A.; Boe, A.; Bruselles, A.; Costantino, A.; et al. A new bioavailable fenretinide formulation with antiproliferative, antimetabolic, and cytotoxic effects on solid tumors. Cell Death Dis. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Potenza, R.L.; Lodeserto, P.; Orienti, I. Fenretinide in Cancer and Neurological Disease: A Two-Face Janus Molecule. Int. J. Mol. Sci. 2022, 23, 7426. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Mahmood, S.; Morrice, N.; Kamli-Salino, S.; Dekeryte, R.; Hoffmann, P.A.; Doherty, M.K.; Whitfield, P.D.; Delibegović, M.; Mody, N. Fenretinide inhibits obesity and fatty liver disease but induces Smpd3 to increase serum ceramides and worsen atherosclerosis in LDLR−/− mice. Sci. Rep. 2023, 13, 3937. [Google Scholar] [CrossRef]
- Yu, H.; Valerio, M.; Bielawski, J. Fenretinide inhibited de novo ceramide synthesis and proinflammatory cytokines induced by Aggregatibacter actinomycetemcomitans. J. Lipid Res. 2013, 54, 189–201. [Google Scholar] [CrossRef]
- Hayashi, Y.; Tsuchiya, K.; Yamamoto, M.; Nemoto-Sasaki, Y.; Tanigawa, K.; Hama, K.; Ueda, Y.; Tanikawa, T.; Gohda, J.; Maeda, K.; et al. N-(4-Hydroxyphenyl) Retinamide Suppresses SARS-CoV-2 Spike Protein-Mediated Cell-Cell Fusion by a Dihydroceramide Δ4-Desaturase 1-Independent Mechanism. J. Virol. 2021, 95, e0080721. [Google Scholar] [CrossRef]
- Orienti, I.; Gentilomi, G.A.; Farruggia, G. Pulmonary Delivery of Fenretinide: A Possible Adjuvant Treatment In COVID-19. Int. J. Mol. Sci. 2020, 21, 3812. [Google Scholar] [CrossRef]
- Veronesi, U.; Mariani, L.; Decensi, A.; Formelli, F.; Camerini, T.; Miceli, R.; Di Mauro, M.G.; Costa, A.; Marubini, E.; Sporn, M.B.; et al. Fifteen-year results of a randomized phase III trial of fenretinide to prevent second breast cancer. Ann. Oncol. 2006, 17, 1065–1071. [Google Scholar] [CrossRef]
- Mir, I.H.; Thirunavukkarasu, C. The relevance of acid sphingomyelinase as a potential target for therapeutic intervention in hepatic disorders: Current scenario and anticipated trends. Arch. Toxicol. 2023, 97, 2069–2087. [Google Scholar] [CrossRef]
- Naz, A.; Asif, S.; Alwutayd, K.M.; Sarfaraz, S.; Abbasi, S.W.; Abbasi, A.; Alenazi, A.M.; Hasan, M.E. Repurposing FIASMAs against Acid Sphingomyelinase for COVID-19: A Computational Molecular Docking and Dynamic Simulation Approach. Molecules 2023, 28, 2989. [Google Scholar] [CrossRef] [PubMed]
- Hoertel, N.; Sánchez-Rico, M.; Gulbins, E.; Kornhuber, J.; Carpinteiro, A.; Lenze, E.J.; Reiersen, A.M.; Abellán, M.; de la Muela, P.; Vernet, R.; et al. Association Between FIASMAs and Reduced Risk of Intubation or Death in Individuals Hospitalized for Severe COVID-19: An Observational Multicenter Study. Clin. Pharmacol. Ther. 2021, 110, 1498–1511. [Google Scholar] [CrossRef] [PubMed]
- Darquennes, G.; Le Corre, P.; Le Moine, O.; Loas, G. Association between Functional Inhibitors of Acid Sphingomyelinase (FIASMAs) and Reduced Risk of Death in COVID-19 Patients: A Retrospective Cohort Study. Pharmaceuticals 2021, 14, 226. [Google Scholar] [CrossRef]
- Loas, G.; Le Corre, P. Update on Functional Inhibitors of Acid Sphingomyelinase (FIASMAs) in SARS-CoV-2 Infection. Pharmaceuticals 2021, 14, 691. [Google Scholar] [CrossRef] [PubMed]
- Gulbins, A.; Schumacher, F.; Becker, K.A.; Wilker, B.; Soddemann, M.; Boldrin, F.; Müller, C.P.; Edwards, M.J.; Goodman, M.; Caldwell, C.C.; et al. Antidepressants act by inducing autophagy controlled by sphingomyelin-ceramide. Mol. Psychiatry 2018, 23, 2324–2346. [Google Scholar] [CrossRef]
- Pauletto, P.J.T.; Delgado, C.P.; da Rocha, J.B.T. Acid sphingomyelinase (ASM) and COVID-19: A review of the potential use of ASM inhibitors against SARS-CoV-2. Cell Biochem. Funct. 2023, 41, 284–295. [Google Scholar] [CrossRef]
- Bedia, C.; Triola, G.; Casas, J.; Llebaria, A.; Fabriàs, G. Analogs of the dihydroceramide desaturase inhibitor GT11 modified at the amide function: Synthesis and biological activities. Org. Biomol. Chem. 2005, 3, 3707–3712. [Google Scholar] [CrossRef]
- Holliday, M.W., Jr.; Cox, S.B.; Kang, M.H.; Maurer, B.J. C22:0- and C24:0-dihydroceramides confer mixed cytotoxicity in T-cell acute lymphoblastic leukemia cell lines. PLoS ONE 2013, 8, e74768. [Google Scholar] [CrossRef]
- Hernández-Tiedra, S.; Fabriàs, G.; Dávila, D.; Salanueva, Í.J.; Casas, J.; Montes, L.R.; Antón, Z.; García-Taboada, E.; Salazar-Roa, M.; Lorente, M.; et al. Dihydroceramide accumulation mediates cytotoxic autophagy of cancer cells via autolysosome destabilization. Autophagy 2016, 12, 2213–2229. [Google Scholar] [CrossRef]
- Jiménez de Oya, N.; San-Félix, A.; Casasampere, M.; Blázquez, A.B.; Mingo-Casas, P.; Escribano-Romero, E.; Calvo-Pinilla, E.; Poderoso, T.; Casas, J.; Saiz, J.C.; et al. Pharmacological Elevation of Cellular Dihydrosphingomyelin Provides a Novel Antiviral Strategy against West Nile Virus Infection. Antimicrob. Agents Chemother. 2023, 67, e0168722. [Google Scholar] [CrossRef]
- Camacho, L.; Simbari, F.; Garrido, M.; Abad, J.L.; Casas, J.; Delgado, A.; Fabriàs, G. 3-Deoxy-3,4-dehydro analogs of XM462. Preparation and activity on sphingolipid metabolism and cell fate. Bioorganic Med. Chem. 2012, 20, 3173–3179. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Gutiérrez, L.; Benito-Cuesta, I.; Abad, J.L.; Casas, J.; Fábrias, G.; Wandosell, F. Dihydroceramide Desaturase 1 Inhibitors Reduce Amyloid-β Levels in Primary Neurons from an Alzheimer’s Disease Transgenic Model. Pharm. Res. 2018, 35, 49. [Google Scholar] [CrossRef] [PubMed]
- French, K.J.; Schrecengost, R.S.; Lee, B.D.; Zhuang, Y.; Smith, S.N.; Eberly, J.L.; Yun, J.K.; Smith, C.D. Discovery and evaluation of inhibitors of human sphingosine kinase. Cancer Res. 2003, 63, 5962–5969. [Google Scholar] [PubMed]
- Vijayan, M.; Seo, Y.J.; Pritzl, C.J.; Squires, S.A.; Alexander, S.; Hahm, B. Sphingosine kinase 1 regulates measles virus replication. Virology 2014, 450–451, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Chithelen, J.; Franke, H.; Länder, N.; Grafen, A.; Schneider-Schaulies, J. The Sphingolipid Inhibitors Ceranib-2 and SKI-II Reduce Measles Virus Replication in Primary Human Lymphocytes: Effects on mTORC1 Downstream Signaling. Front. Physiol. 2022, 13, 856143. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Gao, D.; Fang, Z.Y. Targeting colorectal cancer cells by a novel sphingosine kinase 1 inhibitor PF-543. Biochem. Biophys. Res. Commun. 2016, 470, 728–734. [Google Scholar] [CrossRef]
- Marín-Corral, J.; Rodríguez-Morató, J.; Gomez-Gomez, A.; Pascual-Guardia, S.; Muñoz-Bermúdez, R.; Salazar-Degracia, A.; Pérez-Terán, P.; Restrepo, M.I.; Khymenets, O.; Haro, N.; et al. Metabolic Signatures Associated with Severity in Hospitalized COVID-19 Patients. Int. J. Mol. Sci. 2021, 22, 4794. [Google Scholar] [CrossRef]
- Pyne, S.; Adams, D.R.; Pyne, N.J. Sphingosine 1-phosphate and sphingosine kinases in health and disease: Recent advances. Prog. Lipid Res. 2016, 62, 93–106. [Google Scholar] [CrossRef]
- Hammerschmidt, P.; Ostkotte, D.; Nolte, H.; Gerl, M.J.; Jais, A.; Brunner, H.L.; Sprenger, H.G.; Awazawa, M.; Nicholls, H.T.; Turpin-Nolan, S.M.; et al. CerS6-Derived Sphingolipids Interact with Mff and Promote Mitochondrial Fragmentation in Obesity. Cell 2019, 177, 1536–1552.e23. [Google Scholar] [CrossRef]
- Shaw, J.; Costa-Pinheiro, P.; Patterson, L.; Drews, K.; Spiegel, S.; Kester, M. Novel Sphingolipid-Based Cancer Therapeutics in the Personalized Medicine Era. Adv. Cancer Res. 2018, 140, 327–366. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fenizia, S.; Gaggini, M.; Vassalle, C. The Sphingolipid-Signaling Pathway as a Modulator of Infection by SARS-CoV-2. Curr. Issues Mol. Biol. 2023, 45, 7956-7973. https://doi.org/10.3390/cimb45100503
Fenizia S, Gaggini M, Vassalle C. The Sphingolipid-Signaling Pathway as a Modulator of Infection by SARS-CoV-2. Current Issues in Molecular Biology. 2023; 45(10):7956-7973. https://doi.org/10.3390/cimb45100503
Chicago/Turabian StyleFenizia, Simona, Melania Gaggini, and Cristina Vassalle. 2023. "The Sphingolipid-Signaling Pathway as a Modulator of Infection by SARS-CoV-2" Current Issues in Molecular Biology 45, no. 10: 7956-7973. https://doi.org/10.3390/cimb45100503
APA StyleFenizia, S., Gaggini, M., & Vassalle, C. (2023). The Sphingolipid-Signaling Pathway as a Modulator of Infection by SARS-CoV-2. Current Issues in Molecular Biology, 45(10), 7956-7973. https://doi.org/10.3390/cimb45100503