
Lactic Acid Bacteria-Derived Exopolysaccharides Mitigate the Oxidative Response via the NRF2-KEAP1 Pathway in PC12 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Media and Culture Conditions
2.2. Isolation of EPSs
2.3. The Cell Line Used in the Study
2.4. Reproduction and Storage of PC-12 Cells
2.5. Cell Viability Test
2.6. The Effect of L-EPSs on TDH in PC-12 Cells Induced by OS with H2O2
2.7. Determination of the Effect of L-EPSs on Total Oxidant Status (TOS), and Total Antioxidant Status (TAS) Levels in PC-12 Cells Induced by OS with H2O2
2.8. Determination of the Effect of L-EPSs on Paraoxanase-1 (PON1), Superoxide Dismutase (SOD), Catalase (CAT), and Glutathione Peroxidase (GPx) Antioxidant Enzyme Levels in PC-12 Cells Induced by OS with H2O2
2.9. Determination of the Effect of L-EPSs on Intracellular ROS Level in PC-12 Cells Induced by OS with H2O2 by Flow Cytometry
2.10. Determination of the Effect of L-EPSs on GCLC, HO-1, NRF2, NQO1, and KEAP1 Gene Expression Levels in PC-12 Cells Induced by OS with H2O2 by qRT-PCR
2.11. Determination of GCLC, HO-1, KEAP, NRF2, and NQO1 by an ELISA Assay
2.12. Statistics
3. Results
3.1. Purity and Amount of L-EPSs
3.2. Protective Effect of L-EPSs in PC-12 Cells Induced by OS with H2O2
3.3. Regulatory Effect of L-EPSs on TDH in PC-12 Cells Induced by OS with H2O2
3.4. Regulatory Effect of L-EPSs on TOS and TAS Levels in PC-12 Cells Induced by OS with H2O2
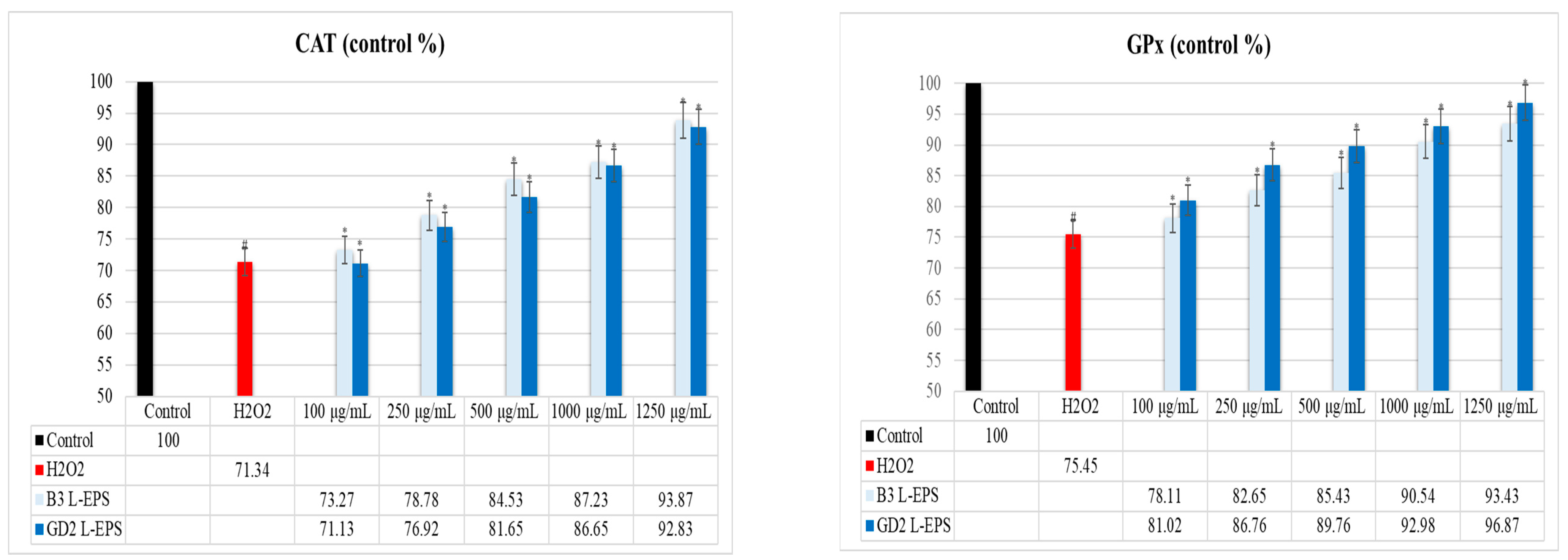
3.5. Inductive Effect of L-EPSs on PON1, SOD, CAT, and GPx Antioxidant Enzyme Levels in PC-12 Cells Induced by OS with H2O2
3.6. Inhibitory Effect of L-EPSs on Intracellular ROS Level in PC-12 Cells Induced by OS with H2O2
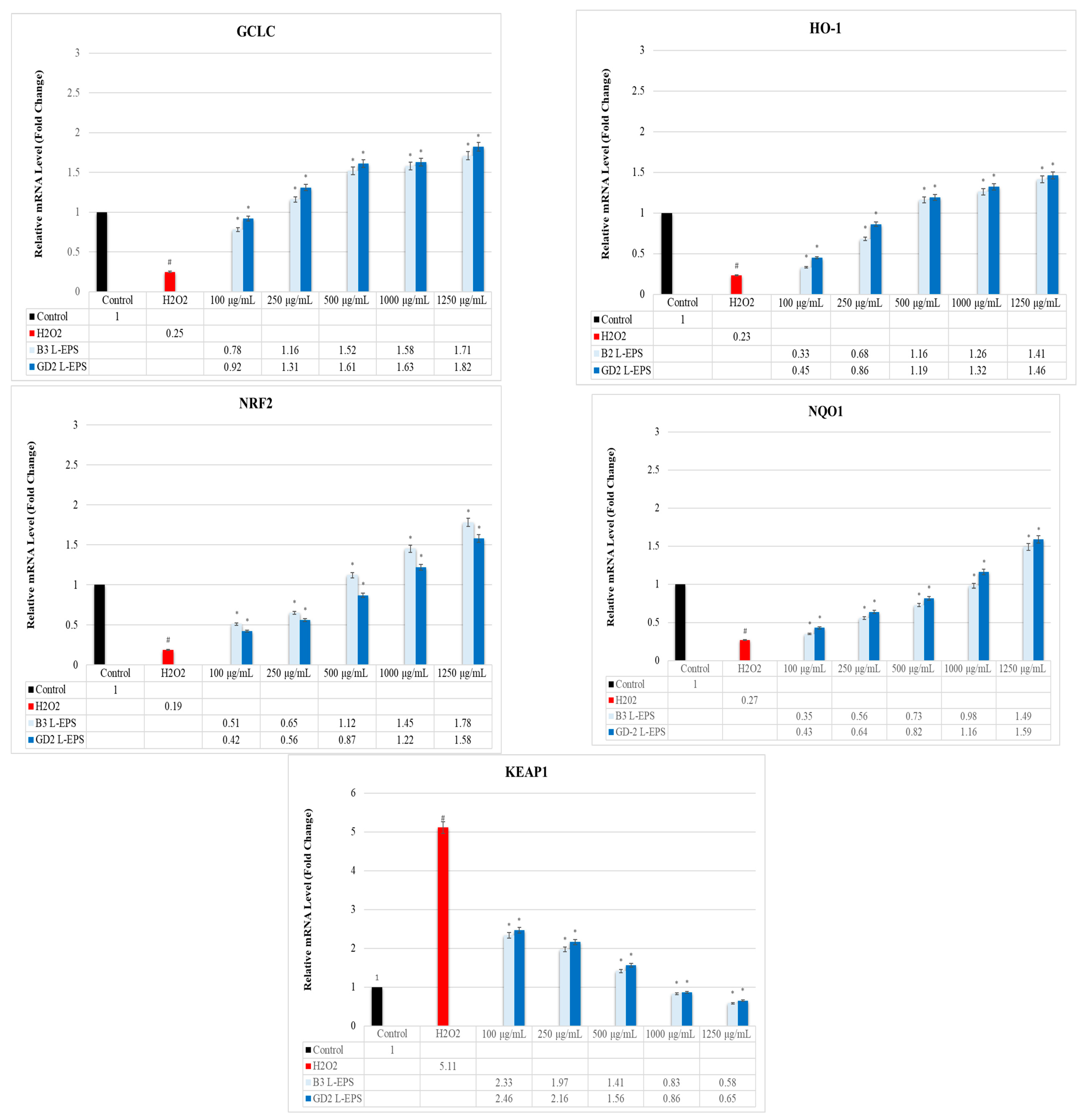
3.7. Regulatory Effect of L-EPSs on GCLC, HO-1, NRF2, NQO1, and KEAP1 Gene Expression Levels in PC-12 Cells Induced by OS with H2O2
3.8. Regulatory Effect of L-EPSs on GCLC, HO-1, NRF2, NQO1, and KEAP1 Protein Expression Levels in PC-12 Cells Induced by OS with H2O2
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, J.; Chitu, V.; Stanley, E.R.; Wszolek, Z.K.; Karrenbauer, V.D.; Harris, R.A. Inhibition of colony stimulating factor-1 receptor (CSF-1R) as a potential therapeutic strategy for neurodegenerative diseases: Opportunities and challenges. Cell. Mol. Life Sci. 2022, 79, 219. [Google Scholar] [CrossRef]
- Wilson, D.M.; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Hallmarks of neurodegenerative diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- Basile, M.S.; Bramanti, P.; Mazzon, E. Inosine in neurodegenerative diseases: From the bench to the bedside. Molecules 2022, 27, 4644. [Google Scholar] [CrossRef] [PubMed]
- Işler, Y.; Kaya, H. Thiol/disulfide homeostasis in patients treated with normobaric or hyperbaric oxygen for carbon monoxide poisoning. Am. J. Emerg. Med. 2022, 59, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Guler, E.M.; Ozkan, B.N.; Yilmaz, N.; Ozgen, F.P.; Taslidere, N.S.; Aktas, S.; Kucuk, O.S. Increased oxidative stress and imbalance dynamic thiol–disulfide homeostasis in Rosacea. Turk. J. Dermatol. 2022, 16, 120. [Google Scholar] [CrossRef]
- Chakkittukandiyil, A.; Sajini, D.V.; Karuppaiah, A.; Selvaraj, D. The principal molecular mechanisms behind the activation of Keap1/Nrf2/ARE pathway leading to neuroprotective action in Parkinson’s disease. Neurochem. Int. 2022, 156, 105325. [Google Scholar] [CrossRef]
- Phukan, B.C.; Roy, R.; Paul, R.; Mazumder, M.K.; Nath, J.; Bhattacharya, P.; Borah, A. Traversing through the cell signaling pathways of neuroprotection by betanin: Therapeutic relevance to Alzheimer’s Disease and Parkinson’s Disease. Metab. Brain Dis. 2023, 38, 805–817. [Google Scholar] [CrossRef]
- Collins, A.E.; Saleh, T.M.; Kalisch, B.E. Naturally occurring antioxidant therapy in Alzheimer’s disease. Antioxidants 2022, 11, 213. [Google Scholar] [CrossRef]
- Toronyi, A.Á.; Giuri, D.; Martiniakova, S.; Tomasini, C. Low-molecular-weight gels as smart materials for the enhancement of antioxidants activity. Cosmetics 2023, 10, 38. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, J.; Jiang, Y.; Wang, D.; Zhao, D.; Wang, C.; Li, M. Bacillus amyloliquefaciens lysate ameliorates photoaging of human skin fibroblasts through NRF2/KEAP1 and TGF-β/SMAD signaling pathways. Appl. Sci. 2022, 12, 9151. [Google Scholar] [CrossRef]
- Wang, J.; Li, M.; Gao, Y.; Li, H.; Fang, L.; Liu, C.; Liu, X.; Min, W. Effects of Exopolysaccharides from Lactiplantibacillus plantarum JLAU103 on intestinal immune response, oxidative stress, and microbial communities in cyclophosphamide-induced immunosuppressed mice. J. Agric. Food Chem. 2022, 70, 2197–2210. [Google Scholar] [CrossRef] [PubMed]
- Sungur, T.; Aslim, B.; Karaaslan, C.; Aktas, B. Impact of Exopolysaccharides (EPSs) of Lactobacillus gasseri strains isolated from human vagina on cervical tumor cells (HeLa). Anaerobe 2017, 47, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars anrelated substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Frengova, G.I.; Simova, E.D.; Beshkova, D.M.; Simov, Z.I. Production and monomer composition of exopolysaccharides by yogurt starter cultures. Can. J. Microbiol. 2000, 46, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, H.; Aslim, B.; Yuksekdag, Z. Assessment of anti-biofilm activity and bifidogenic growth stimulator (BGS) effect of lyophilized exopolysaccharides (lEPSs) from lactobacilli strains. Int. J. Food Prop. 2017, 20, 362–371. [Google Scholar] [CrossRef]
- Kolla, N.; Wei, Z.; Richardson, J.S.; Li, X.M. Amitriptyline and fluoxetine protect PC12 cells from cell death induced by hydrogen peroxide. J. Psychiatry Neurosci. 2005, 30, 196–201. [Google Scholar] [PubMed]
- Wu, H.; Xuan, R.; Li, Y.; Zhang, X.; Wang, Q.; Wang, L. Effects of cadmium exposure on digestive enzymes, antioxidant enzymes, and lipid peroxidation in the freshwater crab Sinopotamon henanense. Environ. Sci. Pollut. Res. 2013, 20, 4085–4092. [Google Scholar] [CrossRef]
- Shen, K.; Wang, Y.; Zhang, Y.; Zhou, H.; Song, Y.; Cao, Z.; Kou, J.; Yu, B. Cocktail of four active components derived from Sheng Mai San inhibits hydrogen peroxide-induced PC12 cell apoptosis linked with the caspase-3/ROCK1/MLC pathway. Rejuvenation Res. 2015, 18, 517–527. [Google Scholar] [CrossRef]
- Kim, E.; Jeon, I.; Kim, J.; Jung, H.-S.; Lee, S. An MTT-based method for quantification of periodontal ligament cell viability. Oral Dis. 2007, 13, 495–499. [Google Scholar] [CrossRef]
- Neshatia, V.; Matin, M.M.; Iranshahi, M.; Bahrami, A.R.; Behravan, J.; Mollazadeh, S.; Rassouli, F.B. Cytotoxicity of vincristine on the 5637 cell line is enhanced by combination with conferone. Z. Naturforsch. C 2009, 64, 317–322. [Google Scholar] [CrossRef]
- Erel, O.; Neselioglu, S. A novel and automated assay for thiol/disulphide homeostasis. Clin. Biochem. 2014, 47, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Küçükgül, A. The antioxidant effects of Ziziphus Jujuba on neurodegeneration. Vet. Mikrobiyol. Derg. 2016, 27, 108–112. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Yumru, M.; Savas, H.A.; Kalenderoglu, A.; Bulut, M.; Celik, H.; Erel, O. Oxidative imbalance in bipolar disorder subtypes: A comparative study. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1070–1074. [Google Scholar] [CrossRef]
- Üstünova, S.; Meral, I.; Koçyiğit, A.; Kılıç, A.; Zeybek, O.; Üyüklü, M.; Meydan, S. Effects of Nigella sativa on plasma oxidative stress, and some apoptotic protein markers in cerebrum and hippocampus in pentylenetetrazol induced-kindling rats. Eur. J. Biol. 2020, 79, 124–131. [Google Scholar] [CrossRef]
- Wu, Y.; Shang, Y.; Sun, S.; Liang, H.; Liu, R. Erythropoietin prevents PC12 cells from 1-methyl-4-phenylpyridinium ion-induced apoptosis via the Akt/GSK-3β/caspase-3 mediated signaling pathway. Apoptosis 2007, 12, 1365–1375. [Google Scholar] [CrossRef]
- Tarozzi, A. Oxidative stress in neurodegenerative diseases: From preclinical studies to clinical applications. J. Clin. Med. 2020, 9, 1223. [Google Scholar] [CrossRef]
- Teixeira, M.I.; Lopes, C.M.; Amaral, M.H.; Costa, P.C. Current insights on lipid nanocarrier-assisted drug delivery in the treatment of neurodegenerative diseases. Eur. J. Pharm. Biopharm. 2020, 149, 192–217. [Google Scholar] [CrossRef]
- Tukenmez, U.; Aktas, B.; Aslim, B.; Yavuz, S. The relationship between the structural characteristics of lactobacilli-EPS and its ability to induce apoptosis in colon cancer cells in vitro. Sci. Rep. 2019, 9, 8268. [Google Scholar] [CrossRef]
- Gientka, I.; Błażejak, S.; Stasiak-Różańska, L.; Chlebowska-Śmigiel, A. Exopolysaccharides from yeast: Insight into optimal conditions for biosynthesis, chemical composition and functional properties? review. Acta Sci. Pol. Technol. Aliment. 2015, 14, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiong, Q.; Lai, X.; Li, X.; Wan, M.; Zhang, J.; Yan, Y.; Cao, M.; Lu, L.; Guan, J.; et al. Molecular modification of polysaccharides and resulting bioactivities. Compr. Rev. Food Sci. Food Saf. 2016, 15, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Tsai, C.L.; Lien, Y.Y.; Lee, M.S.; Sheu, S.C. High molecular weight of polysaccharides from Hericium erinaceus against amyloid beta-induced neurotoxicity. BMC Complement. Altern. Med. 2016, 16, 170. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Sirin, S.; Aslim, B. Characterization of lactic acid bacteria derived exopolysaccharides for use as a defined neuroprotective agent against amyloid beta1–42-induced apoptosis in SH-SY5Y cells. Sci. Rep. 2020, 10, 8124. [Google Scholar] [CrossRef]
- Li, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-derived antioxidant polysaccharides and their pharmacological potential in neurodegenerative diseases. Nutrients 2017, 9, 778. [Google Scholar] [CrossRef]
- Sathiyanarayanan Dineshkumar, K.; Yang, Y.H. Microbial exopolysaccharidemediated synthesis and stabilization of metal nanoparticles. Crit. Rev. Microbiol. 2017, 43, 731–752. [Google Scholar] [CrossRef]
- Vural, G.; Gümüşyayla, Ş.; Deniz, O.; Neşelioğlu, S.; Erel, Ö. Relationship between thiol-disulphide homeostasis and visual evoked potentials in patients with multiple sclerosis. Neurol. Sci. 2019, 40, 385–391. [Google Scholar] [CrossRef]
- Vural, G.; Gumusyayla, S.; Bektas, H.; Deniz, O.; Alisik, M.; Erel, O. Impairment of dynamic thiol–disulphide homeostasis in patients with idiopathic Parkinson’s disease and its relationship with clinical stage of disease. Clin. Neurol. Neurosurg. 2017, 153, 50–55. [Google Scholar] [CrossRef]
- Sabens Liedhegner, E.A.; Gao, X.H.; Mieyal, J.J. Mechanisms of altered redox regulation in neurodegenerative diseases-Focus on S-glutathionylation. Antioxid. Redox Signal. 2012, 16, 543–566. [Google Scholar] [CrossRef]
- Gumusyayla, S.; Vural, G.; Bektas, H.; Deniz, O.; Neselioglu, S.; Erel, O. A novel oxidative stress marker in patients with Alzheimer’s disease: Dynamic thiol–disulphide homeostasis. Acta Neuropsychiatr. 2016, 28, 315–320. [Google Scholar] [CrossRef]
- Kayacan, Y.; Kola, A.Z.; Guandalini, S.; Yazar, H.; Söğüt, M.Ü. The use of probiotics combined with exercise affects thiol/disulfide homeostasis, an oxidative stress parameter. Nutrients 2022, 14, 3555. [Google Scholar] [CrossRef]
- Kütük, A.; Akar, F.; Sadi, G. Changes in hepatic thiol contents and regulation of glutathione S-transferase by high-fructose diet: Effects of kefir and some probiotic bacteria. J. Sci. Perspect. 2023, 3, 127–137. [Google Scholar] [CrossRef]
- Nikitina, E.; Petrova, T.; Sungatullina, A.; Bondar, O.; Kharina, M.; Mikshina, P.; Gavrilova, E.; Kayumov, A. The profile of exopolysaccharides produced by various Lactobacillus species from silage during not-fat milk fermentation. Fermentation 2023, 9, 197. [Google Scholar] [CrossRef]
- Soumya, M.P.; Nampoothiri, K.M. Evaluation of improved biological properties of chemically modified exopolysaccharides from Lactobacillus plantarum BR2. 3 Biotech 2023, 13, 308. [Google Scholar] [CrossRef]
- Zhou, Z.; Zeng, X.; Wu, Z.; Guo, Y.; Pan, D. Relationship of gene-structure–antioxidant ability of exopolysaccharides derived from lactic acid bacteria: A Review. J. Agric. Food Chem. 2023, 71, 9187–9200. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, R.; Li, S.; Wu, M.; Yu, H.; Ge, Q. Metabolism of hydrogen peroxide by Lactobacillus plantarum NJAU-01: A proteomics study. Food Microbiol. 2023, 112, 104246. [Google Scholar] [CrossRef] [PubMed]
- Ayada, C.; Erbay, U.T.; Korkut, Y.; Guleken, Z.; Oner, Z. Evaluation of oxidative system parameters in Alzheimer’s disease before medical treatment. Int. J. Gerontol. 2022, 16, 395–399. [Google Scholar] [CrossRef]
- Tugrul, S.; Koçyiğit, A.; Doğan, R.; Eren, S.B.; Senturk, E.; Ozturan, O.; Ozar, O.F. Total antioxidant status and oxidative stress in recurrent aphthous stomatitis. Int. J. Dermatol. 2016, 55, e130–e135. [Google Scholar] [CrossRef] [PubMed]
- Arikanoglu, A.; Akil, E.; Varol, S.; Yucel, Y.; Yuksel, H.; Cevik, M.U.; Palanci, Y.; Unan, F. Relationship of cognitive performance with prolidase and oxidative stress in Alzheimer disease. Neurol. Sci. 2013, 34, 2117–2121. [Google Scholar] [CrossRef] [PubMed]
- Copoglu, U.S.; Virit, O.; Kokacya, M.H.; Orkmez, M.; Bulbul, F.; Erbagci, A.B.; Semiz, M.; Alpak, G.; Unal, A.; Ari, M.; et al. Increased oxidative stress and oxidative DNA damage in non-remission schizophrenia patients. Psychiatry Res. 2015, 229, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Sirin, S.; Aslim, B. Protective effect of exopolysaccharides from lactic acid bacteria against amyloid beta1-42 induced oxidative stress in SH-SY5Y cells: Involvement of the AKT, MAPK, and NF-κB signaling pathway. Process Biochem. 2021, 106, 50–59. [Google Scholar] [CrossRef]
- Ciltas, A.C.; Toy, C.E.; Güneş, H.; Yaprak, M. Effects of probiotics on GABA/glutamate and oxidative stress in PTZ-induced acute seizure model in rats. Epilepsy Res. 2023, 195, 107190. [Google Scholar] [CrossRef]
- Javed, G.A.; Shazadi, K.; Munir, A.; Javed, S.; Ambreen, H.S.; Arshad, N. Weissella confusa MW051433 and Levilactobacillus brevis MW362788 of human milk origin possess antioxidant competency: In vivo evidence. Process Biochem. 2023, 133, 11–19. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Guo, H.; Cheng, Q.; Abbas, Z.; Tong, Y.; Yang, T.; Zhou, Y.; Zhang, H.; Wei, X.; et al. Optimization of exopolysaccharide produced by Lactobacillus plantarum R301 and its antioxidant and anti-inflammatory activities. Foods 2023, 12, 2481. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.G.; Awady, M.E.E.; Selim, M.S.; Ibrahim, A.Y.; Ibrahim, F.M.; Mohamed, S.S. Characterization of biologically active exopolysaccharide produced by Streptomyces sp. NRCG4 and its anti-Alzheimer efficacy: In-vitro targets. J. Genet. Eng. Biotechnol. 2023, 21, 76. [Google Scholar] [CrossRef]
- Cao, C.; Bian, Y.; Cang, W.; Wu, J.; Wu, R. Structural characterization and hepatoprotective activity of exopolysaccharide from Bacillus velezensis SN-1. J. Sci. Food Agric. 2023, 103, 738–749. [Google Scholar] [CrossRef]
- Prajapati, D.; Bhatt, A.; Gupte, A. Evaluation of bioactive attributes and emulsification potential of exopolysaccharide produced by a brown-rot fungus Fomitopsis meliae AGDP-2. Appl. Biochem. Biotechnol. 2023, 195, 2974–2992. [Google Scholar] [CrossRef]
- Amrutha, T.A.; Beena, A.K. Microbial exopolysaccharides: A promising health booster. J. Phytopharm. 2023, 12, 265–271. [Google Scholar] [CrossRef]
- Mu, S.; Yang, W.; Huang, G. Antioxidant activities and mechanisms of polysaccharides. Chem. Biol. Drug Des. 2021, 97, 628–632. [Google Scholar] [CrossRef]
- Rosa, A.C.; Bruni, N.; Meineri, G.; Corsi, D.; Cavi, N.; Gastaldi, D.; Dosio, F. Strategies to expand the therapeutic potential of superoxide dismutase by exploiting delivery approaches. Int. J. Biol. Macromol. 2021, 168, 846–865. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Liu, R. Oxidative stress response of two fluoroquinolones with catalase and erythrocytes: A combined molecular and cellular study. J. Hazard. Mater. 2013, 252, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Eddaikra, A.; Eddaikra, N. Endogenous enzymatic antioxidant defense and pathologies. IntechOpen 2021, 95504. [Google Scholar] [CrossRef]
- Samani, K.G.; Farrokhi, E. Effects of cumin extract on oxLDL, paraoxanase 1 activity, FBS, total cholesterol, triglycerides, HDL-C, LDL-C, Apo A1, and Apo B in in the patients with hypercholesterolemia. Int. J. Health Sci. 2014, 8, 39–43. [Google Scholar] [CrossRef]
- Ghosh, N.; Ghosh, R.; Mandal, S.C. Antioxidant protection: A promising therapeutic intervention in neurodegenerative disease. Free Radic. Res. 2011, 45, 888–905. [Google Scholar] [CrossRef]
- Obuobi, S.; Karatayev, S.; Chai, C.L.L.; Ee, P.L.R.; Mátyus, P. The role of modulation of antioxidant enzyme systems in the treatment of neurodegenerative diseases. J. Enzym. Inhib. Med. Chem. 2016, 31, 194–204. [Google Scholar] [CrossRef]
- Romani, A.; Trentini, A.; van der Flier, W.M.; Bellini, T.; Zuliani, G.; Cervellati, C.; Teunissen, C.E. Arylesterase activity of paraoxonase-1 in serum and cerebrospinal fluid of patients with Alzheimer’s disease and vascular dementia. Antioxidants 2020, 9, 456. [Google Scholar] [CrossRef]
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of plasma magnesium, copper, zinc, iron and selenium concentrations and some related erythrocyte antioxidant enzyme activities in patients with Alzheimer’s disease. J. Trace Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef]
- Şengül, N.; Işık, S.; Aslım, B.; Uçar, G.; Demirbağ, A.E. The effect of exopolysaccharide-producing probiotic strains on gut oxidative damage in experimental colitis. Dig. Dis. Sci. 2011, 56, 707–714. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef]
- Bhawal, S.; Kumari, A.; Kapila, S.; Kapila, R. Physicochemical characteristics of novel cell-bound exopolysaccharide from probiotic Limosilactobacillus fermentum (MTCC 5898) and its relation to antioxidative activity. J. Agric. Food Chem. 2021, 69, 10338–10349. [Google Scholar] [CrossRef]
- Zhao, H.; Li, J.; Zhang, J.; Wang, X.; Hao, L.; Jia, L. Purification, in vitro antioxidant and in vivo anti-aging activities of exopolysaccharides by Agrocybe cylindracea. Int. J. Biol. Macromol. 2017, 102, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.E.S.; de Vasconcelos, A.S.; da Costa Vilhena, T.; da Silva, T.L.; da Silva Barbosa, A.; Gomes, A.R.Q.; Dolabela, M.F.; Percário, S. Oxidative stress in Alzheimer’s disease: Should we keep trying antioxidant therapies? Cell. Mol. Neurobiol. 2015, 35, 595–614. [Google Scholar] [CrossRef]
- Manoharan, S.; Guillemin, G.J.; Abiramasundari, R.S.; Essa, M.M.; Akbar, M.; Akbar, M.D. The role of reactive oxygen species in the pathogenesis of Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease: A mini review. Oxid. Med. Cell. Longev. 2016, 2016, 8590578. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative stress in neurodegenerative diseases: From molecular mechanisms to clinical applications. Oxid. Med. Cell. Longev. 2017, 2017, 2525967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, Y.; Guo, K.; Tian, H.; Wang, C.; Wang, R.; Chen, Y.; Chen, X.; Zheng, H.; Gao, B.; et al. Macromolecular nanoparticles to attenuate both reactive oxygen species and inflammatory damage for treating Alzheimer’s disease. Bioeng. Transl. Med. 2023, 8, e10459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef]
- Adesulu-Dahunsi, A.T.; Jeyaram, K.; Sanni, A.I.; Banwo, K. Production of exopolysaccharide by strains of Lactobacillus plantarum YO175 and OF101 isolated from traditional fermented cereal beverage. PeerJ 2018, 6, e5326. [Google Scholar] [CrossRef]
- Bylund, J.; Burgess, L.A.; Cescutti, P.; Ernst, R.K.; Speert, D.P. Exopolysaccharides from Burkholderia cenocepacia inhibit neutrophil chemotaxis and scavenge reactive oxygen species. J. Biol. Chem. 2006, 281, 2526–2532. [Google Scholar] [CrossRef]
- Gupta, P.; Diwan, B. Bacterial exopolysaccharide mediated heavy metal removal: A review on biosynthesis, mechanism and remediation strategies. Biotechnol. Rep. 2017, 13, 58–71. [Google Scholar] [CrossRef]
- Xu, R.; Shang, N.; Li, P. In vitro and in vivo antioxidant activity of exopolysaccharide fractions from Bifidobacterium animalis RH. Anaerobe 2011, 17, 226–231. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr. Polym. 2012, 90, 683–689. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Lin, H.Y.; Wang, C.C.; Zhang, M.; Lin, Y.Y.; Huang, F.Y.; Lin, Y.Z.; Tan, G.H. Exopolysaccharide from Paecilomyces lilacinus modulates macrophage activities through the TLR4/NF κB/MAPK pathway. Mol. Med. Rep. 2019, 20, 4943–4952. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohyd. Res. 2020, 487, 107881. [Google Scholar] [CrossRef] [PubMed]
- Adebayo-Tayo, B.; Ishola, R.; Oyewunmi, T. Characterization, antioxidant and immunomodulatory potential on exopolysaccharide produced by wild type and mutant Weissella confusa strains. Biotechnol Rep. 2018, 19, e00271. [Google Scholar] [CrossRef]
- Johnson, D.A.; Johnson, J.A. Nrf2—A therapeutic target for the treatment of neurodegenerative diseases. Free Radic. Biol. Med. 2015, 88, 253–267. [Google Scholar] [CrossRef]
- Deshmukh, P.; Unni, S.; Krishnappa, G.; Padmanabhan, B. The Keap1–Nrf2 pathway: Promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys. Rev. 2017, 9, 41–56. [Google Scholar] [CrossRef]
- Kanninen, K.; Malm, T.M.; Jyrkkänen, H.-K.; Goldsteins, G.; Keksa-Goldsteine, V.; Tanila, H.; Yamamoto, M.; Ylä-Herttuala, S.; Levonen, A.-L.; Koistinaho, J. Nuclear factor erythroid 2-related factor 2 protects against beta amyloid. Mol. Cell Neurosci. 2008, 39, 302–313. [Google Scholar] [CrossRef]
- Gureev, A.P.; Khorolskaya, V.G.; Sadovnikova, I.S.; Shaforostova, E.A.; Cherednichenko, V.R.; Burakova, I.Y.; Plotnikov, E.Y.; Popov, V.N. Age-related decline in Nrf2/ARE signaling is associated with the mitochondrial DNA damage and cognitive impairments. Int. J. Mol. Sci. 2022, 23, 15197. [Google Scholar] [CrossRef] [PubMed]
- Averina, O.V.; Poluektova, E.U.; Marsova, M.V.; Danilenko, V.N. Biomarkers and utility of the antioxidant potential of probiotic Lactobacilli and Bifidobacteria as representatives of the human gut microbiota. Biomedicines 2021, 9, 1340. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer 5′→3′ | Reverse Primer 5′→3′ |
|---|---|---|
| GCLC | GTGGACACCCGATGCAGTAT | TCATCCACCTGGCAACAGTC |
| HO-1 | GCTCTATCGTGCTCGCATGA | AATTCCCACTGCCACGGTC |
| KEAP | TGGGCGTGGCAGTGCTCAAC | GCCCATCGTAGCCTCCTGCG |
| NRF2 | GCTGCCATTAGTCAGTCGCTCTC | ACCGTGCCTTCAGTGTGCTTC |
| NQO1 | ACATCACAGGGGAGCCGAAGGACT | GGCACCCCAAACCAATACAATG |
| GAPDH | CAACTCCCTCAAGATTGTCAGCAA | GGCATGGACTGTGGTCATGA |
| Treatment | Relative Protein Levels (% of Control) | ||||
|---|---|---|---|---|---|
| GCLC | HO-1 | NRF2 | NQO1 | KEAP1 | |
| Control | 100 ± 3 | 100 ± 5 | 100 ± 6 | 100 ± 3 | 100 ± 1 |
| H2O2 | 30 ± 2 # | 33 ± 4 # | 25 ± 3 # | 36 ± 2 # | 225 ± 2 # |
| 100 μg/mL B3 L-EPS | 33± 1 | 34± 5 | 36 ± 7 * | 36± 1 | 217 ± 3 |
| 250 μg/mL B3 L-EPS | 46 ±4 * | 44 ±6 * | 53 ± 1 * | 39 ±5 | 208 ± 1 * |
| 500 μg/mL B3 L-EPS | 59 ± 5 * | 56 ± 1 * | 70 ± 2 * | 51± 4 * | 162 ± 5 * |
| 1000 μg/mL B3 L-EPS | 79 ± 2 * | 76 ± 2 * | 91 ± 4 * | 69 ± 2 * | 133 ± 3 * |
| 1250 μg/mL B3 L-EPS | 93 ± 6 * | 89 ± 3 * | 106 ± 6 * | 82± 1 * | 98 ± 4 * |
| 100 μg/mL GD2 L-EPS | 38± 5 | 35± 1 | 34 ± 2 * | 38± 5 | 220 ± 5 |
| 250 μg/mL GD2 L-EPS | 48 ± 3 * | 46 ±2 * | 51 ± 4 * | 45 ±4 * | 213 ± 3 * |
| 500 μg/mL GD2 L-EPS | 65 ± 1 * | 59 ± 3 * | 68± 1 * | 56± 1 * | 167 ± 1 * |
| 1000 μg/mL GD2 L-EPS | 86 ± 7 * | 79 ± 6 * | 87 ± 3 * | 75 ± 3 * | 137 ± 2 * |
| 1250 μg/mL GD2 L-EPS | 98 ± 2 * | 93 ± 8 * | 102 ± 4 * | 89 ± 4 * | 106 ± 3 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Şirin, S. Lactic Acid Bacteria-Derived Exopolysaccharides Mitigate the Oxidative Response via the NRF2-KEAP1 Pathway in PC12 Cells. Curr. Issues Mol. Biol. 2023, 45, 8071-8090. https://doi.org/10.3390/cimb45100510
Şirin S. Lactic Acid Bacteria-Derived Exopolysaccharides Mitigate the Oxidative Response via the NRF2-KEAP1 Pathway in PC12 Cells. Current Issues in Molecular Biology. 2023; 45(10):8071-8090. https://doi.org/10.3390/cimb45100510
Chicago/Turabian StyleŞirin, Seda. 2023. "Lactic Acid Bacteria-Derived Exopolysaccharides Mitigate the Oxidative Response via the NRF2-KEAP1 Pathway in PC12 Cells" Current Issues in Molecular Biology 45, no. 10: 8071-8090. https://doi.org/10.3390/cimb45100510
APA StyleŞirin, S. (2023). Lactic Acid Bacteria-Derived Exopolysaccharides Mitigate the Oxidative Response via the NRF2-KEAP1 Pathway in PC12 Cells. Current Issues in Molecular Biology, 45(10), 8071-8090. https://doi.org/10.3390/cimb45100510