
Astragalus membranaceus and Cinnamomum cassia Stimulate the Hair Follicle Differentiation-Related Growth Factor by the Wnt/β-Catenin Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Network Analysis
2.2. Functional Enrichment Analysis on the GO Biological Process Database
2.3. Sample Preparation
2.4. Animal Experiments
2.5. Morphological and Dermatological Analysis
2.6. Scanning Electron Microscopy (SEM)
2.7. Histology
2.8. Immunohistochemistry
2.9. Reverse Transcription Polymerase Chain Reaction
2.10. Western Blotting Analysis
2.11. Statistical Analysis
3. Results
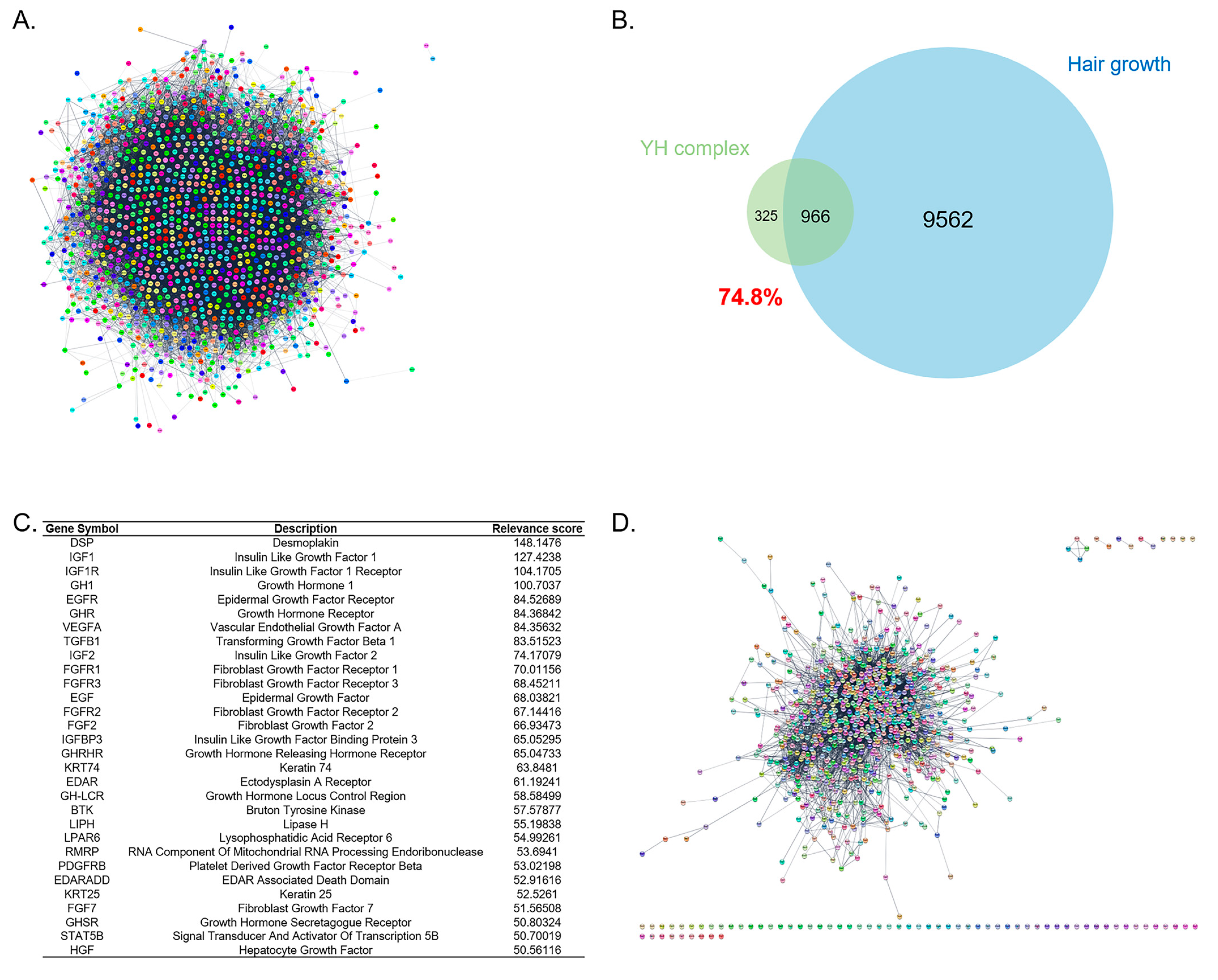
3.1. Investigation of the Association of YH Complex and Hair Growth through Network Construction
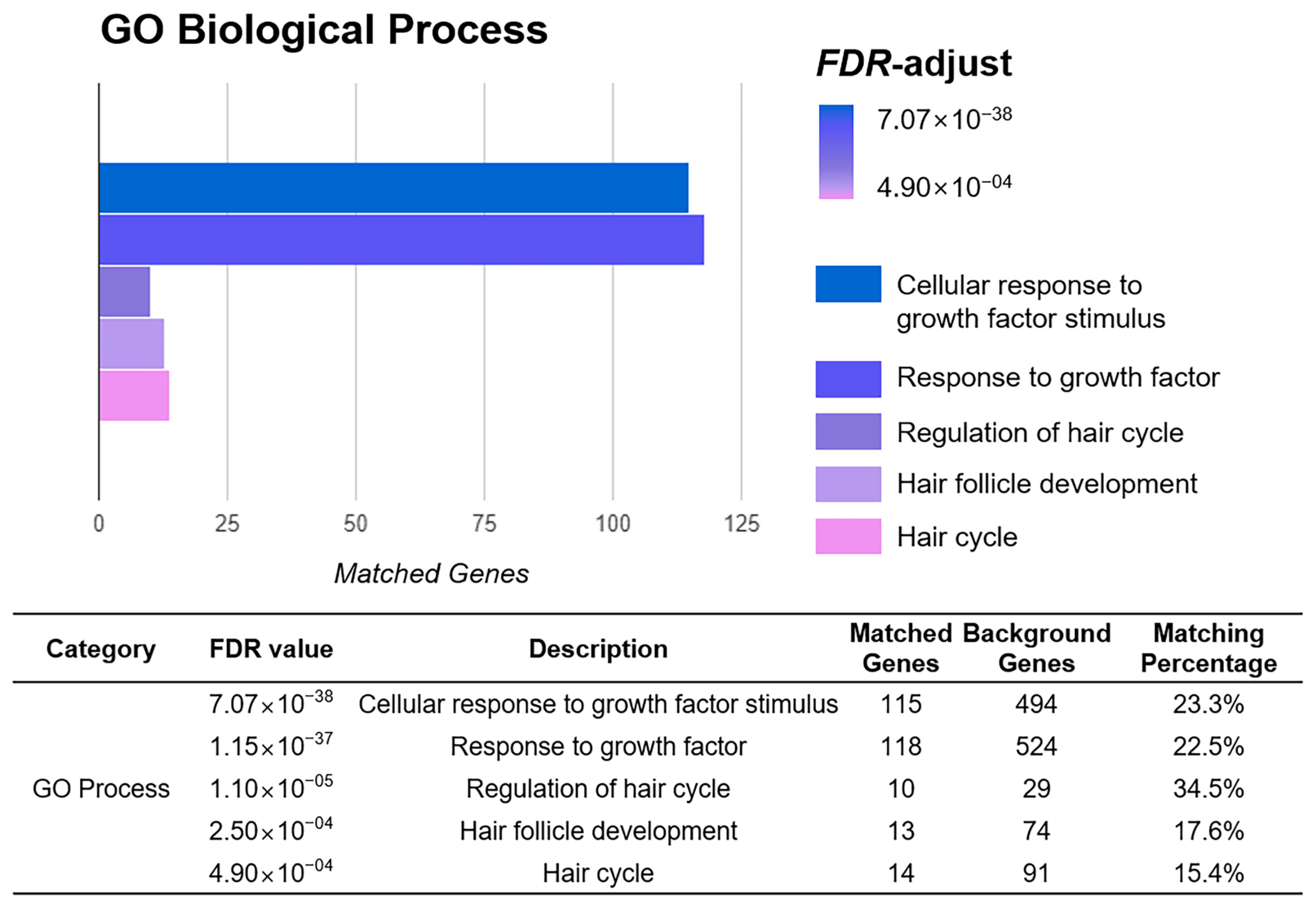
3.2. Prediction of Significant Pathway by Functional Enrichment Analysis of the Target Genes of YH Complex and Hair Growth
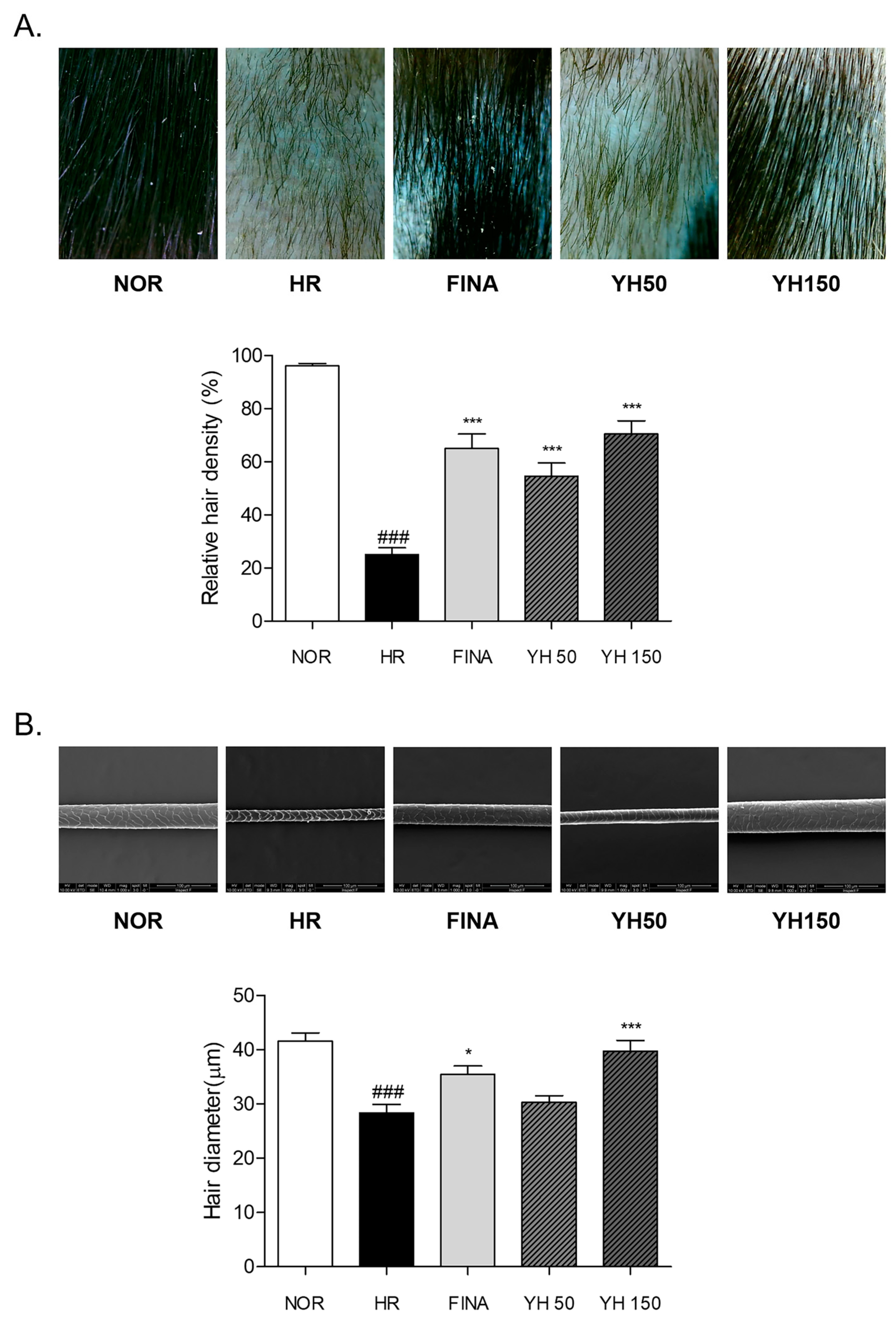
3.3. Effects of YH Complex on Morphological Changes and Hair Density in Hair Removal Model
3.4. Effects of YH Complex on Hair Shaft Morphology
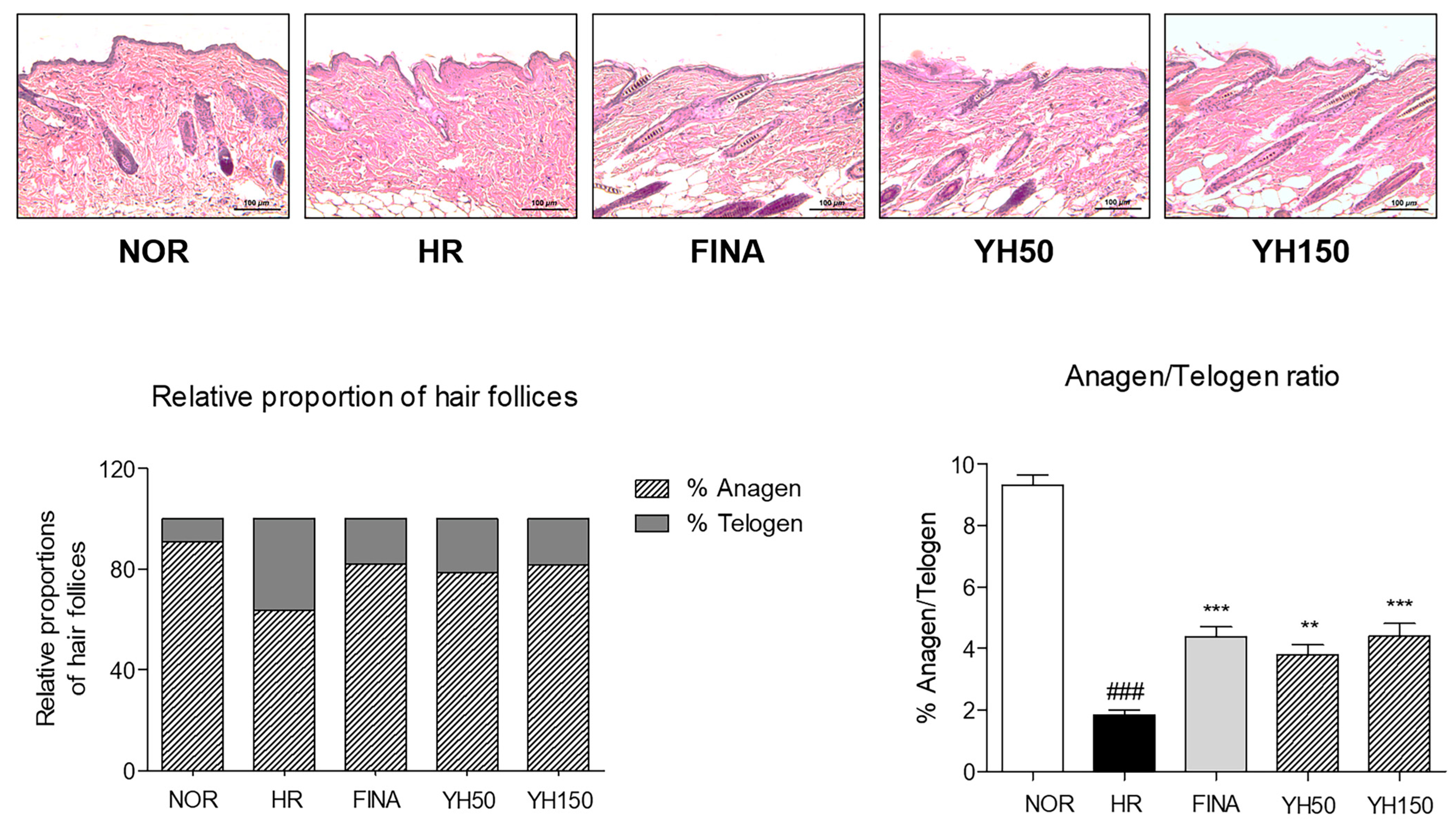
3.5. Effects of YH Complex on Hair Follicles in the Anagen Phase
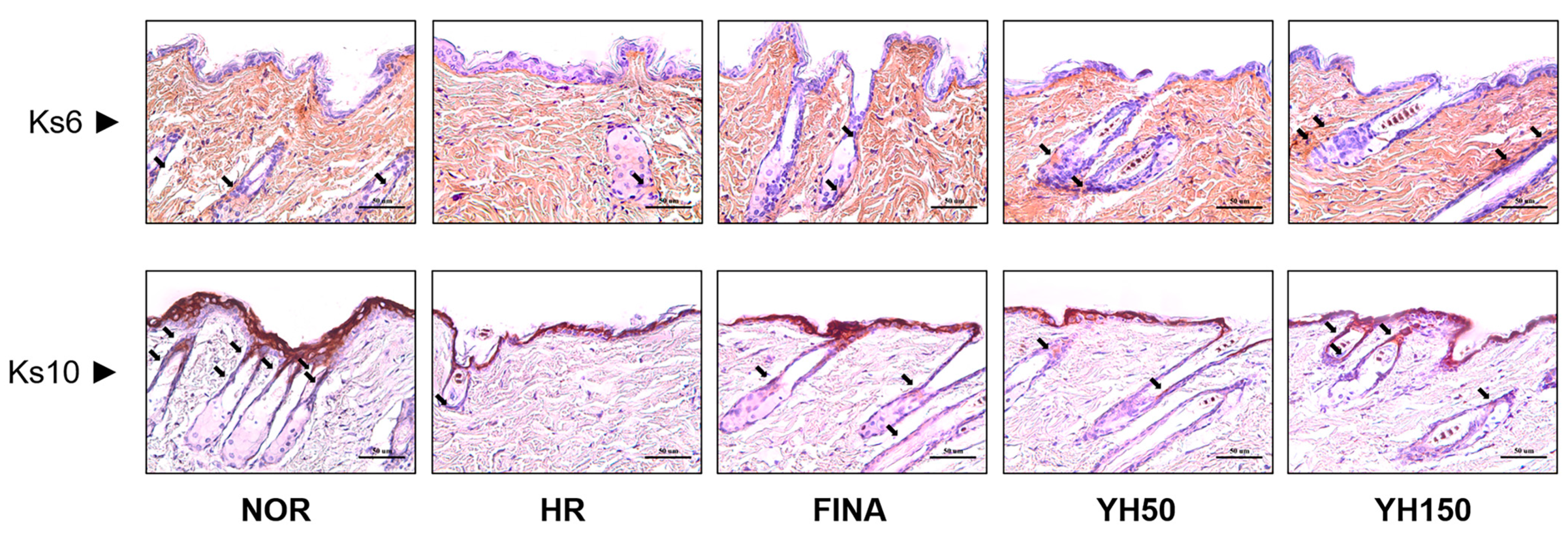
3.6. Effects of YH Complex on Keratinocyte by IHC Analysis
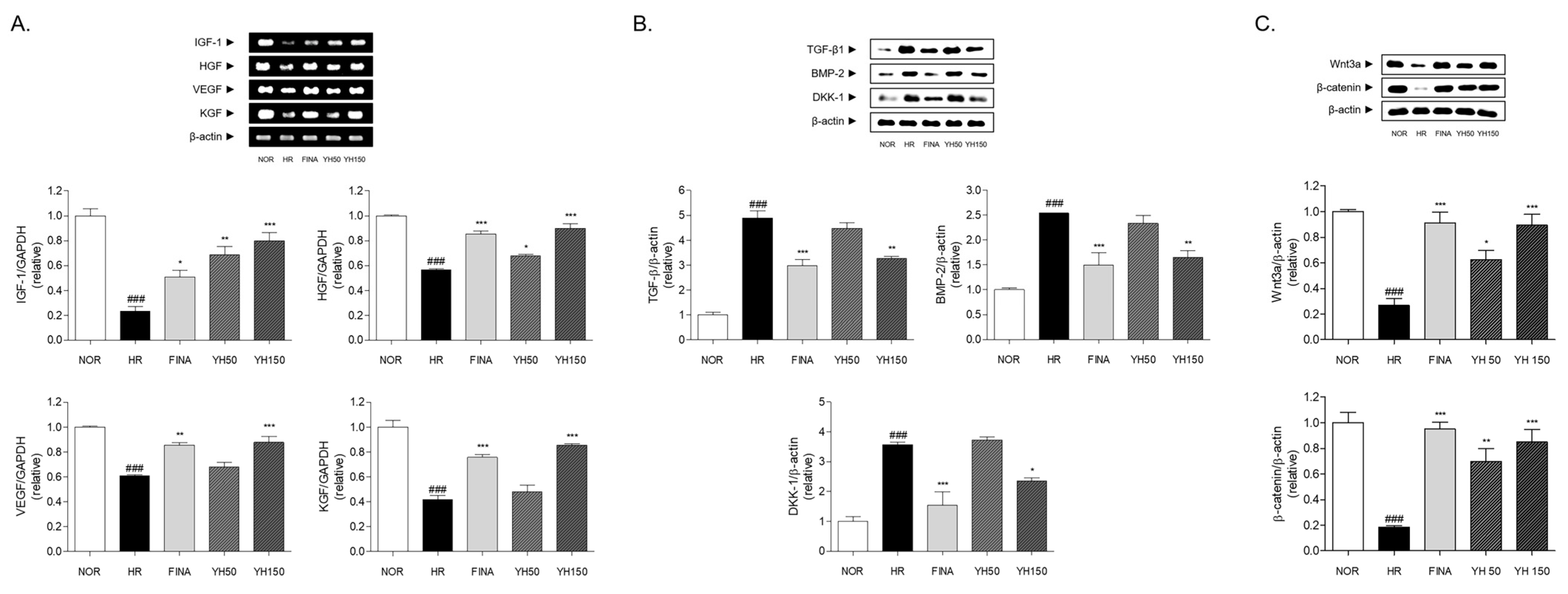
3.7. Effects of YH Complex on Hair Growth Factors in Skin
3.8. Effects of YH Complex on Hair Growth Inhibitor in Skin
3.9. Effects of YH Complex on Wnt/β-Catenin Signaling Pathway in Skin
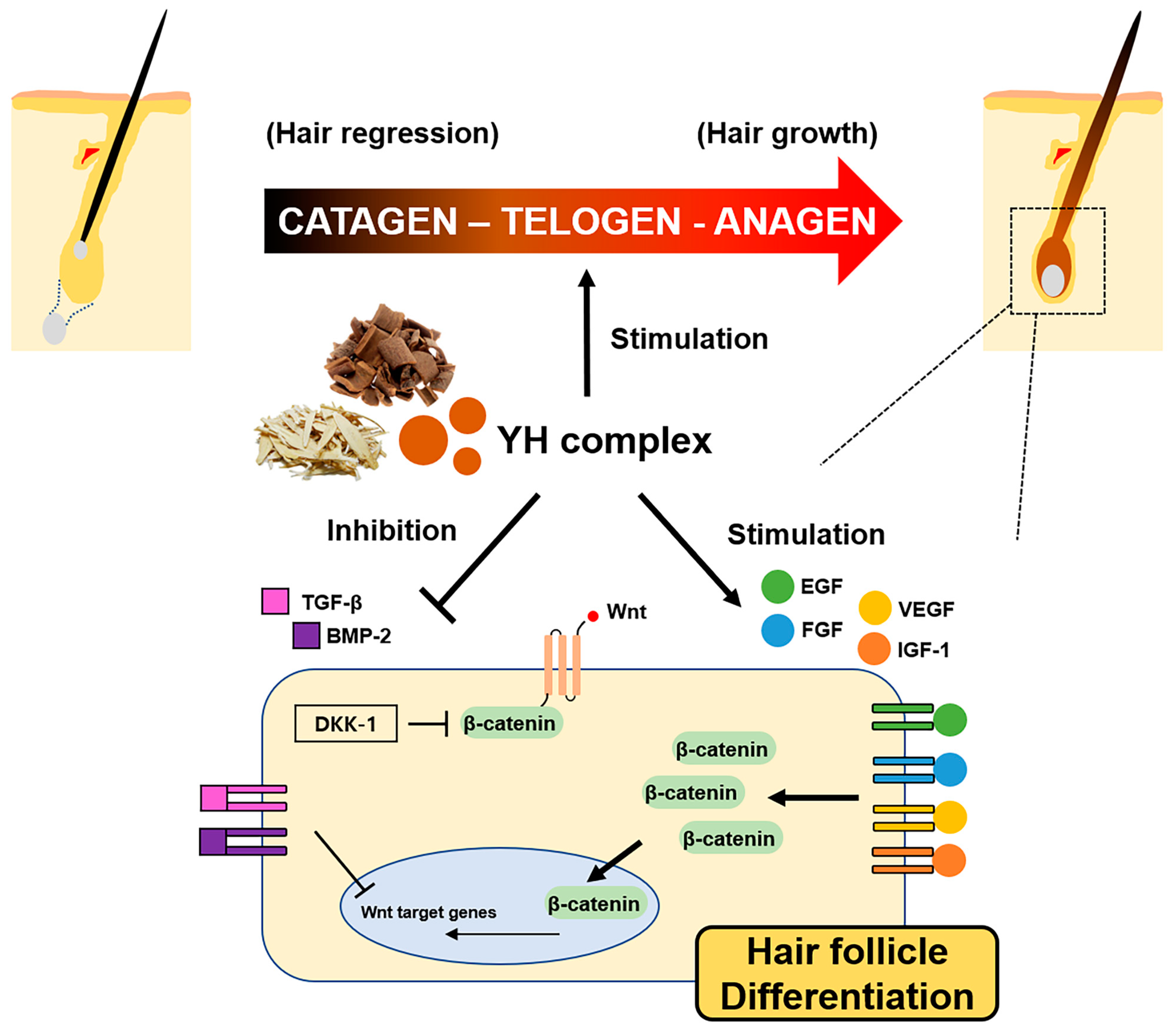
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolff, H.; Fischer, T.W.; Blume-Peytavi, U. The Diagnosis and Treatment of Hair and Scalp Diseases. Dtsch. Arztebl. Int. 2016, 113, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Jaks, V.; Kasper, M.; Toftgard, R. The hair follicle-a stem cell zoo. Exp. Cell Res. 2010, 316, 1422–1428. [Google Scholar] [CrossRef]
- Wall, D.; Meah, N.; Fagan, N.; York, K.; Sinclair, R. Advances in hair growth. Fac. Rev. 2022, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.G.; Slomiany, W.P.; Allison, R. Hair Loss: Common Causes and Treatment. Am. Fam. Physician 2017, 96, 371–378. [Google Scholar]
- Park, S.; Lee, J. Modulation of Hair Growth Promoting Effect by Natural Products. Pharmaceutics 2021, 13, 2163. [Google Scholar] [CrossRef]
- Semalty, M.; Semalty, A.; Joshi, G.P.; Rawat, M.S. Hair growth and rejuvenation: An overview. J. Dermatolog. Treat. 2011, 22, 123–132. [Google Scholar] [CrossRef]
- Krause, K.; Foitzik, K. Biology of the hair follicle: The basics. Semin. Cutan. Med. Surg. 2006, 25, 2–10. [Google Scholar] [CrossRef]
- Johnstone, M.A.; Albert, D.M. Prostaglandin-induced hair growth. Surv. Ophthalmol. 2002, 47 (Suppl. S1), S185–S202. [Google Scholar] [CrossRef]
- Synnott, A. Shame and Glory: A Sociology of Hair. Br. J. Sociol. 1987, 38, 381–413. [Google Scholar] [CrossRef]
- Kranz, D.; Nadarevic, L.; Erdfelder, E. Bald and Bad? Exp. Psychol. 2019, 66, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.; Gonzalez, M.; Finlay, A.Y. The effect of hair loss on quality of life. J. Eur. Acad. Dermatol. Venereol. 2001, 15, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, T.; Endean, N.; Porter, A. Illness perceptions, coping and quality of life in patients with alopecia. Br. J. Dermatol. 2009, 160, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, A.; Kumar, B. Minoxidil vs finasteride in the treatment of men with androgenetic alopecia. Arch. Dermatol. 2003, 139, 1219–1221. [Google Scholar] [CrossRef]
- Olsen, E.A.; Messenger, A.G.; Shapiro, J.; Bergfeld, W.F.; Hordinsky, M.K.; Roberts, J.L.; Stough, D.; Washenik, K.; Whiting, D.A. Evaluation and treatment of male and female pattern hair loss. J. Am. Acad. Dermatol. 2005, 52, 301–311. [Google Scholar] [CrossRef]
- Gordon, K.A.; Tosti, A. Alopecia: Evaluation and treatment. Clin. Cosmet. Investig. Dermatol. 2011, 4, 101–106. [Google Scholar] [CrossRef]
- Kiguradze, T.; Temps, W.H.; Yarnold, P.R.; Cashy, J.; Brannigan, R.E.; Nardone, B.; Micali, G.; West, D.P.; Belknap, S.M. Persistent erectile dysfunction in men exposed to the 5alpha-reductase inhibitors, finasteride, or dutasteride. PeerJ 2017, 5, e3020. [Google Scholar] [CrossRef]
- Ebner, H.; Muller, E. Allergic contact dermatitis from minoxidil. Contact Dermat. 1995, 32, 316–317. [Google Scholar] [CrossRef]
- Panchaprateep, R.; Lueangarun, S. Efficacy and Safety of Oral Minoxidil 5 mg Once Daily in the Treatment of Male Patients with Androgenetic Alopecia: An Open-Label and Global Photographic Assessment. Dermatol. Ther. 2020, 10, 1345–1357. [Google Scholar] [CrossRef]
- Nestor, M.S.; Ablon, G.; Gade, A.; Han, H.; Fischer, D.L. Treatment options for androgenetic alopecia: Efficacy, side effects, compliance, financial considerations, and ethics. J. Cosmet. Dermatol. 2021, 20, 3759–3781. [Google Scholar] [CrossRef]
- Hosking, A.M.; Juhasz, M.; Atanaskova Mesinkovska, N. Complementary and Alternative Treatments for Alopecia: A Comprehensive Review. Skin Appendage Disord. 2019, 5, 72–89. [Google Scholar] [CrossRef]
- Auyeung, K.K.; Han, Q.B.; Ko, J.K. Astragalus membranaceus: A Review of its Protection Against Inflammation and Gastrointestinal Cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 28, 1275–1283. [Google Scholar] [CrossRef]
- Cho, W.C.; Leung, K.N. In vitro and in vivo immunomodulating and immunorestorative effects of Astragalus membranaceus. J. Ethnopharmacol. 2007, 113, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.F.; Aziz, M.; Muhammad, J.S.; Kadowaki, M. Review: Diverse pharmacological properties of Cinnamomum cassia: A review. Pak. J. Pharm. Sci. 2015, 28, 1433–1438. [Google Scholar] [PubMed]
- Wu, H.C.; Horng, C.T.; Lee, Y.L.; Chen, P.N.; Lin, C.Y.; Liao, C.Y.; Hsieh, Y.S.; Chu, S.C. Cinnamomum Cassia Extracts Suppress Human Lung Cancer Cells Invasion by Reducing u-PA/MMP Expression through the FAK to ERK Pathways. Int. J. Med. Sci. 2018, 15, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Hsieh, Y.S.; Chu, S.C.; Hsu, L.S.; Huang, S.C.; Chen, P.N. Reduction of invasion and cell stemness and induction of apoptotic cell death by Cinnamomum cassia extracts on human osteosarcoma cells. Environ. Toxicol. 2022, 37, 1261–1274. [Google Scholar] [CrossRef]
- Kim, M.H.; Bok, M.; Lim, H.; Yang, W.M. An Integrative Study on the Inhibition of Bone Loss via Osteo-F Based on Network Pharmacology, Experimental Verification, and Clinical Trials in Postmenopausal Women. Cells 2023, 12, 1992. [Google Scholar] [CrossRef]
- Ji, S.; Zhu, Z.; Sun, X.; Fu, X. Functional hair follicle regeneration: An updated review. Signal Transduct. Target Ther. 2021, 6, 66. [Google Scholar] [CrossRef]
- Shimomura, Y.; Christiano, A.M. Biology and genetics of hair. Annu. Rev. Genomics Hum. Genet. 2010, 11, 109–132. [Google Scholar] [CrossRef]
- Geyfman, M.; Plikus, M.V.; Treffeisen, E.; Andersen, B.; Paus, R. Resting no more: Re-defining telogen, the maintenance stage of the hair growth cycle. Biol. Rev. Camb. Philos. Soc. 2015, 90, 1179–1196. [Google Scholar] [CrossRef]
- Natarelli, N.; Gahoonia, N.; Sivamani, R.K. Integrative and Mechanistic Approach to the Hair Growth Cycle and Hair Loss. J. Clin. Med. 2023, 12, 893. [Google Scholar] [CrossRef]
- Kimura-Ueki, M.; Oda, Y.; Oki, J.; Komi-Kuramochi, A.; Honda, E.; Asada, M.; Suzuki, M.; Imamura, T. Hair cycle resting phase is regulated by cyclic epithelial FGF18 signaling. J. Investig. Dermatol. 2012, 132, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; He, J.; Wang, J.; Chen, X.; Yang, R. Regulation of signaling pathways in hair follicle stem cells. Burn. Trauma 2022, 10, tkac022. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Wosicka, H.; Cal, K. Targeting to the hair follicles: Current status and potential. J. Dermatol. Sci. 2010, 57, 83–89. [Google Scholar] [CrossRef]
- Kim, H.J.; Rice, D.P.; Kettunen, P.J.; Thesleff, I. FGF-, BMP- and Shh-mediated signalling pathways in the regulation of cranial suture morphogenesis and calvarial bone development. Development 1998, 125, 1241–1251. [Google Scholar] [CrossRef]
- Shin, D.W. The Molecular Mechanism of Natural Products Activating Wnt/beta-Catenin Signaling Pathway for Improving Hair Loss. Life 2022, 12, 1856. [Google Scholar] [CrossRef]
- Su, H.Y.; Hickford, J.G.; Bickerstaffe, R.; Palmer, B.R. Insulin-like growth factor 1 and hair growth. Dermatol. Online J. 1999, 5, 1. [Google Scholar] [CrossRef]
- Stewart, C.E.; Rotwein, P. Growth, differentiation, and survival: Multiple physiological functions for insulin-like growth factors. Physiol. Rev. 1996, 76, 1005–1026. [Google Scholar] [CrossRef]
- Nakamura, T.; Mizuno, S. The discovery of hepatocyte growth factor (HGF) and its significance for cell biology, life sciences and clinical medicine. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 588–610. [Google Scholar] [CrossRef] [PubMed]
- Prugsakij, W.; Numsawat, S.; Netchareonsirisuk, P.; Tengamnuay, P.; De-Eknamkul, W. Mechanistic synergy of hair growth promotion by the Avicennia marina extract and its active constituent (avicequinone C) in dermal papilla cells isolated from androgenic alopecia patients. PLoS ONE 2023, 18, e0284853. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yu, Q.C.; Fuchs, E. Targeting expression of keratinocyte growth factor to keratinocytes elicits striking changes in epithelial differentiation in transgenic mice. EMBO J. 1993, 12, 973–986. [Google Scholar] [CrossRef]
- Choi, B.Y. Targeting Wnt/beta-Catenin Pathway for Developing Therapies for Hair Loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef]
- Collins, C.A.; Kretzschmar, K.; Watt, F.M. Reprogramming adult dermis to a neonatal state through epidermal activation of beta-catenin. Development 2011, 138, 5189–5199. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhu, L.; He, J. Morphogenesis, Growth Cycle and Molecular Regulation of Hair Follicles. Front. Cell Dev. Biol. 2022, 10, 899095. [Google Scholar] [CrossRef]
- Kulessa, H.; Turk, G.; Hogan, B.L. Inhibition of Bmp signaling affects growth and differentiation in the anagen hair follicle. EMBO J. 2000, 19, 6664–6674. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.H.; Jin, S.C.; Baek, H.K.; Yang, W.M. Astragalus membranaceus and Cinnamomum cassia Stimulate the Hair Follicle Differentiation-Related Growth Factor by the Wnt/β-Catenin Signaling Pathway. Curr. Issues Mol. Biol. 2023, 45, 8607-8621. https://doi.org/10.3390/cimb45110541
Kim MH, Jin SC, Baek HK, Yang WM. Astragalus membranaceus and Cinnamomum cassia Stimulate the Hair Follicle Differentiation-Related Growth Factor by the Wnt/β-Catenin Signaling Pathway. Current Issues in Molecular Biology. 2023; 45(11):8607-8621. https://doi.org/10.3390/cimb45110541
Chicago/Turabian StyleKim, Mi Hye, Seong Chul Jin, Hee Kyung Baek, and Woong Mo Yang. 2023. "Astragalus membranaceus and Cinnamomum cassia Stimulate the Hair Follicle Differentiation-Related Growth Factor by the Wnt/β-Catenin Signaling Pathway" Current Issues in Molecular Biology 45, no. 11: 8607-8621. https://doi.org/10.3390/cimb45110541
APA StyleKim, M. H., Jin, S. C., Baek, H. K., & Yang, W. M. (2023). Astragalus membranaceus and Cinnamomum cassia Stimulate the Hair Follicle Differentiation-Related Growth Factor by the Wnt/β-Catenin Signaling Pathway. Current Issues in Molecular Biology, 45(11), 8607-8621. https://doi.org/10.3390/cimb45110541