
miRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy

 and
and 
Abstract
:1. Introduction
2. Breast Cancer Treatment Strategies Based on Molecular Characteristics
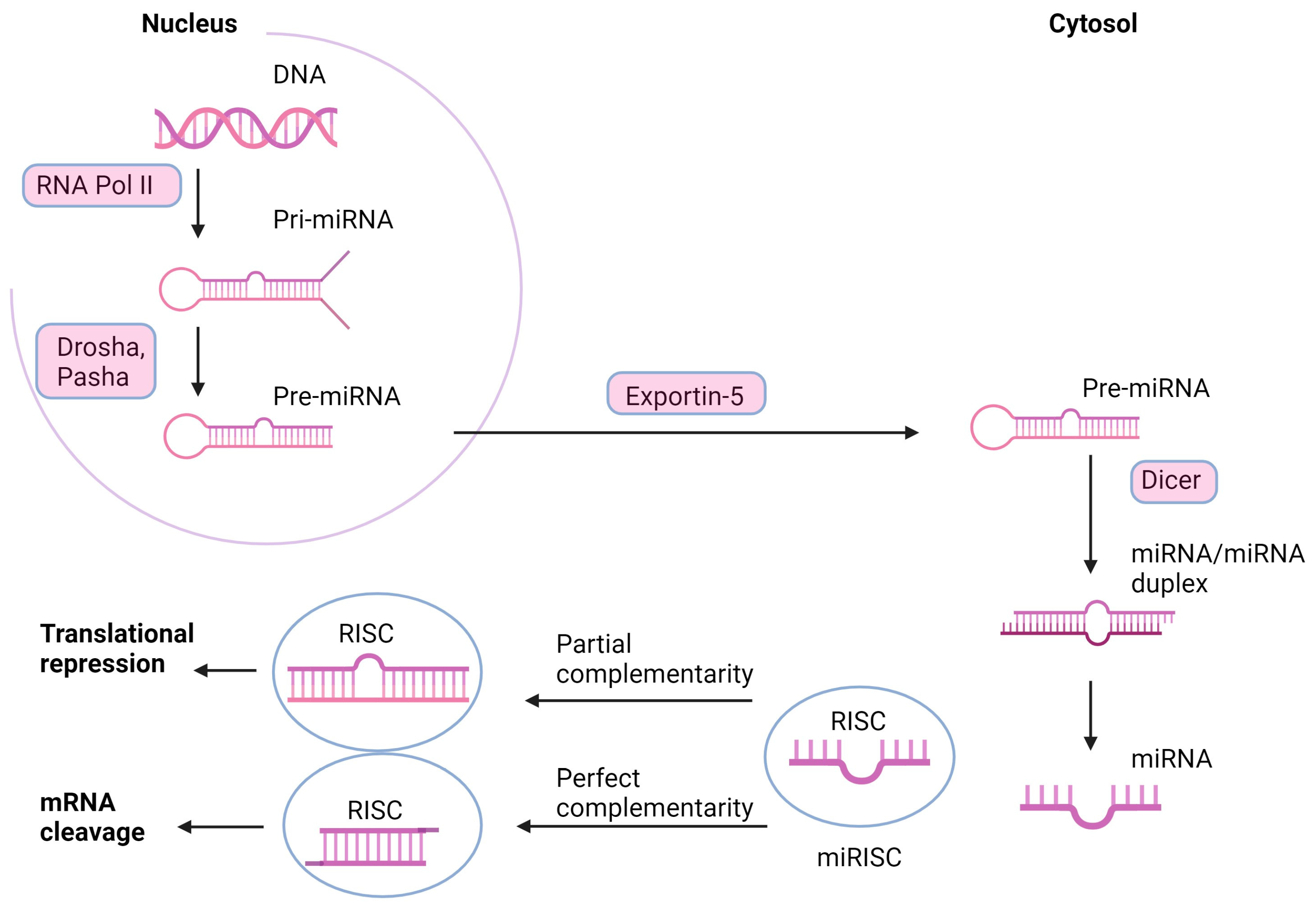
3. miRNAs—The Mechanism of Action
4. The Potential of miRNAs in Current Cancer Diagnostics and Therapy
5. microRNA Profiling
5.1. miR-21
5.2. miR-106a
5.3. miR-155
5.4. miR-141
5.5. Let-7c miRNA
5.6. miR-335
5.7. miR-126
5.8. miR-199a
5.9. miR-101
5.10. miR-9
6. miRNA as a Therapy Target
6.1. The Role of miRNA in Breast Cancer Chemoresistance
6.2. The Role of miRNA in Breast Cancer Stem Cells
6.3. The Role of miRNA in Cancer Cell Cycle Control
6.4. miRNAs and Cell Death
7. Tools in miRNA-Based Therapy Adjustment
8. Conclusions: Challenges in miRNA Modulation Approach
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABCG2 | adenosine triphosphate binding cassette subfamily G member 2 |
| ADAM9 | disintegrin and metalloproteinase domain-containing protein 9 |
| ADC | antibody-drug conjugate |
| Akt | protein kinase B |
| AMOs | anti-messenger ribonucleic acid oligonucleotides |
| ANP32E | acidic nuclear phosphoprotein 32 family member E |
| ATG12 | autophagy-related gene 12 |
| ATM | ataxia telangiectasia mutated kinase |
| BAK1 | Bcl-2 homologous antagonist killer 1 |
| BAX | Bcl-2 associated X protein, apoptosis regulator |
| BC | breast cancer |
| Bcl-2 | B-cell lymphoma 2 |
| BCSCs | breast cancer stem cells |
| BNIP3 | BCL2 interacting protein 3 |
| BRCA1/2 | breast cancer gene 1/2 |
| CCND1 | cyclin D1 |
| CDKs | cyclin-dependent kinases |
| cDNA | complementary deoxyribonucleic acid |
| CIS | cisplatin |
| COX-2 | cyclooxygenase 2 |
| CSCs | cancer stem cells |
| Dicer | endoribonuclease Dicer |
| DNA | deoxyribonucleic acid |
| DNMT3A | deoxyribonucleic acid methyltransferase 3 alpha |
| DOX | doxorubicin |
| DTX | docetaxel |
| c-Met | mesenchymal-epithelial transition factor |
| EGFL7 | epidermal growth factor-like protein 7 |
| EMT | epithelial-to-mesenchymal transition |
| ER | estrogen receptor |
| ERBB2 | erythroblastic oncogene B2 |
| ERCC6 | deoxyribonucleic acid excision repair protein |
| ERKs | extracellular signal-regulated kinases |
| FDA | Food and Drug Administration |
| FOXO1 | forkhead box protein O1 |
| GDP | guanosine diphosphate |
| GEFs | guanine–nucleotide exchange factors |
| GTP | guanosine triphosphate |
| HAX-1 | HCLS1-associated protein X-1 |
| HER2 | human epidermal growth factor receptor 2 |
| HGF | hepatocyte growth factor |
| IGFBP3 | insulin-like growth factor binding protein 3 |
| JAK/STAT | janus kinase/signal transducer and activator of transcription |
| MAPKs | mitogen-activated protein kinases |
| MDR | multidrug resistance |
| MMP1 | matrix metallopeptidase 1 |
| mRNA | messenger ribonucleic acid |
| miRNA | micro-ribonucleic acid |
| mTOR | mammalian target of rapamycin |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NVB | vinorelbine |
| P53 | tumor protein p53 |
| PAB | palbociclib |
| PARP | poly (ADP-ribose) polymerase |
| PAK4 | serine/threonine-protein kinase |
| PDCD4 | programmed cell death protein 4 |
| PI3K | phosphoinositide 3-kinase |
| PIK3R2 | phosphoinositide 3-kinase regulatory subunit 2 |
| Pol II | ribonucleic acid polymerase II |
| POMP | proteasome maturation protein |
| PR | progesterone receptor |
| pri-miRNA | primary micro-ribonucleic acid |
| pre-miRNA | precursor micro-ribonucleic acid |
| PTEN | phosphatase and tensin homolog |
| PTX | paclitaxel |
| RAF | rapidly accelerated fibrosarcoma |
| RISC | ribonucleic acid-induced silencing complex |
| RNA | ribonucleic acid |
| RNase | ribonuclease |
| RTKs | receptor tyrosine kinases |
| RUNX3 | runt-related transcription factor 3 |
| ssRNAs | single-stranded ribonucleic acids |
| STAT3 | signal transducer and activator of transcription 3 |
| TFAM | transcription factor A, mitochondrial |
| TGF-β | transforming growth factor-beta |
| TKI | tyrosine kinase inhibitors |
| TNM | tumor: node, metastasis |
| TPM1 | tropomyosin 1 |
| VEGF | vascular endothelial growth factor |
| WHO | World Health Organization |
| RT-qPCR | quantitative reverse transcription polymerase chain reaction |
| 3′UTR | three prime untranslated region |
| 5′UTR | five prime untranslated region |
References
- Akram, M.; Iqbal, M.; Daniyal, M.; Khan, A.U. Awareness and Current Knowledge of Breast Cancer. Biol. Res. 2017, 50, 33. [Google Scholar] [CrossRef] [PubMed]
- Macias, H.; Hinck, L. Mammary Gland Development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [PubMed]
- Yao, N.; Shi, W.; Liu, T.; Siyin, S.T.; Wang, W.; Duan, N.; Xu, G.; Qu, J. Clinicopathologic Characteristics and Prognosis for Male Breast Cancer Compared to Female Breast Cancer. Sci. Rep. 2022, 12, 220. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L.; Gathani, T. Understanding Breast Cancer as a Global Health Concern. Br. J. Radiol. 2022, 95, 20211033. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Krajewska-Kułak, E. Rola Zespołu Interdyscyplinarnego w Opiece nad Pacjentami Onkologicznymi. T. 1; Uniwersytet Medyczny w Białymstoku: Białystok, Poland, 2021. [Google Scholar]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Komuravelli, M.; Gaddala, A.; Nama, S.; Matli, P.; Anchuri, K. Awareness and Attitude Regarding the Risk Factors, and Self Examination for Breast Cancer in the Students of an Engineering College of South India. Eur. J. Mol. Clin. Med. 2022, 9. [Google Scholar]
- Maughan, K.L.; Lutterbie, M.A.; Ham, P.S. Treatment of Breast Cancer. Am. Fam. Physician 2010, 81, 1339–1346. [Google Scholar]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef]
- Advances in Breast Cancer Research—NCI. Available online: https://www.cancer.gov/types/breast/research (accessed on 26 September 2023).
- Sher, G.; Salman, N.A.; Khan, A.Q.; Prabhu, K.S.; Raza, A.; Kulinski, M.; Dermime, S.; Haris, M.; Junejo, K.; Uddin, S. Epigenetic and Breast Cancer Therapy: Promising Diagnostic and Therapeutic Applications. Semin. Cancer Biol. 2022, 83, 152–165. [Google Scholar] [CrossRef]
- Hagemann, I.S. Molecular Testing in Breast Cancer: A Guide to Current Practices. Arch. Pathol. Lab. Med. 2016, 140, 815–824. [Google Scholar] [CrossRef]
- Breast Cancer Hormone Receptor Status|Estrogen Receptor. Available online: https://www.cancer.org/cancer/types/breast-cancer/understanding-a-breast-cancer-diagnosis/breast-cancer-hormone-receptor-status.html (accessed on 26 September 2023).
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef] [PubMed]
- Targeted Drug Therapy|Breast Cancer Treatment. Available online: https://www.cancer.org/cancer/types/breast-cancer/treatment/targeted-therapy-for-breast-cancer.html (accessed on 26 September 2023).
- Leivonen, S.-K.; Sahlberg, K.K.; Mäkelä, R.; Due, E.U.; Kallioniemi, O.; Børresen-Dale, A.-L.; Perälä, M. High-Throughput Screens Identify microRNAs Essential for HER2 Positive Breast Cancer Cell Growth. Mol. Oncol. 2014, 8, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Uras, I.Z.; Sexl, V.; Kollmann, K. CDK6 Inhibition: A Novel Approach in AML Management. Int. J. Mol. Sci. 2020, 21, 2528. [Google Scholar] [CrossRef]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. mTOR Signaling Pathway and mTOR Inhibitors in Cancer: Progress and Challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Varol, U.; Kucukzeybek, Y.; Alacacioglu, A.; Somali, I.; Altun, Z.; Aktas, S.; Oktay Tarhan, M. BRCA Genes: BRCA 1 and BRCA 2. J. BUON Off. J. Balk. Union Oncol. 2018, 23, 862–866. [Google Scholar]
- Chang, S.; Sharan, S.K. BRCA1 and MicroRNAs: Emerging Networks and Potential Therapeutic Targets. Mol. Cells 2012, 34, 425–432. [Google Scholar] [CrossRef]
- Chang, S.; Wang, R.-H.; Akagi, K.; Kim, K.-A.; Martin, B.K.; Cavallone, L.; Haines, D.C.; Basik, M.; Mai, P.; Poggi, E.; et al. Tumor Suppressor BRCA1 Epigenetically Controls Oncogenic microRNA-155. Nat. Med. 2011, 17, 1275–1282. [Google Scholar] [CrossRef]
- Kawai, S.; Amano, A. BRCA1 Regulates microRNA Biogenesis via the DROSHA Microprocessor Complex. J. Cell Biol. 2012, 197, 201–208. [Google Scholar] [CrossRef]
- SEER. SEER Cancer Statistics Review, 1975–2017. Available online: https://seer.cancer.gov/csr/1975_2017/index.html (accessed on 26 September 2023).
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A Uniform System for microRNA Annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef]
- Budzyński, M.; Grenda, A.; Filip, A.A. Cząsteczki mikroRNA Jako Istotny Składnik Mechanizmów Regulacji Ekspresji Genów Związanych z Nowotworami. Nowotw. J. Oncol. 2014, 64, 48. [Google Scholar] [CrossRef]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. Bantam Encodes a Developmentally Regulated microRNA That Controls Cell Proliferation and Regulates the Proapoptotic Gene Hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef] [PubMed]
- PLoS Biology. Identification of Drosophila MicroRNA Targets. Available online: https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0000060 (accessed on 26 September 2023).
- Lai, E.C.; Tam, B.; Rubin, G.M. Pervasive Regulation of Drosophila Notch Target Genes by GY-Box-, Brd-Box-, and K-Box-Class microRNAs. Genes Dev. 2005, 19, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Thai, T.-H.; Calado, D.P.; Casola, S.; Ansel, K.M.; Xiao, C.; Xue, Y.; Murphy, A.; Frendewey, D.; Valenzuela, D.; Kutok, J.L.; et al. Regulation of the Germinal Center Response by microRNA-155. Science 2007, 316, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Vigorito, E.; Perks, K.L.; Abreu-Goodger, C.; Bunting, S.; Xiang, Z.; Kohlhaas, S.; Das, P.P.; Miska, E.A.; Rodriguez, A.; Bradley, A.; et al. microRNA-155 Regulates the Generation of Immunoglobulin Class-Switched Plasma Cells. Immunity 2007, 27, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of Bic/microRNA-155 for Normal Immune Function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef]
- Fazi, F.; Rosa, A.; Fatica, A.; Gelmetti, V.; De Marchis, M.L.; Nervi, C.; Bozzoni, I. A Minicircuitry Comprised of microRNA-223 and Transcription Factors NFI-A and C/EBPalpha Regulates Human Granulopoiesis. Cell 2005, 123, 819–831. [Google Scholar] [CrossRef]
- Chang, T.-C.; Yu, D.; Lee, Y.-S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microRNA Repression by Myc Contributes to Tumorigenesis. Nat. Genet. 2008, 40, 43–50. [Google Scholar] [CrossRef]
- Esau, C.; Kang, X.; Peralta, E.; Hanson, E.; Marcusson, E.G.; Ravichandran, L.V.; Sun, Y.; Koo, S.; Perera, R.J.; Jain, R.; et al. MicroRNA-143 Regulates Adipocyte Differentiation. J. Biol. Chem. 2004, 279, 52361–52365. [Google Scholar] [CrossRef]
- Lecellier, C.-H.; Dunoyer, P.; Arar, K.; Lehmann-Che, J.; Eyquem, S.; Himber, C.; Saïb, A.; Voinnet, O. A Cellular microRNA Mediates Antiviral Defense in Human Cells. Science 2005, 308, 557–560. [Google Scholar] [CrossRef]
- Michael, M.Z.; O’ Connor, S.M.; van Holst Pellekaan, N.G.; Young, G.P.; James, R.J. Reduced Accumulation of Specific microRNAs in Colorectal Neoplasia. Mol. Cancer Res. MCR 2003, 1, 882–891. [Google Scholar] [PubMed]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J.; et al. A microRNA Polycistron as a Potential Human Oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. C-Myc-Regulated microRNAs Modulate E2F1 Expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Mizielska, A.; Dziechciowska, I.; Szczepański, R.; Cisek, M.; Dąbrowska, M.; Ślężak, J.; Kosmalska, I.; Rymarczyk, M.; Wilkowska, K.; Jacczak, B.; et al. Doxorubicin and Cisplatin Modulate miR-21, miR-106, miR-126, miR-155 and miR-199 Levels in MCF7, MDA-MB-231 and SK-BR-3 Cells That Makes Them Potential Elements of the DNA-Damaging Drug Treatment Response Monitoring in Breast Cancer Cells-A Preliminary Study. Genes 2023, 14, 702. [Google Scholar] [CrossRef]
- Ye, Q.; Raese, R.A.; Luo, D.; Feng, J.; Xin, W.; Dong, C.; Qian, Y.; Guo, N.L. MicroRNA-Based Discovery of Biomarkers, Therapeutic Targets, and Repositioning Drugs for Breast Cancer. Cells 2023, 12, 1917. [Google Scholar] [CrossRef]
- Ahmadi, S.M.; Amirkhanloo, S.; Yazdian-Robati, R.; Ebrahimi, H.; Pirhayati, F.H.; Almalki, W.H.; Ebrahimnejad, P.; Kesharwani, P. Recent Advances in Novel miRNA Mediated Approaches for Targeting Breast Cancer. J. Drug Target. 2023, 31, 777–793. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Fang, H.; Xie, J.; Zhang, M.; Zhao, Z.; Wan, Y.; Yao, Y. miRNA-21 Promotes Proliferation and Invasion of Triple-Negative Breast Cancer Cells through Targeting PTEN. Am. J. Transl. Res. 2017, 9, 953–961. [Google Scholar]
- Magri, F.; Vanoli, F.; Corti, S. miRNA in Spinal Muscular Atrophy Pathogenesis and Therapy. J. Cell. Mol. Med. 2018, 22, 755–767. [Google Scholar] [CrossRef]
- Singh, R.; Mo, Y.-Y. Role of microRNAs in Breast Cancer. Cancer Biol. Ther. 2013, 14, 201–212. [Google Scholar] [CrossRef]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Hamam, R.; Hamam, D.; Alsaleh, K.A.; Kassem, M.; Zaher, W.; Alfayez, M.; Aldahmash, A.; Alajez, N.M. Circulating microRNAs in Breast Cancer: Novel Diagnostic and Prognostic Biomarkers. Cell Death Dis. 2017, 8, e3045. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Abolghasemi, M.; Tehrani, S.S.; Yousefi, T.; Karimian, A.; Mahmoodpoor, A.; Ghamari, A.; Jadidi-Niaragh, F.; Yousefi, M.; Kafil, H.S.; Bastami, M.; et al. MicroRNAs in Breast Cancer: Roles, Functions, and Mechanism of Actions. J. Cell. Physiol. 2020, 235, 5008–5029. [Google Scholar] [CrossRef] [PubMed]
- Safa, A.; Abak, A.; Shoorei, H.; Taheri, M.; Ghafouri-Fard, S. MicroRNAs as Regulators of ERK/MAPK Pathway: A Comprehensive Review. Biomed. Pharmacother. 2020, 132, 110853. [Google Scholar] [CrossRef] [PubMed]
- Sudhesh Dev, S.; Zainal Abidin, S.A.; Farghadani, R.; Othman, I.; Naidu, R. Receptor Tyrosine Kinases and Their Signaling Pathways as Therapeutic Targets of Curcumin in Cancer. Front. Pharmacol. 2021, 12, 772510. [Google Scholar] [CrossRef] [PubMed]
- McCain, J. The MAPK (ERK) Pathway: Investigational Combinations for the Treatment of BRAF-Mutated Metastatic Melanoma. P T Peer-Rev. J. Formul. Manag. 2013, 38, 96–108. [Google Scholar]
- Xuhong, J.-C.; Qi, X.-W.; Zhang, Y.; Jiang, J. Mechanism, Safety and Efficacy of Three Tyrosine Kinase Inhibitors Lapatinib, Neratinib and Pyrotinib in HER2-Positive Breast Cancer. Am. J. Cancer Res. 2019, 9, 2103–2119. [Google Scholar]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA Profiling: Approaches and Considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Real-Time qRT-PCR. Available online: https://www.ncbi.nlm.nih.gov/probe/docs/techqpcr/ (accessed on 26 September 2023).
- Benes, V.; Castoldi, M. Expression Profiling of microRNA Using Real-Time Quantitative PCR, How to Use It and What Is Available. Methods 2010, 50, 244–249. [Google Scholar] [CrossRef]
- Najjary, S.; Mohammadzadeh, R.; Mokhtarzadeh, A.; Mohammadi, A.; Kojabad, A.B.; Baradaran, B. Role of miR-21 as an Authentic Oncogene in Mediating Drug Resistance in Breast Cancer. Gene 2020, 738, 144453. [Google Scholar] [CrossRef] [PubMed]
- TargetScanHuman 8.0. Available online: https://www.targetscan.org/vert_80/ (accessed on 26 September 2023).
- Si, H.; Sun, X.; Chen, Y.; Cao, Y.; Chen, S.; Wang, H.; Hu, C. Circulating microRNA-92a and microRNA-21 as Novel Minimally Invasive Biomarkers for Primary Breast Cancer. J. Cancer Res. Clin. Oncol. 2013, 139, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Lu, B.-B.; Wang, H.; Cheng, Z.-X.; Yin, Y.-M. MicroRNA-21 Modulates Chemosensitivity of Breast Cancer Cells to Doxorubicin by Targeting PTEN. Arch. Med. Res. 2011, 42, 281–290. [Google Scholar] [CrossRef]
- Gong, C.; Yao, Y.; Wang, Y.; Liu, B.; Wu, W.; Chen, J.; Su, F.; Yao, H.; Song, E. Up-Regulation of miR-21 Mediates Resistance to Trastuzumab Therapy for Breast Cancer. J. Biol. Chem. 2011, 286, 19127–19137. [Google Scholar] [CrossRef]
- Wu, Z.-H.; Tao, Z.-H.; Zhang, J.; Li, T.; Ni, C.; Xie, J.; Zhang, J.-F.; Hu, X.-C. MiRNA-21 Induces Epithelial to Mesenchymal Transition and Gemcitabine Resistance via the PTEN/AKT Pathway in Breast Cancer. Tumor Biol. 2016, 37, 7245–7254. [Google Scholar] [CrossRef]
- Shi, Y.; Ye, P.; Long, X. Differential Expression Profiles of the Transcriptome in Breast Cancer Cell Lines Revealed by Next Generation Sequencing. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Si, M.-L.; Wu, H.; Mo, Y.-Y. MicroRNA-21 Targets the Tumor Suppressor Gene Tropomyosin 1 (TPM1). J. Biol. Chem. 2007, 282, 14328–14336. [Google Scholar] [CrossRef]
- Zhu, S.; Wu, H.; Wu, F.; Nie, D.; Sheng, S.; Mo, Y.-Y. MicroRNA-21 Targets Tumor Suppressor Genes in Invasion and Metastasis. Cell Res. 2008, 18, 350–359. [Google Scholar] [CrossRef]
- Han, M.; Liu, M.; Wang, Y.; Mo, Z.; Bi, X.; Liu, Z.; Fan, Y.; Chen, X.; Wu, C. Re-Expression of miR-21 Contributes to Migration and Invasion by Inducing Epithelial-Mesenchymal Transition Consistent with Cancer Stem Cell Characteristics in MCF-7 Cells. Mol. Cell Biochem. 2012, 363, 427–436. [Google Scholar] [CrossRef]
- Ahmed Mohmmed, E.; Shousha, W.G.; El-Saiid, A.S.; Ramadan, S.S. A Clinical Evaluation of Circulating MiR-106a and Raf-1 as Breast Cancer Diagnostic and Prognostic Markers. Asian Pac. J. Cancer Prev. APJCP 2021, 22, 3513–3520. [Google Scholar] [CrossRef]
- You, F.; Luan, H.; Sun, D.; Cui, T.; Ding, P.; Tang, H.; Sun, D. miRNA-106a Promotes Breast Cancer Cell Proliferation, Clonogenicity, Migration, and Invasion Through Inhibiting Apoptosis and Chemosensitivity. DNA Cell Biol. 2019, 38, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Yan, L.; Qi, Q.; Peng, X.; Edge, S.B.; Young, J.; Yao, S.; Liu, S.; Otsuji, E.; Takabe, K. Novel MicroRNA-Based Risk Score Identified by Integrated Analyses to Predict Metastasis and Poor Prognosis in Breast Cancer. Ann. Surg. Oncol. 2018, 25, 4037–4046. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Chadalapaka, G.; Lee, S.-O.; Yamada, D.; Sastre-Garau, X.; Defossez, P.-A.; Park, Y.-Y.; Lee, J.-S.; Safe, S. Identification of Oncogenic microRNA-17-92/ZBTB4/Specificity Protein Axis in Breast Cancer. Oncogene 2012, 31, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Chadalapaka, G.; Cho, S.-G.; Lee, S.; Jin, U.-H.; Jutooru, I.; Choi, K.; Leung, Y.-K.; Ho, S.-M.; Safe, S.; et al. The Transcriptional Repressor ZBTB4 Regulates EZH2 through a MicroRNA-ZBTB4-Specificity Protein Signaling Axis. Neoplasia 2014, 16, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.; Rack, B.; Müller, V.; Janni, W.; Pantel, K.; Schwarzenbach, H. Circulating microRNAs as Blood-Based Markers for Patients with Primary and Metastatic Breast Cancer. Breast Cancer Res. BCR 2010, 12, R90. [Google Scholar] [CrossRef] [PubMed]
- Dinami, R.; Ercolani, C.; Petti, E.; Piazza, S.; Ciani, Y.; Sestito, R.; Sacconi, A.; Biagioni, F.; le Sage, C.; Agami, R.; et al. miR-155 Drives Telomere Fragility in Human Breast Cancer by Targeting TRF1. Cancer Res. 2014, 74, 4145–4156. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 Functions as an OncomiR in Breast Cancer by Targeting the Suppressor of Cytokine Signaling 1 Gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef]
- Ding, L.; Gu, H.; Xiong, X.; Ao, H.; Cao, J.; Lin, W.; Yu, M.; Lin, J.; Cui, Q. MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer. Cells 2019, 8, 1492. [Google Scholar] [CrossRef]
- Johansson, J.; Berg, T.; Kurzejamska, E.; Pang, M.-F.; Tabor, V.; Jansson, M.; Roswall, P.; Pietras, K.; Sund, M.; Religa, P.; et al. MiR-155-Mediated Loss of C/EBPβ Shifts the TGF-β Response from Growth Inhibition to Epithelial-Mesenchymal Transition, Invasion and Metastasis in Breast Cancer. Oncogene 2013, 32, 5614–5624. [Google Scholar] [CrossRef]
- Li, P.; Xu, T.; Zhou, X.; Liao, L.; Pang, G.; Luo, W.; Han, L.; Zhang, J.; Luo, X.; Xie, X.; et al. Downregulation of miRNA-141 in Breast Cancer Cells Is Associated with Cell Migration and Invasion: Involvement of ANP32E Targeting. Cancer Med. 2017, 6, 662–672. [Google Scholar] [CrossRef]
- Xiong, Z.; Ye, L.; Zhenyu, H.; Li, F.; Xiong, Y.; Lin, C.; Wu, X.; Deng, G.; Shi, W.; Song, L.; et al. ANP32E Induces Tumorigenesis of Triple-negative Breast Cancer Cells by Upregulating E2F1. Mol. Oncol. 2018, 12, 896–912. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 Family and miR-205 Regulate Epithelial to Mesenchymal Transition by Targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-X.; Gao, S.-Y.; Wang, P.-Y.; Zhou, X.; Li, Y.-J.; Yu, Y.; Yan, Y.-F.; Zhang, H.-H.; Lv, C.-J.; Zhou, H.-H.; et al. Reduced Expression Levels of Let-7c in Human Breast Cancer Patients. Oncol. Lett. 2015, 9, 1207–1212. [Google Scholar] [CrossRef]
- Fu, X.; Mao, X.; Wang, Y.; Ding, X.; Li, Y. Let-7c-5p Inhibits Cell Proliferation and Induces Cell Apoptosis by Targeting ERCC6 in Breast Cancer. Oncol. Rep. 2017, 38, 1851–1856. [Google Scholar] [CrossRef]
- Bozgeyik, E. Bioinformatic Analysis and In Vitro Validation of Let-7b and Let-7c in Breast Cancer. Comput. Biol. Chem. 2020, 84, 107191. [Google Scholar] [CrossRef]
- Sun, X.; Xu, C.; Tang, S.-C.; Wang, J.; Wang, H.; Wang, P.; Du, N.; Qin, S.; Li, G.; Xu, S.; et al. Let-7c Blocks Estrogen-Activated Wnt Signaling in Induction of Self-Renewal of Breast Cancer Stem Cells. Cancer Gene Ther. 2016, 23, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Swellam, M.; Mahmoud, M.S.; Hashim, M.; Hassan, N.M.; Sobeih, M.E.; Nageeb, A.M. Clinical Aspects of Circulating miRNA-335 in Breast Cancer Patients: A Prospective Study. J. Cell. Biochem. 2019, 120, 8975–8982. [Google Scholar] [CrossRef]
- Heyn, H.; Engelmann, M.; Schreek, S.; Ahrens, P.; Lehmann, U.; Kreipe, H.; Schlegelberger, B.; Beger, C. MicroRNA miR-335 Is Crucial for the BRCA1 Regulatory Cascade in Breast Cancer Development. Int. J. Cancer 2011, 129, 2797–2806. [Google Scholar] [CrossRef]
- Hao, J.; Lai, M.; Liu, C. Expression of miR-335 in Triple-Negative Breast Cancer and Its Effect on Chemosensitivity. J. BUON Off. J. Balk. Union Oncol. 2019, 24, 1526–1531. [Google Scholar]
- Zhu, N.; Zhang, D.; Xie, H.; Zhou, Z.; Chen, H.; Hu, T.; Bai, Y.; Shen, Y.; Yuan, W.; Jing, Q.; et al. Endothelial-Specific Intron-Derived miR-126 Is down-Regulated in Human Breast Cancer and Targets Both VEGFA and PIK3R2. Mol. Cell. Biochem. 2011, 351, 157–164. [Google Scholar] [CrossRef]
- Fu, R.; Tong, J.-S. miR-126 Reduces Trastuzumab Resistance by Targeting PIK3R2 and Regulating AKT/mTOR Pathway in Breast Cancer Cells. J. Cell. Mol. Med. 2020, 24, 7600–7608. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Yuan, P.; Li, Y. MiR-126 Regulated Breast Cancer Cell Invasion by Targeting ADAM9. Int. J. Clin. Exp. Pathol. 2015, 8, 6547–6553. [Google Scholar]
- Zuo, Y.; Qu, C.; Tian, Y.; Wen, Y.; Xia, S.; Ma, M. The HIF-1/SNHG1/miR-199a-3p/TFAM Axis Explains Tumor Angiogenesis and Metastasis under Hypoxic Conditions in Breast Cancer. BioFactors Oxf. Engl. 2021, 47, 444–460. [Google Scholar] [CrossRef]
- Li, S.-Q.; Wang, Z.-H.; Mi, X.-G.; Liu, L.; Tan, Y. MiR-199a/b-3p Suppresses Migration and Invasion of Breast Cancer Cells by Downregulating PAK4/MEK/ERK Signaling Pathway. IUBMB Life 2015, 67, 768–777. [Google Scholar] [CrossRef]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-Sequence-Based microRNA Expression Signature in Breast Cancer: Tumor-Suppressive miR-101-5p Regulates Molecular Pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef]
- Harati, R.; Mohammad, M.G.; Tlili, A.; El-Awady, R.A.; Hamoudi, R. Loss of miR-101-3p Promotes Transmigration of Metastatic Breast Cancer Cells through the Brain Endothelium by Inducing COX-2/MMP1 Signaling. Pharmaceuticals 2020, 13, 144. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, M.; Zheng, X.; Zheng, L.; Liu, H.; Lu, J.; Liu, Y.; Chen, W. Interactions Between lncRNA TUG1 and miR-9-5p Modulate the Resistance of Breast Cancer Cells to Doxorubicin by Regulating eIF5A2. OncoTargets Ther. 2020, 13, 13159–13170. [Google Scholar] [CrossRef]
- Liu, D.-Z.; Chang, B.; Li, X.-D.; Zhang, Q.-H.; Zou, Y.-H. MicroRNA-9 Promotes the Proliferation, Migration, and Invasion of Breast Cancer Cells via down-Regulating FOXO1. Clin. Transl. Oncol. 2017, 19, 1133–1140. [Google Scholar] [CrossRef]
- D’Ippolito, E.; Plantamura, I.; Bongiovanni, L.; Casalini, P.; Baroni, S.; Piovan, C.; Orlandi, R.; Gualeni, A.V.; Gloghini, A.; Rossini, A.; et al. miR-9 and miR-200 Regulate PDGFRβ-Mediated Endothelial Differentiation of Tumor Cells in Triple-Negative Breast Cancer. Cancer Res. 2016, 76, 5562–5572. [Google Scholar] [CrossRef]
- Li, X.; Zeng, Z.; Wang, J.; Wu, Y.; Chen, W.; Zheng, L.; Xi, T.; Wang, A.; Lu, Y. MicroRNA-9 and Breast Cancer. Biomed. Pharmacother. 2020, 122, 109687. [Google Scholar] [CrossRef]
- Ma, L.; Young, J.; Prabhala, H.; Pan, E.; Mestdagh, P.; Muth, D.; Teruya-Feldstein, J.; Reinhardt, F.; Onder, T.T.; Valastyan, S.; et al. miR-9, a MYC/MYCN-Activated microRNA, Regulates E-Cadherin and Cancer Metastasis. Nat. Cell Biol. 2010, 12, 247–256. [Google Scholar] [CrossRef]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. microRNA-21 Promotes Breast Cancer Proliferation and Metastasis by Targeting LZTFL1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef]
- TNM Staging for Breast Cancer. Available online: https://www.cancerresearchuk.org/about-cancer/breast-cancer/stages-grades/tnm-staging (accessed on 26 September 2023).
- You, F.; Li, J.; Zhang, P.; Zhang, H.; Cao, X. miR106a Promotes the Growth of Transplanted Breast Cancer and Decreases the Sensitivity of Transplanted Tumors to Cisplatin. Cancer Manag. Res. 2020, 12, 233–246. [Google Scholar] [CrossRef]
- Loh, H.-Y.; Norman, B.P.; Lai, K.-S.; Rahman, N.M.A.N.A.; Alitheen, N.B.M.; Osman, M.A. The Regulatory Role of MicroRNAs in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef] [PubMed]
- TERF1 Telomeric Repeat Binding Factor 1 [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/7013 (accessed on 26 September 2023).
- Taha, M.; Mitwally, N.; Soliman, A.S.; Yousef, E. Potential Diagnostic and Prognostic Utility of miR-141, miR-181b1, and miR-23b in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 8589. [Google Scholar] [CrossRef]
- GeneCards—Human Genes|Gene Database|Gene Search. Available online: https://www.genecards.org/ (accessed on 26 September 2023).
- Bertoli, G.; Cava, C.; Castiglioni, I. MicroRNAs: New Biomarkers for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Breast Cancer. Theranostics 2015, 5, 1122–1143. [Google Scholar] [CrossRef]
- Gao, Y.; Zeng, F.; Wu, J.-Y.; Li, H.-Y.; Fan, J.-J.; Mai, L.; Zhang, J.; Ma, D.-M.; Li, Y.; Song, F. MiR-335 Inhibits Migration of Breast Cancer Cells through Targeting Oncoprotein c-Met. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2015, 36, 2875–2883. [Google Scholar] [CrossRef]
- Ye, L.; Wang, F.; Wu, H.; Yang, H.; Yang, Y.; Ma, Y.; Xue, A.; Zhu, J.; Chen, M.; Wang, J.; et al. Functions and Targets of miR-335 in Cancer. OncoTargets Ther. 2021, 14, 3335–3349. [Google Scholar] [CrossRef]
- Alhasan, L. MiR-126 Modulates Angiogenesis in Breast Cancer by Targeting VEGF-A -mRNA. Asian Pac. J. Cancer Prev. APJCP 2019, 20, 193–197. [Google Scholar] [CrossRef]
- Soofiyani, S.R.; Hosseini, K.; Ebrahimi, T.; Forouhandeh, H.; Sadeghi, M.; Beirami, S.M.; Ghasemnejad, T.; Tarhriz, V.; Montazersaheb, S. Prognostic Value and Biological Role of miR-126 in Breast Cancer. MicroRNA Shariqah United Arab Emir. 2022, 11, 95–103. [Google Scholar] [CrossRef]
- Qattan, A.; Al-Tweigeri, T.; Alkhayal, W.; Suleman, K.; Tulbah, A.; Amer, S. Clinical Identification of Dysregulated Circulating microRNAs and Their Implication in Drug Response in Triple Negative Breast Cancer (TNBC) by Target Gene Network and Meta-Analysis. Genes 2021, 12, 549. [Google Scholar] [CrossRef]
- Fan, X.; Zhou, S.; Zheng, M.; Deng, X.; Yi, Y.; Huang, T. MiR-199a-3p Enhances Breast Cancer Cell Sensitivity to Cisplatin by Downregulating TFAM (TFAM). Biomed. Pharmacother. 2017, 88, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Fuso, P.; Di Salvatore, M.; Santonocito, C.; Guarino, D.; Autilio, C.; Mulè, A.; Arciuolo, D.; Rinninella, A.; Mignone, F.; Ramundo, M.; et al. Let-7a-5p, miR-100-5p, miR-101-3p, and miR-199a-3p Hyperexpression as Potential Predictive Biomarkers in Early Breast Cancer Patients. J. Pers. Med. 2021, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Z.; Deng, F.; Li, H.; Wang, D.-D.; Zhang, W.; Ding, L.; Tang, J.-H. MiR-101: A Potential Therapeutic Target of Cancers. Am. J. Transl. Res. 2018, 10, 3310–3321. [Google Scholar] [PubMed]
- Jiang, H.; Li, L.; Zhang, J.; Wan, Z.; Wang, Y.; Hou, J.; Yu, Y. MiR-101-3p and Syn-Cal14.1a Synergy in Suppressing EZH2-Induced Progression of Breast Cancer. OncoTargets Ther. 2020, 13, 9599–9609. [Google Scholar] [CrossRef]
- Sporn, J.C.; Katsuta, E.; Yan, L.; Takabe, K. Expression of MicroRNA-9 Is Associated with Overall Survival in Breast Cancer Patients. J. Surg. Res. 2019, 233, 426–435. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Bonci, D.; Coppola, V.; Musumeci, M.; Addario, A.; Giuffrida, R.; Memeo, L.; D’Urso, L.; Pagliuca, A.; Biffoni, M.; Labbaye, C.; et al. The miR-15a-miR-16-1 Cluster Controls Prostate Cancer by Targeting Multiple Oncogenic Activities. Nat. Med. 2008, 14, 1271–1277. [Google Scholar] [CrossRef]
- Long, X.; Shi, Y.; Ye, P.; Guo, J.; Zhou, Q.; Tang, Y. MicroRNA-99a Suppresses Breast Cancer Progression by Targeting FGFR3. Front. Oncol. 2020, 9, 1473. [Google Scholar] [CrossRef]
- Menon, A.; Abd-Aziz, N.; Khalid, K.; Poh, C.L.; Naidu, R. miRNA: A Promising Therapeutic Target in Cancer. Int. J. Mol. Sci. 2022, 23, 11502. [Google Scholar] [CrossRef]
- Shah, V.; Shah, J. Recent Trends in Targeting miRNAs for Cancer Therapy. J. Pharm. Pharmacol. 2020, 72, 1732–1749. [Google Scholar] [CrossRef]
- Baumann, V.; Winkler, J. miRNA-Based Therapies: Strategies and Delivery Platforms for Oligonucleotide and Non-Oligonucleotide Agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA Therapeutics: Towards a New Era for the Management of Cancer and Other Diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The Role and Mechanisms of Action of MicroRNAs in Cancer Drug Resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.; Shi, L.; Garofalo, M. Role of MicroRNAs in Chemoresistance. Ann. Transl. Med. 2015, 3, 332. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Ma, T.; Zhang, X.; Lv, M.; Chen, L.; Tang, J.; Zhao, J. MicroRNA Expression Profiling and Bioinformatics Analysis of Dysregulated MicroRNAs in Vinorelbine-Resistant Breast Cancer Cells. Gene 2015, 556, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.-D.; Ye, X.-M.; Zhang, M.-Y.; Zhu, H.-Y.; Xi, W.-J.; Huang, X.; Zhao, J.; Gu, B.; Zheng, G.-X.; Yang, A.-G.; et al. MiR-200c Suppresses TGF-β Signaling and Counteracts Trastuzumab Resistance and Metastasis by Targeting ZNF217 and ZEB1 in Breast Cancer: MiR-200c/ZNF217/TGF-β Regulates Trastuzumab Resistance. Int. J. Cancer 2014, 135, 1356–1368. [Google Scholar] [CrossRef]
- Song, L.; Liu, D.; Wang, B.; He, J.; Zhang, S.; Dai, Z.; Ma, X.; Wang, X. MiR-494 Suppresses the Progression of Breast Cancer In Vitro by Targeting CXCR4 through the Wnt/β-Catenin Signaling Pathway. Oncol. Rep. 2015, 34, 525–531. [Google Scholar] [CrossRef]
- Abedi, N.; Mohammadi-Yeganeh, S.; Koochaki, A.; Karami, F.; Paryan, M. MiR-141 as Potential Suppressor of β-Catenin in Breast Cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2015, 36, 9895–9901. [Google Scholar] [CrossRef]
- Yu, Z.; Xu, Z.; Disante, G.; Wright, J.; Wang, M.; Li, Y.; Zhao, Q.; Ren, T.; Ju, X.; Gutman, E.; et al. MiR-17/20 Sensitization of Breast Cancer Cells to Chemotherapy-Induced Apoptosis Requires Akt1. Oncotarget 2014, 5, 1083–1090. [Google Scholar] [CrossRef]
- He, X.; Xiao, X.; Dong, L.; Wan, N.; Zhou, Z.; Deng, H.; Zhang, X. MiR-218 Regulates Cisplatin Chemosensitivity in Breast Cancer by Targeting BRCA1. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2015, 36, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cano, I.; Pattanayak, B.; Adam-Artigues, A.; Lameirinhas, A.; Torres-Ruiz, S.; Tormo, E.; Cervera, R.; Eroles, P. MicroRNAs as a Clue to Overcome Breast Cancer Treatment Resistance. Cancer Metastasis Rev. 2022, 41, 77–105. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials. Available online: https://clinicaltrials.gov/ (accessed on 27 September 2023).
- Pan, G.; Liu, Y.; Shang, L.; Zhou, F.; Yang, S. EMT-Associated MicroRNAs and Their Roles in Cancer Stemness and Drug Resistance. Cancer Commun. Lond. Engl. 2021, 41, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Farzaneh, M. Signaling Pathways Governing Breast Cancer Stem Cells Behavior. Stem Cell Res. Ther. 2021, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef]
- Yousefnia, S.; Seyed Forootan, F.; Seyed Forootan, S.; Nasr Esfahani, M.H.; Gure, A.O.; Ghaedi, K. Mechanistic Pathways of Malignancy in Breast Cancer Stem Cells. Front. Oncol. 2020, 10, 452. [Google Scholar] [CrossRef]
- Kastl, L.; Brown, I.; Schofield, A.C. MiRNA-34a Is Associated with Docetaxel Resistance in Human Breast Cancer Cells. Breast Cancer Res. Treat. 2012, 131, 445–454. [Google Scholar] [CrossRef]
- Zhao, X.; Li, J.; Huang, S.; Wan, X.; Luo, H.; Wu, D. MiRNA-29c regulates cell growth and invasion by targeting CDK6 in bladder cancer. Am. J. Transl. Res. 2015, 7, 1382–1389. [Google Scholar]
- Citron, F.; Segatto, I.; Vinciguerra, G.L.R.; Musco, L.; Russo, F.; Mungo, G.; D’Andrea, S.; Mattevi, M.C.; Perin, T.; Schiappacassi, M.; et al. Downregulation of miR-223 Expression Is an Early Event during Mammary Transformation and Confers Resistance to CDK4/6 Inhibitors in Luminal Breast Cancer. Cancer Res. 2020, 80, 1064–1077. [Google Scholar] [CrossRef]
- Bisso, A.; Faleschini, M.; Zampa, F.; Capaci, V.; De Santa, J.; Santarpia, L.; Piazza, S.; Cappelletti, V.; Daidone, M.; Agami, R.; et al. Oncogenic MiR-181a/b Affect the DNA Damage Response in Aggressive Breast Cancer. Cell Cycle Georget. Tex 2013, 12, 1679–1687. [Google Scholar] [CrossRef]
- Feng, D.; Zhang, H.; Zhang, P.; Zheng, Y.; Zhang, X.; Han, B.; Luo, X.; Xu, L.; Zhou, H.; Qu, L.; et al. Down-regulated MiR-331–5p and MiR-27a Are Associated with Chemotherapy Resistance and Relapse in Leukaemia. J. Cell. Mol. Med. 2011, 15, 2164–2175. [Google Scholar] [CrossRef]
- Zou, Z.; Zou, R.; Zong, D.; Shi, Y.; Chen, J.; Huang, J.; Zhu, J.; Chen, L.; Bao, X.; Liu, Y.; et al. MiR-495 Sensitizes MDR Cancer Cells to the Combination of Doxorubicin and Taxol by Inhibiting MDR1 Expression. J. Cell. Mol. Med. 2017, 21, 1929–1943. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, R.; Wang, M.; Kumar, A.K.; Pan, F.; He, L.; Hu, Z.; Guo, Z. MicroRNA-140 Impedes DNA Repair by Targeting FEN1 and Enhances Chemotherapeutic Response in Breast Cancer. Oncogene 2020, 39, 234–247. [Google Scholar] [CrossRef]
- Zhao, W.; Li, H.; Yang, S.; Guo, D.; Chen, J.; Miao, S.; Xin, Y.; Liang, M. MicroRNA-152 Suppresses Cisplatin Resistance in A549 Cells. Oncol. Lett. 2019, 18, 4613–4620. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Wang, S.; Zhao, Y.; Zhang, Z.; Qin, C.; Yang, X. MiR-519d Impedes Cisplatin-Resistance in Breast Cancer Stem Cells by down-Regulating the Expression of MCL-1. Oncotarget 2017, 8, 22003–22013. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Tian, W.; Chen, H.; Jiang, K. MiR-944 Functions as a Novel Oncogene and Regulates the Chemoresistance in Breast Cancer. Tumor Biol. 2016, 37, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Ren, T.; Huang, Y.; Sun, K.; Wang, S.; Liu, K.; Zheng, B.; Guo, W. Knockdown of Long Non-Coding RNA HOTAIR Increases MiR-454-3p by Targeting Stat3 and Atg12 to Inhibit Chondrosarcoma Growth. Cell Death Dis. 2017, 8, e2605. [Google Scholar] [CrossRef]
- Chong, Z.X.; Yeap, S.K.; Ho, W.Y. Regulation of autophagy by microRNAs in human breast cancer. J. Biomed. Sci. 2021, 28, 21. [Google Scholar] [CrossRef]
- Akkoc, Y.; Gozuacik, D. MicroRNAs as Major Regulators of the Autophagy Pathway. Biochim. Biophys. Acta BBA Mol. Cell Res. 2020, 1867, 118662. [Google Scholar] [CrossRef]
- Shan, C.; Chen, X.; Cai, H.; Hao, X.; Li, J.; Zhang, Y.; Gao, J.; Zhou, Z.; Li, X.; Liu, C.; et al. The Emerging Roles of Autophagy-Related MicroRNAs in Cancer. Int. J. Biol. Sci. 2021, 17, 134–150. [Google Scholar] [CrossRef]
- UCSC Xena. Available online: https://xenabrowser.net/ (accessed on 27 September 2023).
- GEPIA 2. Available online: http://gepia2.cancer-pku.cn/#index (accessed on 9 February 2023).
- OncoLnc. Available online: http://www.oncolnc.org/ (accessed on 27 September 2023).
- Boon, R.A.; Vickers, K.C. Intercellular Transport of microRNAs. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Makarova, J.; Turchinovich, A.; Shkurnikov, M.; Tonevitsky, A. Extracellular miRNAs and Cell–Cell Communication: Problems and Prospects. Trends Biochem. Sci. 2021, 46, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cheng, Z.; Wang, Y.; Han, T. The Risks of miRNA Therapeutics: In a Drug Target Perspective. Drug Des. Devel. Ther. 2021, 15, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Momin, M.Y.; Gaddam, R.R.; Kravitz, M.; Gupta, A.; Vikram, A. The Challenges and Opportunities in the Development of MicroRNA Therapeutics: A Multidisciplinary Viewpoint. Cells 2021, 10, 3097. [Google Scholar] [CrossRef] [PubMed]
- Geary, R.S.; Norris, D.; Yu, R.; Bennett, C.F. Pharmacokinetics, Biodistribution and Cell Uptake of Antisense Oligonucleotides. Adv. Drug Deliv. Rev. 2015, 87, 46–51. [Google Scholar] [CrossRef]
- McMahon, B.M.; Mays, D.; Lipsky, J.; Stewart, J.A.; Fauq, A.; Richelson, E. Pharmacokinetics and Tissue Distribution of a Peptide Nucleic Acid after Intravenous Administration. Antisense Nucleic Acid Drug Dev. 2002, 12, 65–70. [Google Scholar] [CrossRef]
- Cacheux, J.; Bancaud, A.; Leichlé, T.; Cordelier, P. Technological Challenges and Future Issues for the Detection of Circulating MicroRNAs in Patients with Cancer. Front. Chem. 2019, 7, 815. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Nagaraj, S. Pros and Cons of miRNAs as Non-Invasive Circulatory Biomarkers. Res. Rev. Res. J. Biol. 2017, 5, 1–7. [Google Scholar]


{kind=link}
| miRNA | Regulation in Breast Cancer Cells | Source and Detection Method | Target | Target Effects/Action | Metabolic Consequences |
|---|---|---|---|---|---|
| miR-21 | Upregulated | Serum, qRT-PCR [60] | PTEN [44,58] | Downregulation of PTEN expression [44,58,61] | Drug resistance to doxorubicin in HER2- BC cells [61] |
| * miR-21 inhibition induces PTEN expression [62] | * Restored trastuzumab sensitivity in the resistant BC xenografts in vivo [62] | ||||
| PTEN/Akt [63] | Downregulation of PTEN expression and Akt activation [63] | Induction of EMT and gemcitabine resistance [63] | |||
| PI3K/Akt, MEK/ERK [58] | Activation PI3K/Akt and MEK/ERK signaling pathways [58] | Development of MDR [58] | |||
| TPM1, TGF-β [64] | Repression of expression TPM1 [65,66] | Increased BC cells proliferation, migration, invasion, survival, and EMT [64] | |||
| Mesenchymal cell markers (N-cadherin, Vimentin, α-SMA) [67] | Activation of mesenchymal cell markers [67] | Re-expression of miR-21 is responsible for migration and invasion by activating the EMT process in MCF7 cells [67] | |||
| Epithelial cell marker (E-cadherin) [67] | Inhibition of epithelial cell marker [67] | ||||
| miR-106a | Upregulated | Serum, qRT-PCR [68] | Bcl-2,ABCG2, BAX, P53, RUNX3 [69] | Upregulation of Bcl-2 protein and multidrug transporter ABCG2. Downregulation of BAX protein and genes products: P53, RUNX3 [69] | Promotes BC cells proliferation and invasion [69] |
| * Inhibition of miR-106a downregulates the expression of Bcl-2, ABCG2 and upregulates the BAX, P53, RUNX3 expression [69] | |||||
| RAF-1 [68] | Decreases RAF-1 levels and RAF-1 is a part of MAPK/ERK signaling pathway [68] | Possibly induces proliferation and decreases apoptosis in BC cells through regulation of the MAPK/ERK signaling pathway, which controls gene expression [68] | |||
| ZBTB4 [70,71,72] | Negative regulation of ZBTB4 gene, which functions as a tumor suppressor gene [70,71,72] | * Restoration of ZBTB4 suppress Sp1, Sp3, Sp4 expression resulting in inhibition of BC cells proliferation, invasion [70,71,72] | |||
| miR-155 | Upregulated | Serum, qRT-PCR [73] | TERF1 [74] | Reduction in the shelterin component TRF1 expression. TRF1 regulates telomere length and suppresses DNA breakage [74] | Antagonization of telomere integrity in BC cells and increased genomic instability [74] |
| SOCS1 [75] | Repression of SOCS1 (negative feedback regulator of JAK/STAT signaling) [75] | Constitutive activation of STAT3 in BC cells, promotion of cell proliferation and colony formation [75] | |||
| C/EBPβ [76,77] | Loss of CCAAT-enhancer binding protein beta (C/EBPβ) [76,77] | Modification of TGF-β response; from growth inhibition to EMT, invasion, and metastasis in BC. Promotion of BC progression [76,77] | |||
| mir-141 | Downregulated | Tissue, qRT-PCR, Microarray [78] | ANP32E [78] | Regulation of ANP32E (positive regulator of tumor growth and metastasis) [78,79] | ANP32E induces tumorigenesis of BC by upregulating E2F1 and promoting the G1/S transition [79] |
| ** Overexpression of miR-141 downregulated ANP32E expression [78] | ** Inhibition of BC cells proliferation, migration, and invasion [78] | ||||
| SIP1 [80] | Regulation of EMT [80] | EMT plays a crucial role in early tumor metastasis and SIP1 is a promoter of cancer progression [80] | |||
| let-7c | Downregulated | Serum, qRT-PCR [81] | ERCC6 [82] | Upregulation of ERCC6 [82] | Intensified cancer growth ability and lower rate of apoptosis; DNA damage accumulation [82] |
| BCL2, BAX [83] | ** Overexpression of let-7c decreases level of Bcl-2 and increases the level of BAX, TP53, PTEN [83] | ** Promotion of apoptotic cell death, suppression of cancer progression [83] | |||
| ERα and Wnt signaling [84] | ** Overexpression of let-7c inhibits estrogen induction in ERα and Wnt signaling [84] | ** Inhibition of BCSCs self-renew and suppresses tumor formation [84] | |||
| miR-335 | Downregulated | Serum, qRT-PCR [85] | BRCA1 [86,87] | Downregulation of BRCA1 [86] | Accelerated tumor growth, genomic instability, BC progression [86] |
| ** Overexpression of miR-335 upregulates the level of BRCA1 [86,87] | ** Decreased cell viability and increased apoptosis [86,87] | ||||
| miR-126 | Downregulated | Tissue, qRT-PCR [88] | VEGFA [88], PIK3R2 [89] | Inactivation of the PIK3R2/PI3K/Akt/mTOR signaling pathway [89] | Vasculogenesis, angiogenesis resulting in tumor growth [88] Resistance to trastuzumab [89] in SKBR3 and BT747 cell lines |
| ADAM9 [90] | ** Upregulation of miR-126 is silencing ADAM9 gene [90] | ** Inhibition of BC cells invasion and metastasis [90] | |||
| miR-199a | Downregulated | Tissue, qRT-PCR [91] | PAK4/MEK/ERK signaling pathway [92] | Regulation of PAK4/MEK/ERK signaling pathway [92] | PAK4 activates the ERK pathway, and MEK/ERK pathway plays a part in PAK4-induced cell growth regulation [92] |
| ** MiR-199a/b-3p downregulates PAK4 expression and PAK4/MEK/ERK signaling pathway [92] | ** Suppression of BC cells migration and invasion [92] | ||||
| miR-101 | Downregulated | Tissue, qRT-PCR [93] | COX-2/MMP1 signaling pathway [94] | Upregulation of COX-2/MMP1 signaling pathway [94] | Promotes transmigration of metastatic BC cells through the brain endothelium [94] |
| ** Restoring miR-101-3p in BC cells reduces COX-2/MMP1 expression [94] | ** Reduction in transmigratory ability [94] | ||||
| miR-9 | Upregulated | Cell culture, qRT-PCR [95] | FOXO1 [96] | Downregulation of FOXO1 expression [96] | Promotion of proliferation, migration, and invasion of BC cells [96] |
| STARD13 [97] | Repression of STARD13 [97] | Upon stimulation of PDGFRβ signaling, miR-9 could promote the formation of vascular-like structures of TNBC [97] | |||
| E-cadherin [96,98] | E-cadherin downregulation [98,99] | Increased tumor angiogenesis [99] Primes BC cells to EMT and invasion [98] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziechciowska, I.; Dąbrowska, M.; Mizielska, A.; Pyra, N.; Lisiak, N.; Kopczyński, P.; Jankowska-Wajda, M.; Rubiś, B. miRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy. Curr. Issues Mol. Biol. 2023, 45, 9500-9525. https://doi.org/10.3390/cimb45120595
Dziechciowska I, Dąbrowska M, Mizielska A, Pyra N, Lisiak N, Kopczyński P, Jankowska-Wajda M, Rubiś B. miRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy. Current Issues in Molecular Biology. 2023; 45(12):9500-9525. https://doi.org/10.3390/cimb45120595
Chicago/Turabian StyleDziechciowska, Iga, Małgorzata Dąbrowska, Anna Mizielska, Natalia Pyra, Natalia Lisiak, Przemysław Kopczyński, Magdalena Jankowska-Wajda, and Błażej Rubiś. 2023. "miRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy" Current Issues in Molecular Biology 45, no. 12: 9500-9525. https://doi.org/10.3390/cimb45120595
APA StyleDziechciowska, I., Dąbrowska, M., Mizielska, A., Pyra, N., Lisiak, N., Kopczyński, P., Jankowska-Wajda, M., & Rubiś, B. (2023). miRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy. Current Issues in Molecular Biology, 45(12), 9500-9525. https://doi.org/10.3390/cimb45120595