
Therapeutic Effects of Aloe saponaria against Ulcerative Colitis Induced by Dextran Sulfate Sodium



Abstract
:1. Introduction
2. Materials and Methods
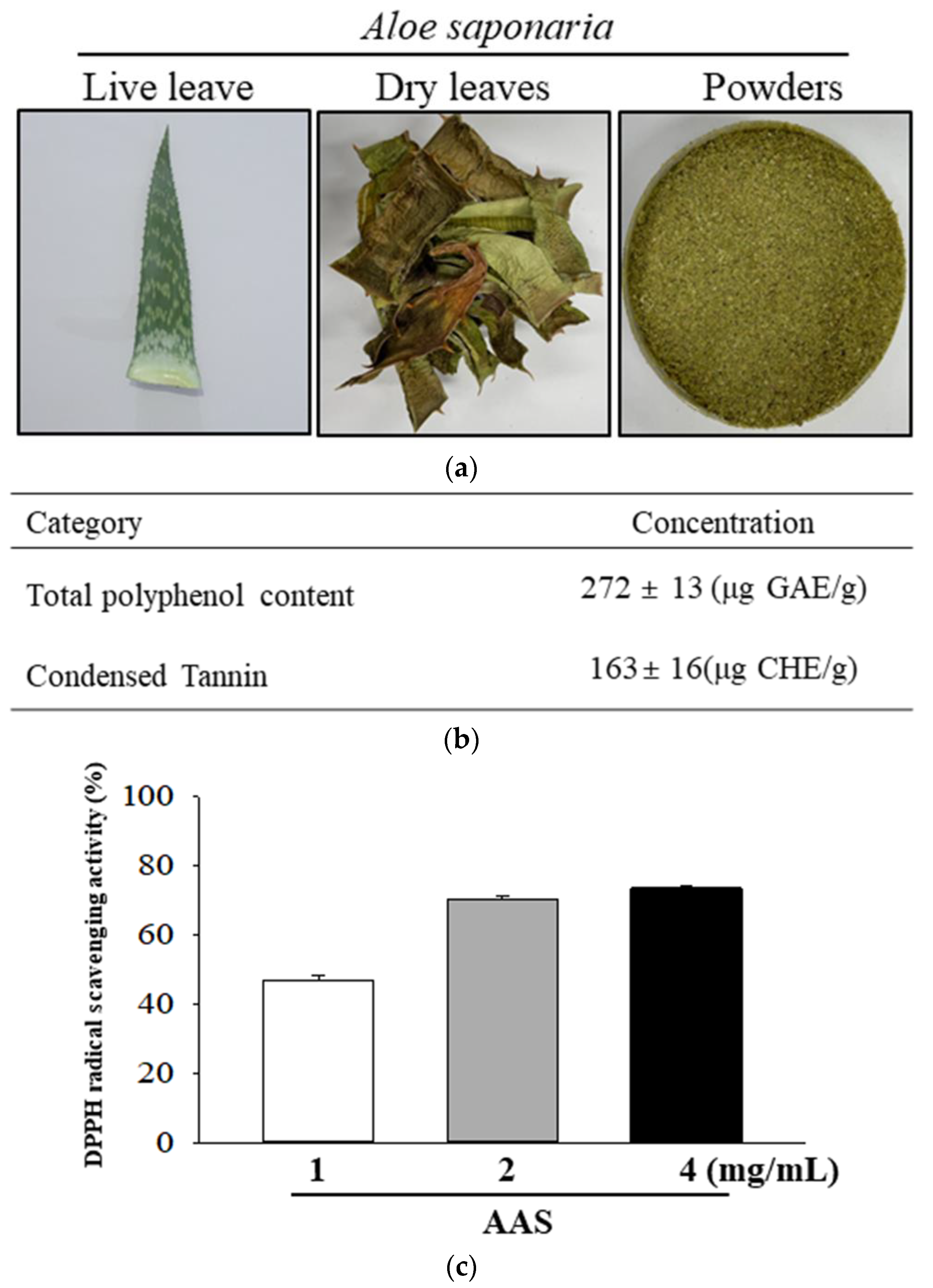
2.1. Preparation of A. saponaria (AAS)
2.2. Determination of Total Polyphenol Content and Tannin
2.3. Free Radical Scavenging Activity Analysis
2.4. Experimental Design for the Animal Study
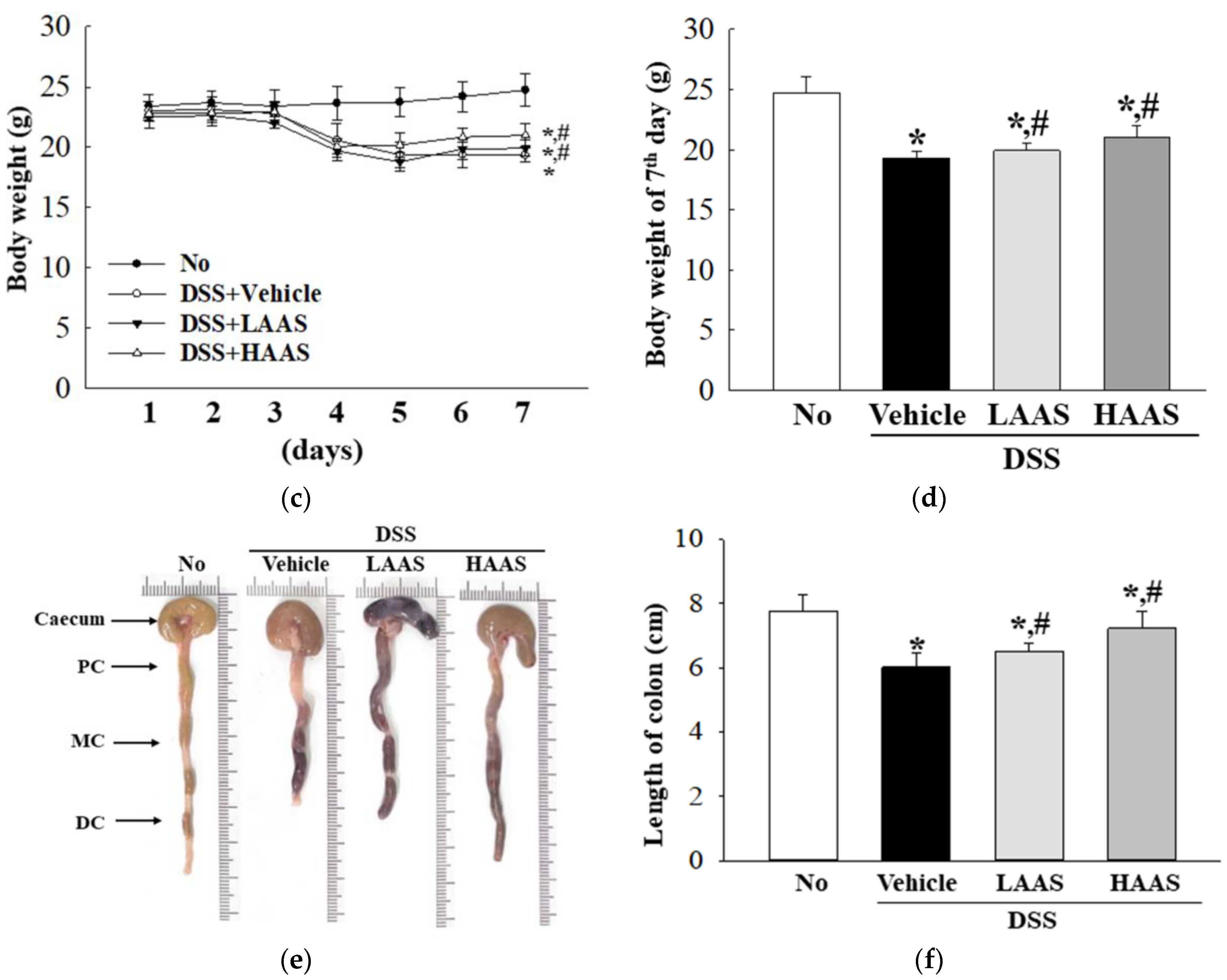
2.5. Measurement of Body and Organ Weights
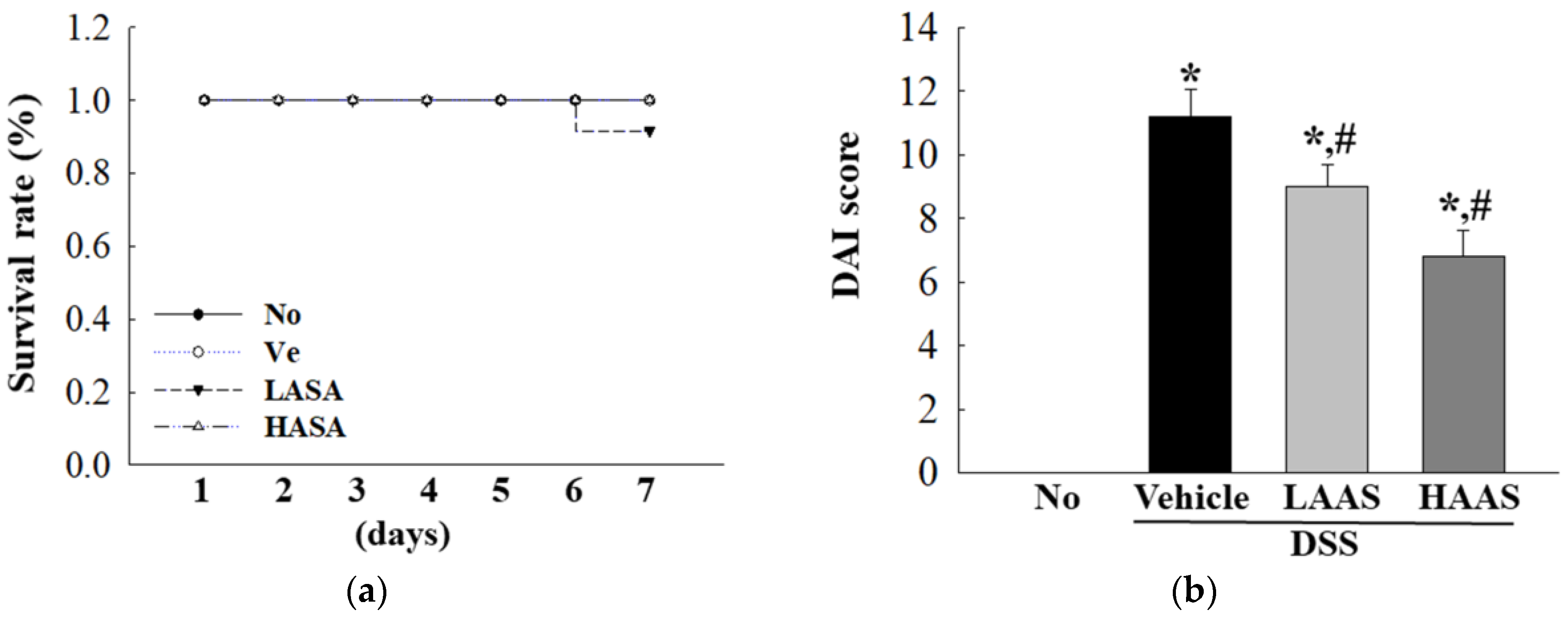
2.6. Disease Activity Index (DAI) Scoring System
2.7. Measurement of Colon Length
2.8. Serum Biochemical Analysis
2.9. Quantitative Real Time-PCR (RT-qPCR) Analysis
2.10. Western Blot Analysis
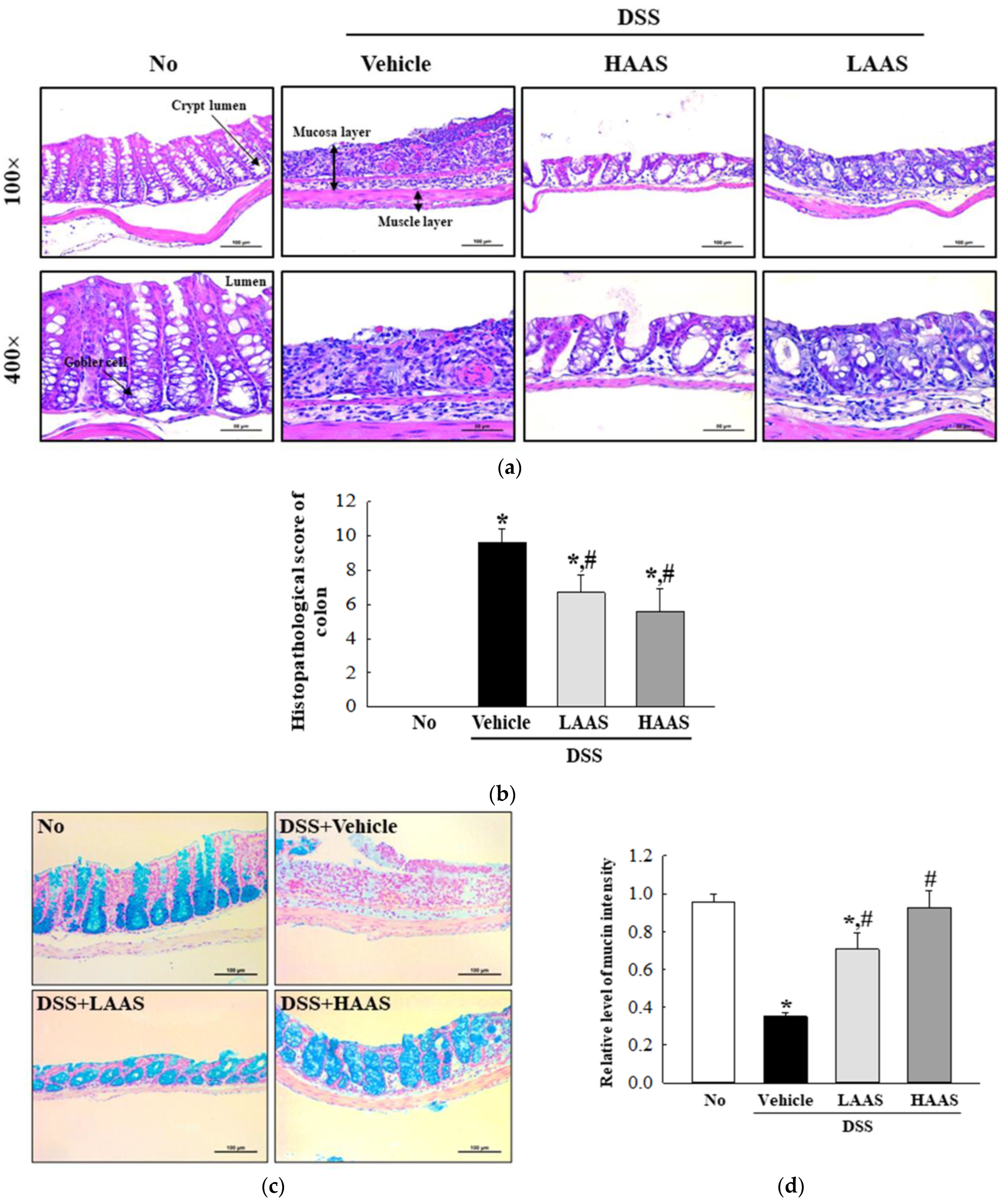
2.11. Histopathological Analysis
2.12. Statistical Significance Analysis
3. Results
3.1. Active Components and Antioxidant Activity of AAS
3.2. Treatment with AAS Improves Clinical Symptoms of UC in DSS-Induced UC Mice
3.3. Treatment with AAS Leads to Improvement in the Morphological and Histopathological Characteristics of the Colon in DSS-Induced UC Mice
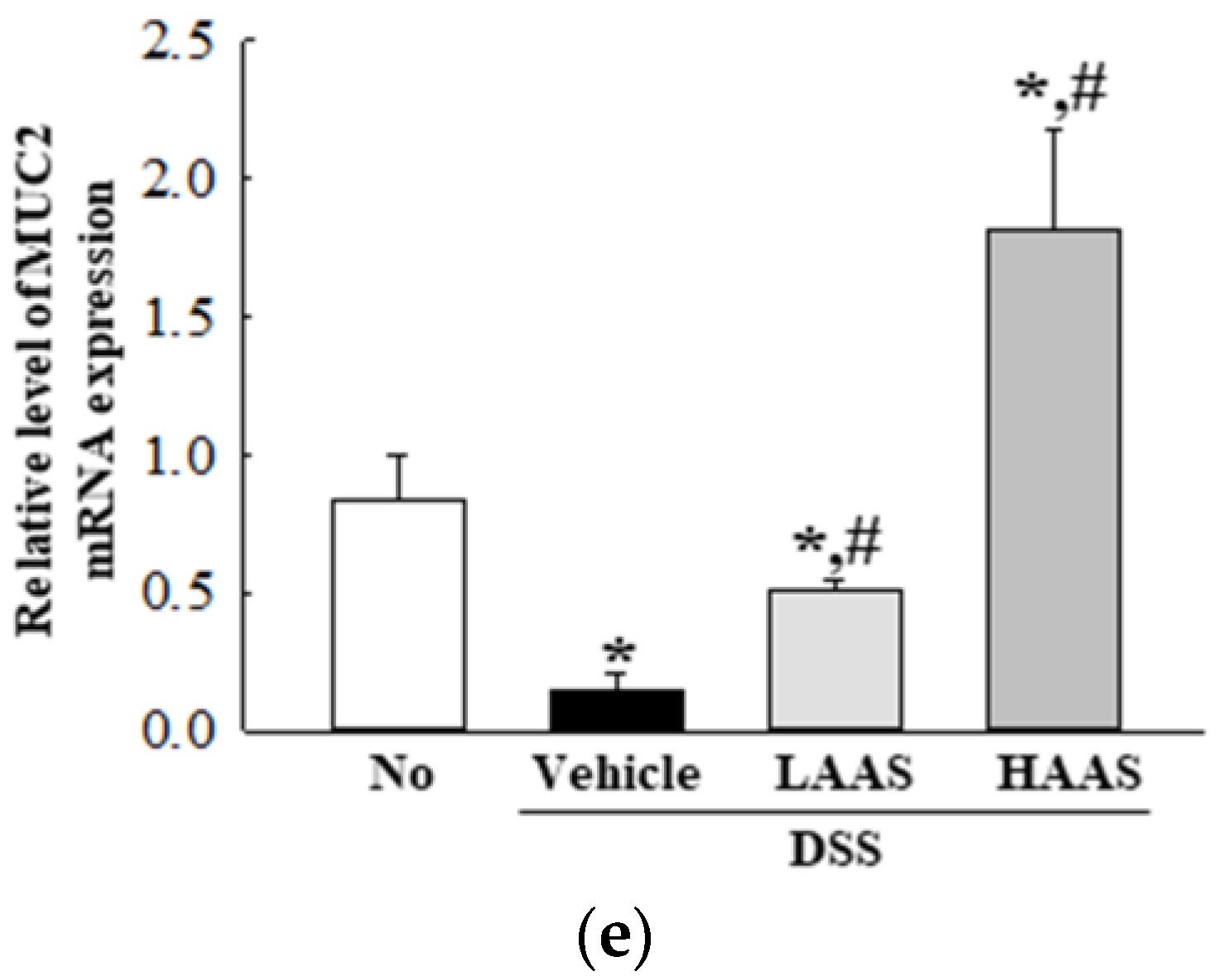
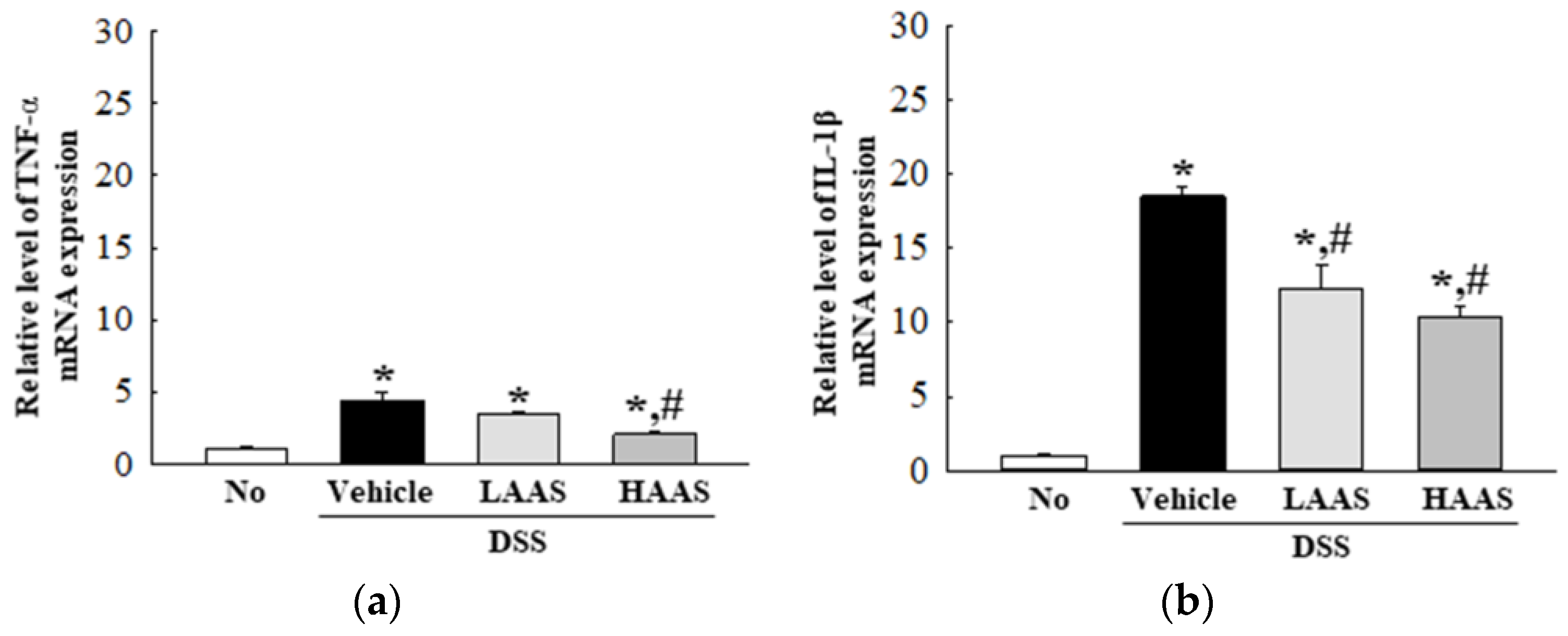
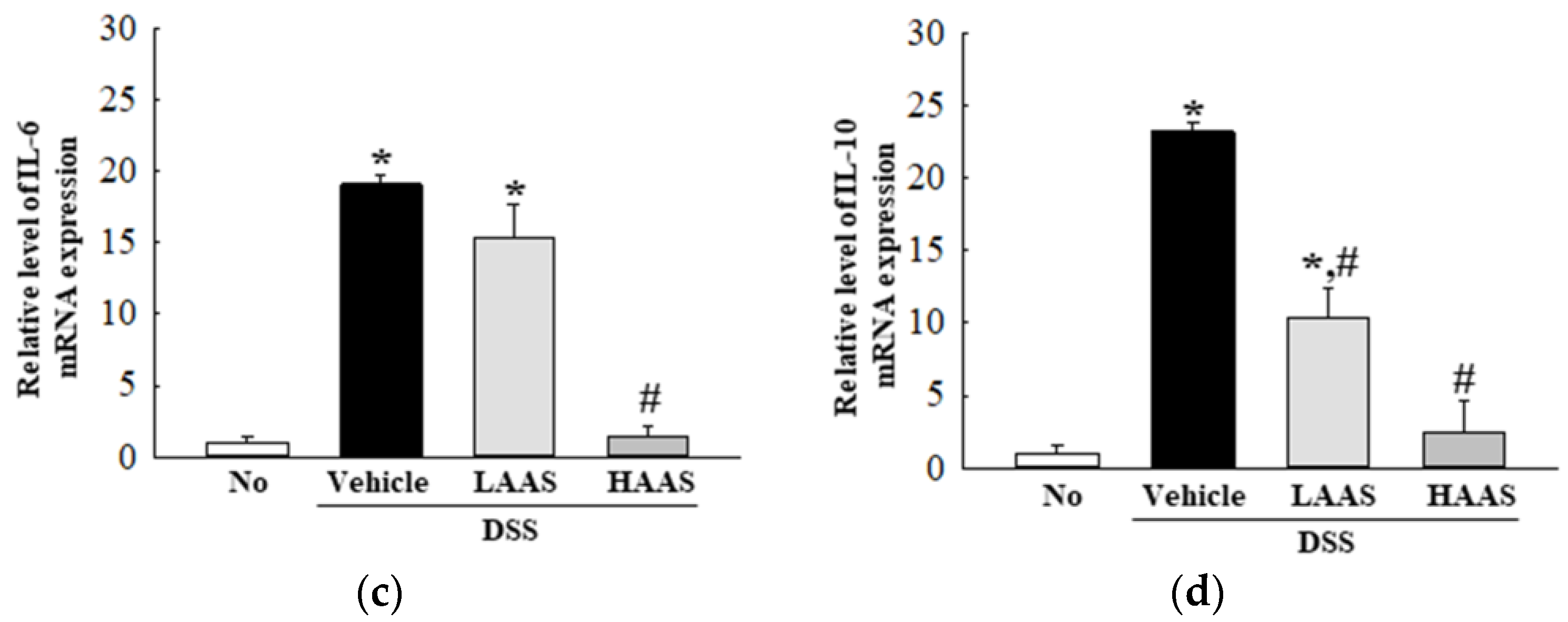
3.4. Treatment with AAS Attenuated the Inflammatory Response in the Colon of DSS-Induced UC Mice
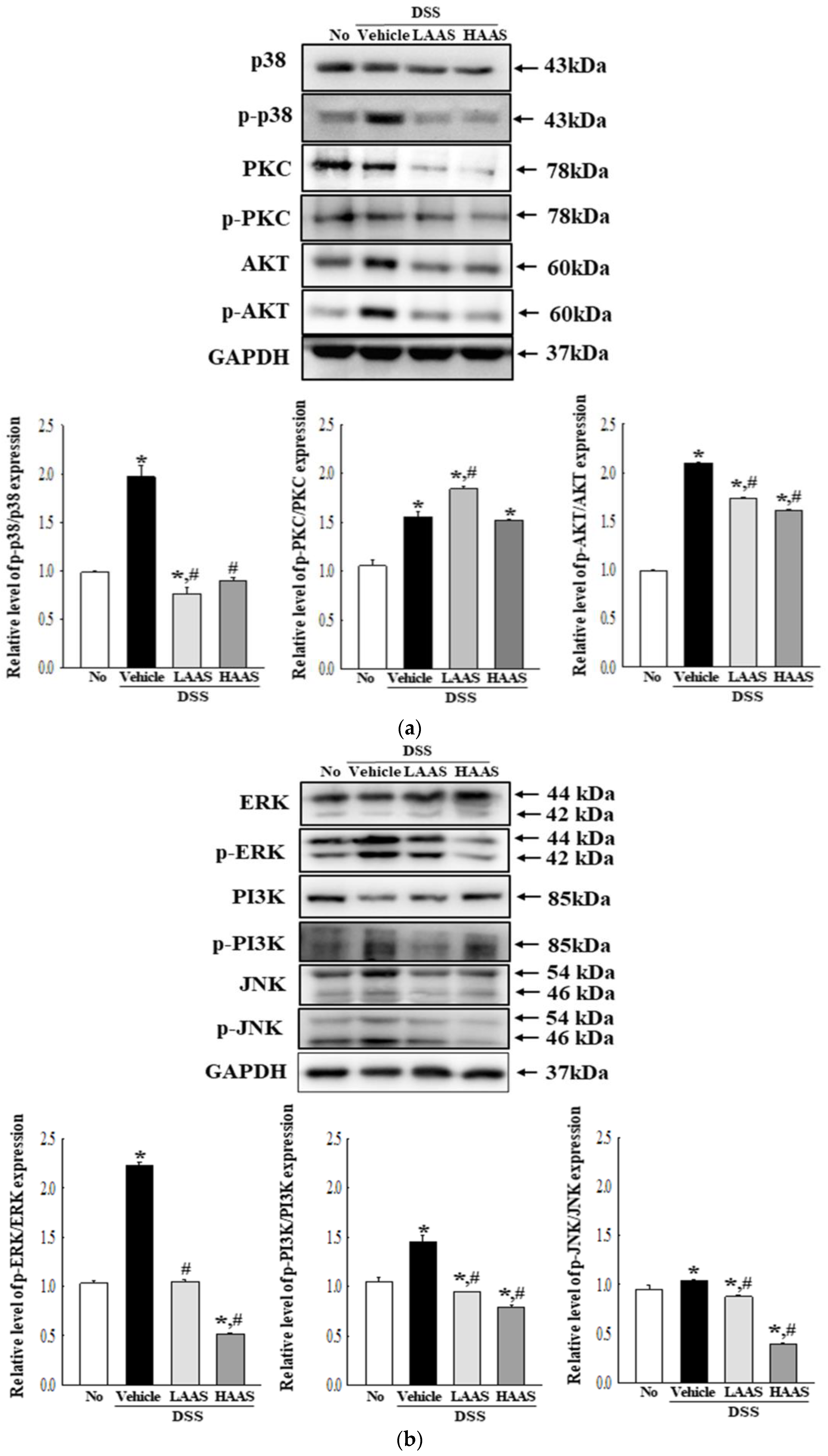
3.5. Effects of AAS on the PI3K/Akt and PKC/ERK Signaling Pathway in the Colon of DSS-Induced UC Mice
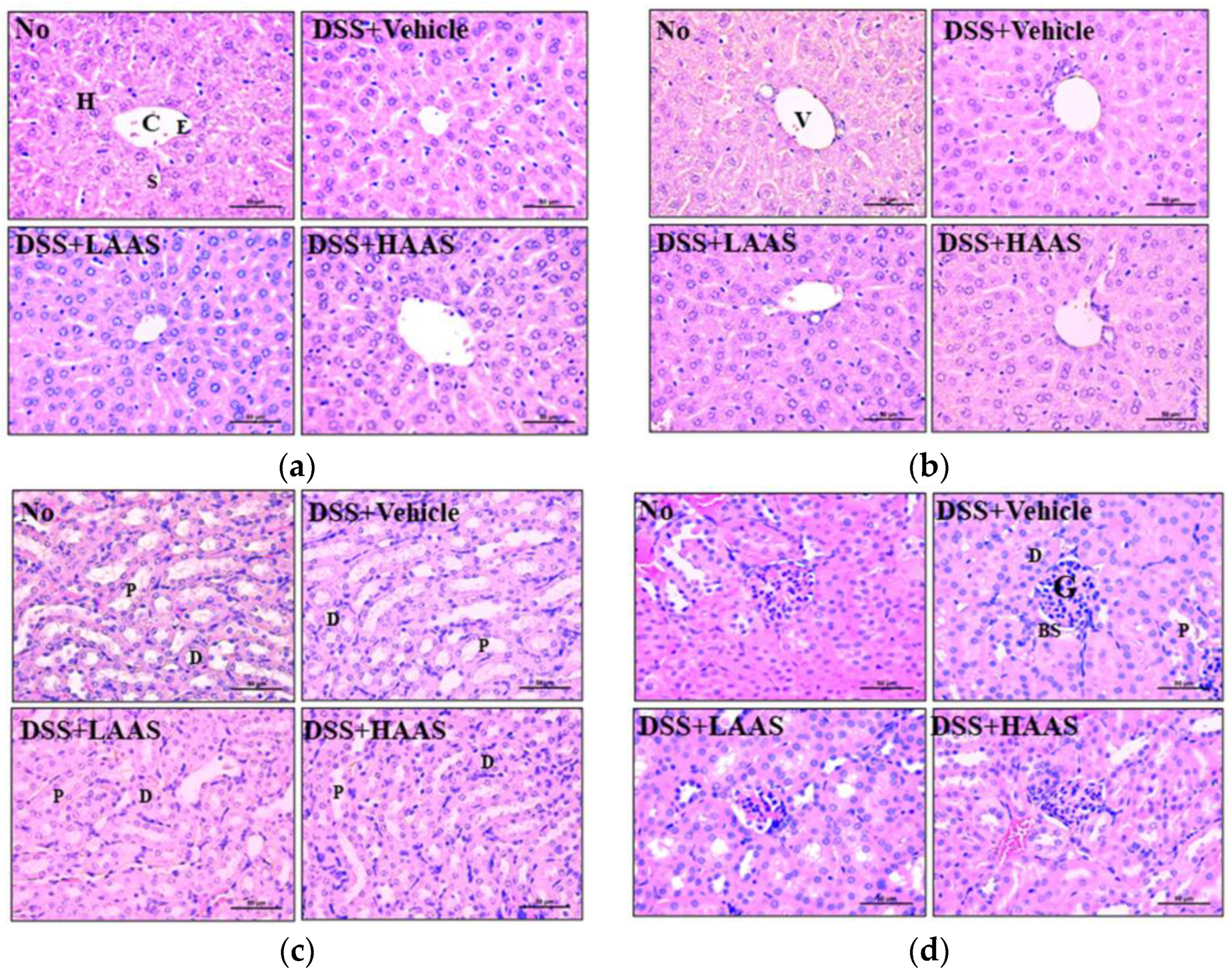
3.6. Hepatotoxicity and Nephrotoxicity of AAS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar]
- Gajendran, M.; Loganathan, P.; Jimenez, G.; Catinella, A.P.; Ng, N.; Umapathy, C.; Ziade, N.; Hashash, J.G. A comprehensive review and update on ulcerative colitis. Dis. A Mon. 2019, 65, 100851. [Google Scholar]
- Lombardi, V.R.; Etcheverría, I.; Carrera, I.; Cacabelos, R.; Chacón, A.R. Prevention of chronic experimental colitis induced by dextran sulphate sodium (DSS) in mice treated with FR91. J. Biomed. Biotechnol. 2012, 2012, 826178. [Google Scholar] [CrossRef]
- Turner, D.; Levine, A.; Escher, J.C.; Griffiths, A.M.; Russell, R.K.; Dignass, A.; Dias, J.A.; Bronsky, J.; Braegger, C.P.; Cucchiara, S.; et al. Management of pediatric ulcerative colitis: Joint ECCO and ESPGHAN evidence-based consensus guidelines. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 340–361. [Google Scholar] [CrossRef]
- Bentley, E.; Jenkins, D.; Campbell, F.; Warren, B. How could pathologists improve the initial diagnosis of colitis? Evidence from an international workshop. J. Clin. Pathol. 2002, 55, 955–960. [Google Scholar]
- Seldenrijk, C.A.; Morson, B.C.; Meuwissen, S.G.; Schipper, N.W.; Lindeman, J.; Meijer, C.J. Histopathological evaluation of colonic mucosal biopsy specimens in chronic inflammatory bowel disease: Diagnostic implications. Gut 1991, 32, 1514–1520. [Google Scholar]
- Lu, A.; Magupalli, V.G.; Ruan, J.; Yin, Q.; Atianand, M.K.; Vos, M.; Schröder, G.F.; Fitzgerald, K.A.; Wu, H.; Egelman, E.H. Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell 2014, 156, 1193–1206. [Google Scholar]
- Rahimian, R.; Zirak, M.R.; Keshavarz, M.; Fakhraei, N.; Mohammadi-Farani, A.; Hamdi, H.; Mousavizadeh, K. Involvement of PPAR γ in the protective action of tropisetron in an experimental model of ulcerative colitis. Immunopharmacol. Immunotoxicol. 2016, 38, 432–440. [Google Scholar] [CrossRef]
- Yadav, P.N.; Liu, Z.; Rafi, M.M. A diarylheptanoid from lesser galangal (Alpinia officinarum) inhibits proinflammatory mediators via inhibition of mitogen-activated protein kinase, p44/42, and transcription factor nuclear factor-κB. J. Pharmacol. Exp. Ther. 2003, 305, 925–931. [Google Scholar] [CrossRef]
- Kaliyeva, S.; Simohina, N.; Yukhnevich, Y.; Myasnikova, Z.; Myasnikov, Y. Pharmacoeconomic assessment of biological therapy of ulcerative colitis. Value Health 2018, 21, S84. [Google Scholar] [CrossRef]
- Gupta, M.; Mishra, V.; Gulati, M.; Kapoor, B.; Kaur, A.; Gupta, R.; Tambuwala, M.M. Natural compounds as safe therapeutic options for ulcerative colitis. Inflammopharmacology 2022, 30, 397–434. [Google Scholar]
- Zhang, S.Z.; Zhao, X.H.; Zhang, D.C. Cellular and molecular immunopathogenesis of ulcerative colitis. Cell. Mol. Immunol. 2006, 3, 35–40. [Google Scholar]
- Lin, G.; Li, M.; Xu, N.; Wu, X.; Liu, J.; Wu, Y.; Zhang, Q.; Cai, J.; Gao, C.; Su, Z. Anti-inflammatory effects of Heritiera littoralis fruits on dextran sulfate sodium-(DSS-) induced ulcerative colitis in mice by regulating gut microbiota and suppressing NF-κB pathway. BioMed Res. Int. 2020, 2020, 8893621. [Google Scholar] [CrossRef]
- Zhang, Z.; Shen, P.; Xie, W.; Cao, H.; Liu, J.; Cao, Y.; Zhang, N. Pingwei San ameliorates dextran sulfate sodium-induced chronic colitis in mice. J. Ethnopharmacol. 2019, 236, 91–99. [Google Scholar] [CrossRef]
- Babalola, W.O.; Ofusori, D.A.; Awoniran, P.; Falana, B.A. Aloe vera gel attenuates acetic acid-induced ulcerative colitis in adult male Wistar rats. Toxicol. Rep. 2022, 9, 640–646. [Google Scholar] [CrossRef]
- Bahrami, G.; Malekshahi, H.; Miraghaee, S.; Madani, H.; Babaei, A.; Mohammadi, B.; Hatami, R. Protective and therapeutic effects of Aloe vera gel on ulcerative colitis induced by acetic acid in rats. Clin. Nutr. Res. 2020, 9, 223–234. [Google Scholar]
- Hassanshahi, N.; Masoumi, S.J.; Mehrabani, D.; Hashemi, S.S.; Zare, M. The healing effect of Aloe vera gel on acetic acid-induced ulcerative colitis in rat. Middle East J. Dig. Dis. 2020, 12, 154–161. [Google Scholar]
- Langmead, L.; Feakins, R.M.; Goldthorpe, S.; Holt, H.; Tsironi, E.; De Silva, A.; Jewell, D.P.; Rampton, D.S. Randomized, double-blind, placebo-controlled trial of oral Aloe vera gel for active ulcerative colitis. Aliment. Pharmacol. Ther. 2004, 19, 739–747. [Google Scholar]
- Radha, M.H.; Laxmipriya, N.P. Evaluation of biological properties and clinical effectiveness of Aloe vera: A systematic review. J. Tradit. Complement. Med. 2015, 5, 21–26. [Google Scholar]
- Korkina, L.; Suprun, M.; Petrova, A.; Mikhal’Chik, E.; Luci, A.; Luca, C.D. The protective and healing effects of a natural antioxidant formulation based on ubiquinol and Aloe vera against dextran sulfate-induced ulcerative colitis in rats. Biofactors 2003, 18, 255–264. [Google Scholar] [CrossRef]
- Naini, M.A.; Zargari-Samadnejad, A.; Mehrvarz, S.; Tanideh, R.; Ghorbani, M.; Dehghanian, A.; Hasanzarrini, M.; Banaee, F.; Hosseinabadi, O.K.; Iraji, A. Anti-inflammatory, antioxidant, and healing-promoting effects of Aloe vera extract in the experimental colitis in rats. Evid. Based. Complement. Altern. Med. 2021, 2021, 9945244. [Google Scholar]
- Choi, S.M.; Supeno, D.; Byun, J.Y.; Kwon, S.H.; Chung, S.W.; Kwon, S.G.; Park, J.M.; Kim, J.S.; Kwon, D.Y.; Choi, W.S. Chemical characteristics of Aloe vera and Aloe saponaria in Ulsan Korea. Int. J. Bio-Sci. Bio-Technol. 2016, 8, 109–118. [Google Scholar] [CrossRef]
- Shi, G.; Jiang, H.; Feng, J.; Zheng, X.; Zhang, D.; Jiang, C.; Zhang, J. Aloe vera mitigates dextran sulfate sodium-induced rat ulcerative colitis by potentiating colon mucus barrier. J. Ethnopharmacol. 2021, 279, 114108. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT-Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Price, M.L.; Hagerman, A.E.; Butler, L.G. Tannin content of cowpeas, chickpeas, pigeon peas, and mung beans. J. Food Chem. 1980, 28, 459–461. [Google Scholar] [CrossRef]
- Hassan, S.M.; Al Aqil, A.A.; Attimarad, M. Determination of crude saponin and total flavonoids content in guar meal. Adv. Med. Plant Res. 2013, 1, 24–28. [Google Scholar]
- Lee, S.J.; Kim, J.E.; Choi, Y.J.; Gong, J.E.; Park, S.H.; Douangdeuane, B.; Souliya, O.; Park, J.M.; Lee, H.S.; Kim, B.H.; et al. Therapeutic effects of Dipterocarpus tuberculatus with high antioxidative activity against UV-induced photoaging of NHDF cells and nude mice. Antioxidants 2021, 10, 791. [Google Scholar]
- Kitajima, S.; Takuma, S.; Morimoto, M. Histological analysis of murine colitis induced by dextran sulfate sodium of different molecular weights. Exp. Anim. 2000, 49, 9–15. [Google Scholar] [CrossRef]
- Xia, B.; Deng, C.S.; Chen, D.J.; Zhou, Y.; Xiao, J.Q. Role of copper zinc superoxide dismutase in the short-term treatment of acetic acid-induced colitis in rats. Acta Gastroenterol. Latinoam. 1996, 26, 227–230. [Google Scholar]
- Zea-Iriarte, W.L.; Makiyama, K.; Goto, S.; Murase, K.; Urata, Y.; Sekine, I.; Hara, K.; Kondo, T. Impairment of antioxidants in colonic epithelial cells isolated from trinitrobenzene sulphonic acid-induced colitis rats protective effect of rebamipide. Scand. J. Gastroenterol. 1996, 31, 985–992. [Google Scholar] [CrossRef]
- Kihara, N.; de la Fuente, S.G.; Fujino, K.; Takahashi, T.; Pappas, T.N.; Mantyh, C.R. Vanilloid receptor-1 containing primary sensory neurons mediate dextran sulphate sodium induced colitis in rats. Gut 2003, 52, 713–719. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Andujar, I.; Recio, M.C.; Giner, R.M.; Cienfuegos-Jovellanos, E.; Laghi, S.; Muguerza, B.; Rios, J.L. Inhibition of ulcerative colitis in mice after oral administration of a polyphenol-enriched cocoa extract is mediated by the inhibition of STAT1 and STAT3 phosphorylation in colon cells. Food Chem. 2011, 59, 6474–6483. [Google Scholar]
- Porter, R.J.; Kalla, R.; Ho, G.T. Ulcerative colitis: Recent advances in the understanding of disease pathogenesis. F1000Research 2020, 9, 294. [Google Scholar]
- Sninsky, C.A. New research in ulcerative colitis: Optimizing 5-ASA administration for efficacy and adherence. Gastroenterol. Hepatol. 2010, 6, 4. [Google Scholar]
- Lee, D.; Albenberg, L.; Compher, C.; Baldassano, R.; Piccoli, D.; Lewis, J.D.; Wu, G.D. Diet in the pathogenesis and treatment of inflammatory bowel diseases. Gastroenterology 2015, 148, 1087–1106. [Google Scholar] [CrossRef]
- Peyrin-Biroulet, L.; Desreumaux, P.; Sandborn, W.J.; Colombel, J.F. Crohn’s disease: Beyond antagonists of tumor necrosis factor. Lancet 2008, 372, 67–81. [Google Scholar]
- Adams, S.M.; Close, E.D.; Shreenath, A.P. Ulcerative colitis: Rapid evidence review. Am. Fam. Physician 2022, 105, 406–411. [Google Scholar]
- Jump, R.L.; Levine, A.D. Mechanisms of natural tolerance in the intestine: Implications for inflammatory bowel disease. Inflamm. Bowel Dis. 2004, 10, 462–478. [Google Scholar]
- Neuman, M.G. Immune dysfunction in inflammatory bowel disease. Transl. Res. 2007, 149, 173–186. [Google Scholar] [CrossRef]
- Leon, F.; Smythies, L.E.; Smith, P.D.; Kelsall, B.L. Involvement of dendritic cells in the pathogenesis of inflammatory bowel disease. Adv. Exp. Med. Biol. 2006, 579, 117–132. [Google Scholar]
- Papadakis, K.A.; Targan, S.R. Role of cytokines in the pathogenesis of inflammatory bowel disease. Annu. Rev. Med. 2000, 51, 289. [Google Scholar]
- Sanchez-Muñoz, F.; Dominguez-Lopez, A.; Yamamoto-Furusho, J.K. Role of cytokines in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 4280. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | No | DSS | Statistical Difference | |||
|---|---|---|---|---|---|---|
| Vehicle | LAAS | HAAS | ||||
| Organ weights | Liver (g) | 1.39 ± 0.10 | 0.97 ± 0.03 | 0.85 ± 0.04 | 0.89 ± 0.06 | No significance |
| Kidney (g) | 0.36 ± 0.03 | 0.32 ± 0.02 | 0.32 ± 0.02 | 0.32 ± 0.04 | No significance | |
| Serum biochemical factor | ALT (U/L) | 30.4 ± 2.8 | 31.5 ± 8.1 | 31.75 ± 8.6 | 34.0 ± 5.2 | No significance |
| AST (U/L) | 47.0 ± 3.3 | 44.3 ± 4.0 | 48.0 ± 1.7 | 46.75 ± 1.1 | No significance | |
| ALP (g/mL) | 91.0 ± 2.8 | 90.0 ± 4.2 | 88.0 ± 7.0 | 88.5 ± 2.1 | No significance | |
| BUN (mg/dL) | 23.8 ± 4.0 | 26.9 ± 2.2 | 28.8 ± 4.1 | 29.7 ± 4.0 | No significance | |
| Crea (mg/dL) | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | No significance | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kweon, D.Y.; Song, H.J.; Kim, J.E.; Jin, Y.J.; Roh, Y.J.; Seol, A.; Park, J.M.; Lee, E.S.; Choi, W.S.; Hwang, D.Y. Therapeutic Effects of Aloe saponaria against Ulcerative Colitis Induced by Dextran Sulfate Sodium. Curr. Issues Mol. Biol. 2023, 45, 1483-1499. https://doi.org/10.3390/cimb45020096
Kweon DY, Song HJ, Kim JE, Jin YJ, Roh YJ, Seol A, Park JM, Lee ES, Choi WS, Hwang DY. Therapeutic Effects of Aloe saponaria against Ulcerative Colitis Induced by Dextran Sulfate Sodium. Current Issues in Molecular Biology. 2023; 45(2):1483-1499. https://doi.org/10.3390/cimb45020096
Chicago/Turabian StyleKweon, Do Yeong, Hee Jin Song, Ji Eun Kim, You Jeong Jin, Yu Jeong Roh, Ayun Seol, Ju Min Park, Eun Suk Lee, Won Sik Choi, and Dae Youn Hwang. 2023. "Therapeutic Effects of Aloe saponaria against Ulcerative Colitis Induced by Dextran Sulfate Sodium" Current Issues in Molecular Biology 45, no. 2: 1483-1499. https://doi.org/10.3390/cimb45020096
APA StyleKweon, D. Y., Song, H. J., Kim, J. E., Jin, Y. J., Roh, Y. J., Seol, A., Park, J. M., Lee, E. S., Choi, W. S., & Hwang, D. Y. (2023). Therapeutic Effects of Aloe saponaria against Ulcerative Colitis Induced by Dextran Sulfate Sodium. Current Issues in Molecular Biology, 45(2), 1483-1499. https://doi.org/10.3390/cimb45020096