

Crocin Attenuates NLRP3 Inflammasome Activation by Inhibiting Mitochondrial Reactive Oxygen Species and Ameliorates Monosodium Urate-Induced Mouse Peritonitis
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of MSU Crystals
2.3. Isolation of Mouse Peritoneal Macrophages
2.4. Cell Culture and Treatment
2.5. Measurement of IL-1β and IL-18 Concentrations
2.6. Protein Precipitation of Cell Culture Supernatants for IL-1β and Caspase-1
2.7. Western Blot Analysis
2.8. LDH Release Assay
2.9. Cell Viability Assay
2.10. ASC Oligomerization Assay
2.11. Immunofluorescence Microscopy
2.12. Measurement of mtROS
2.13. MSU-Induced Murine Peritonitis
2.14. Statistical Analysis
3. Results
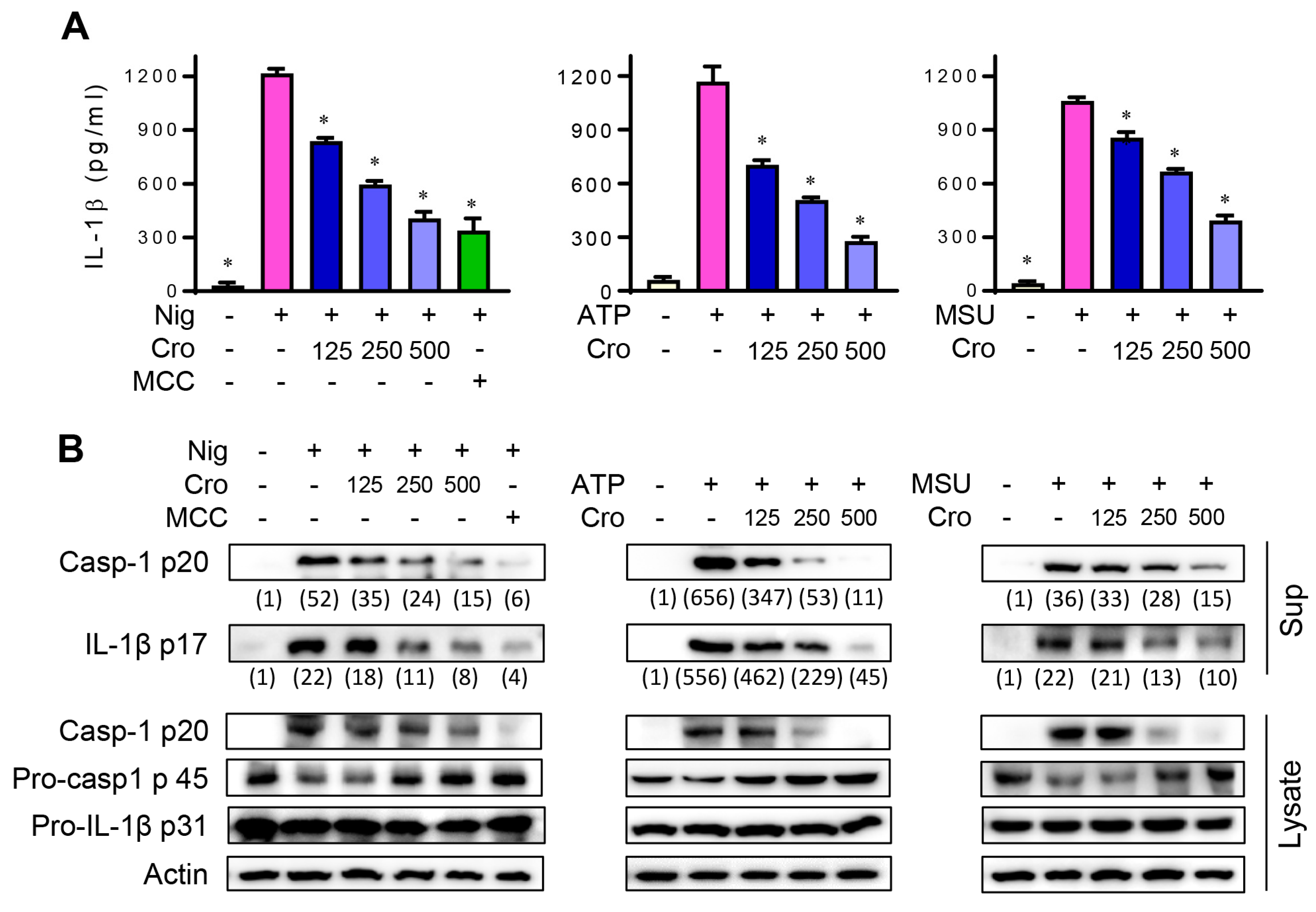
3.1. Crocin Suppresses NLRP3 Inflammasome Activation in Nigericin, ATP, or MSU-Stimulated Macrophages
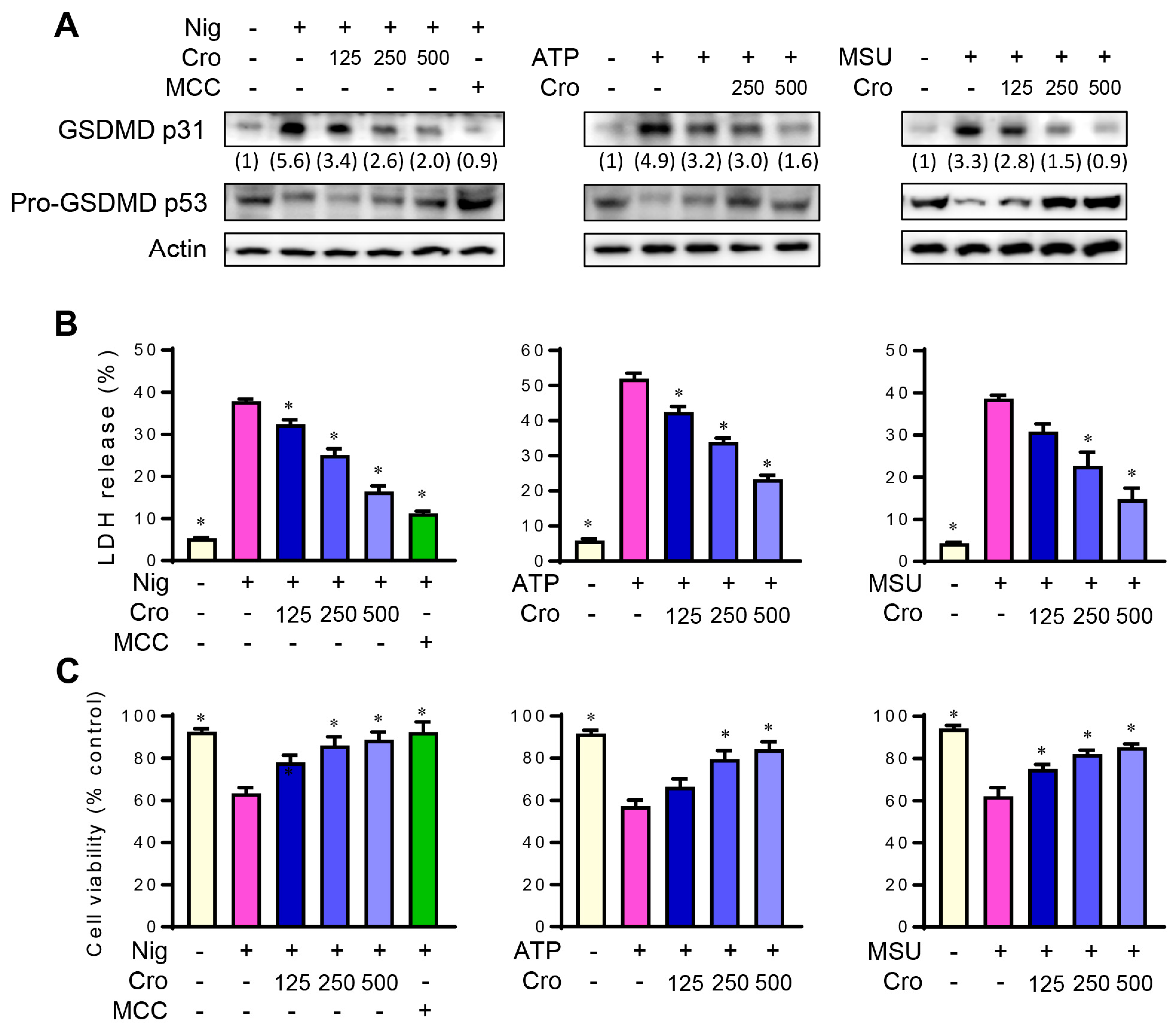
3.2. Crocin Inhibits NLRP3 Inflammasome-Induced GSDMD Cleavage and Pyroptosis
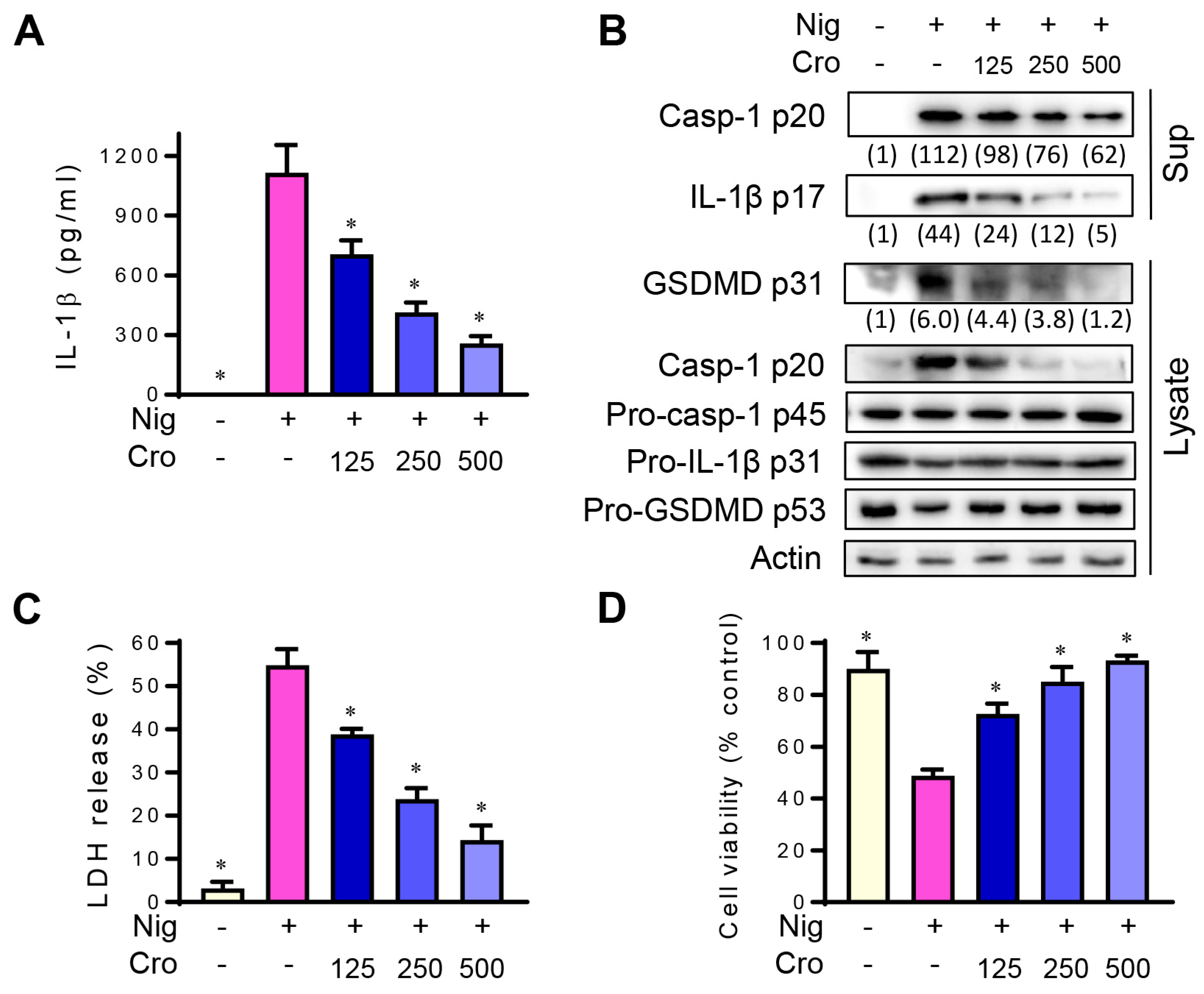
3.3. Crocin Inhibits NLRP3 Inflammasome in Mouse Peritoneal Macrophages
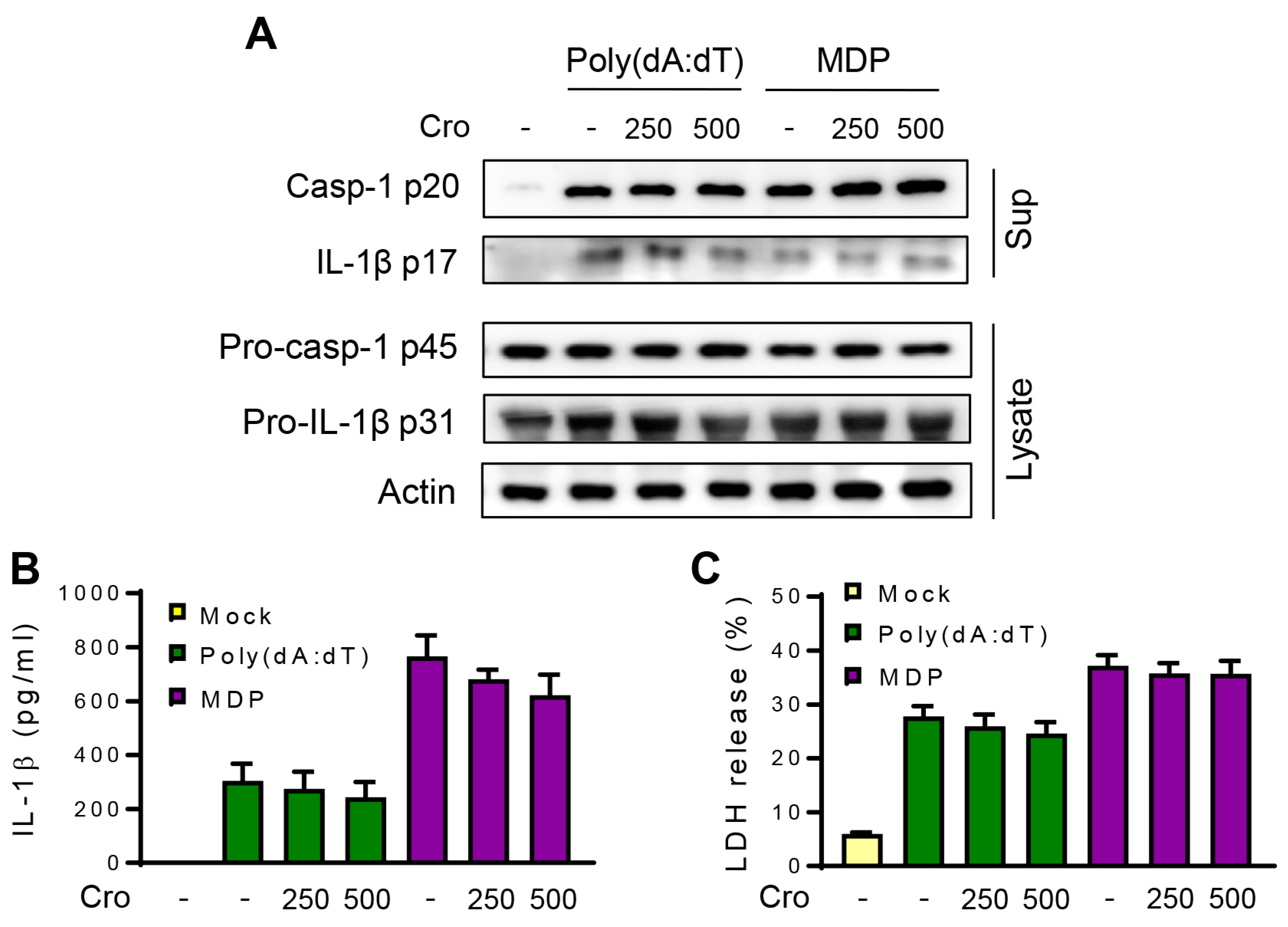
3.4. Crocin Does Not Affect NLRP1 or AIM2 Inflammasome
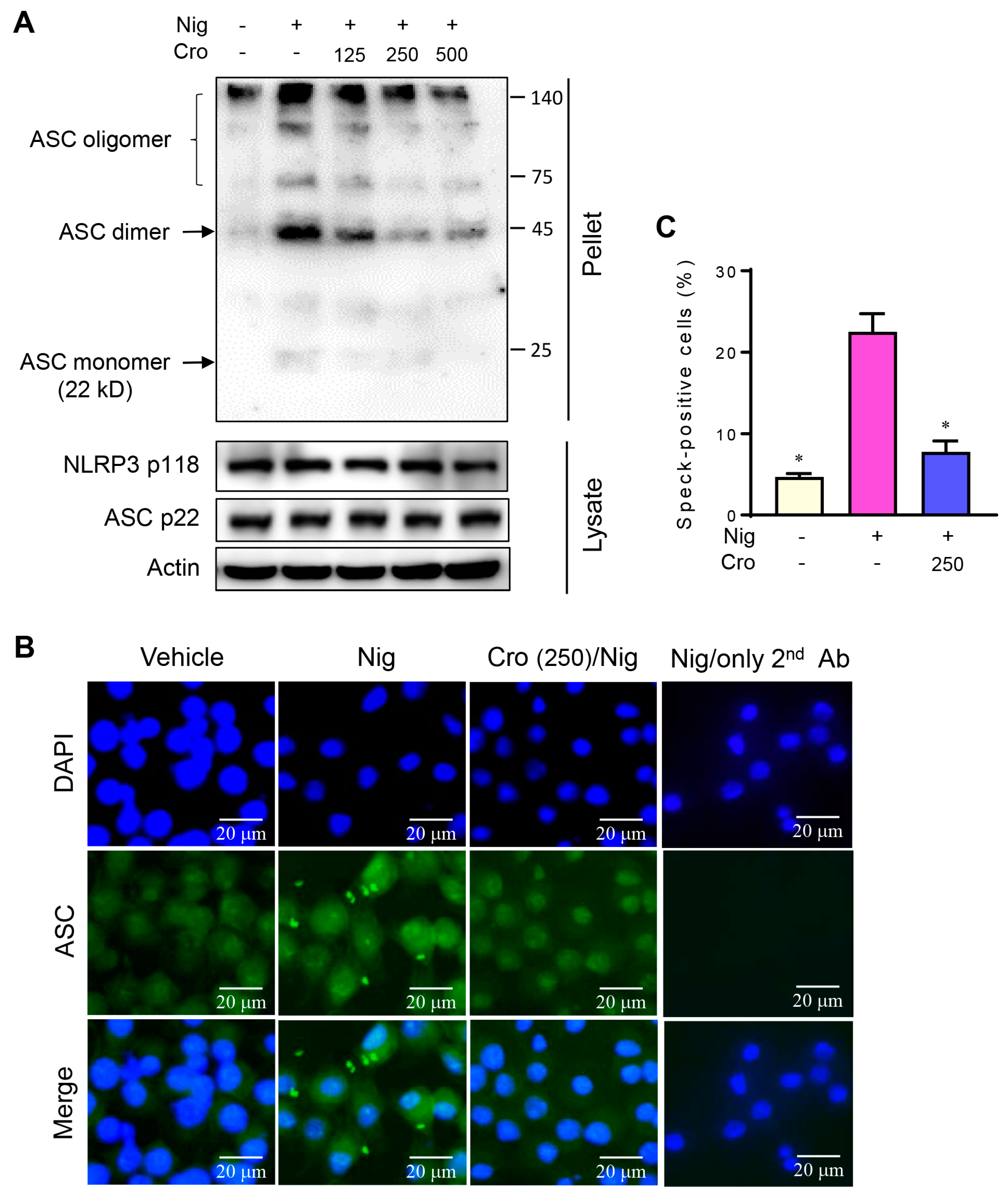
3.5. Crocin Suppresses ASC Oligomerization and ASC Speck Formation
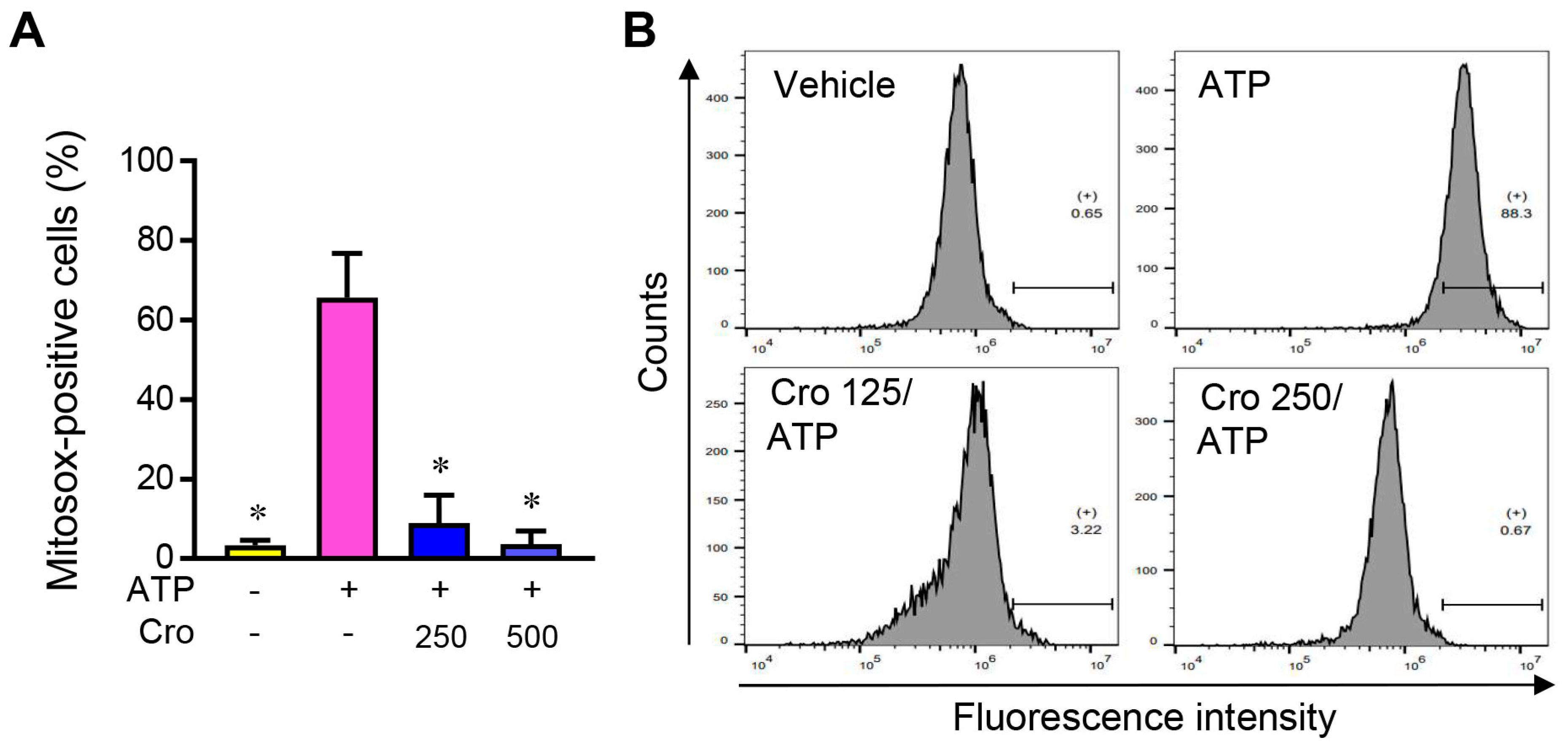
3.6. Crocin Suppresses mtROS Production
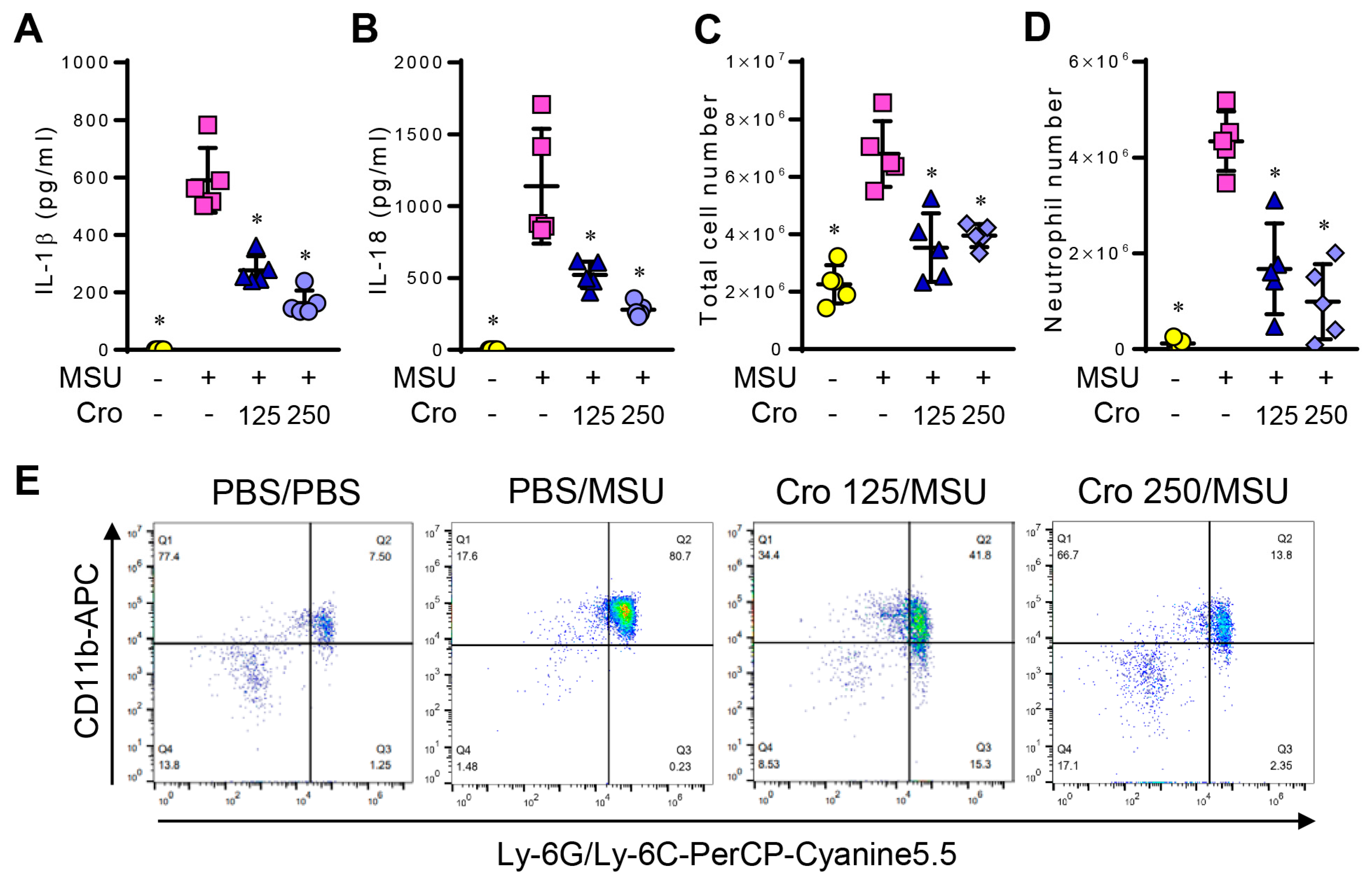
3.7. Crocin Inhibits IL-1β and IL-18 Production and Recruitment of Neutrophils in MSU-Induced Peritonitis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Lan, Z.; Xin, Z.; He, C.; Guo, Z.; Xia, X.; Hu, T. Emerging insights into molecular mechanisms underlying pyroptosis and functions of inflammasomes in diseases. J. Cell. Physiol. 2019, 235, 3207–3221. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An overview of mechanisms of activation and regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Xu, A.; Lv, J.; Zhang, Q.; Ran, Y.; Wei, C.; Wu, J. Development of small molecule inhibitors targeting NLRP3 inflammasome pathway for inflammatory diseases. Eur. J. Med. Chem. 2020, 185, 111822. [Google Scholar] [CrossRef]
- Pellegrini, C.; Fornai, M.; Antonioli, L.; Blandizzi, C.; Calderone, V. Phytochemicals as novel therapeutic strategies for NLRP3 inflammasome-related neurological, metabolic, and inflammatory diseases. Int. J. Mol. Sci. 2019, 20, 2876. [Google Scholar] [CrossRef] [Green Version]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and natural carotenoids: Biochemical activities and anti-tumor effects. Biochim. Biophys. Acta 2014, 1845, 20–30. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.F.; Li, F.; Zhang, H.Y. Fructus Gardenia (Gardenia jasminoides J. Ellis) phytochemistry, pharmacology of cardiovascular, and safety with the perspective of new drugs development. J. Asian Nat. Prod. Res. 2013, 15, 94–110. [Google Scholar] [CrossRef]
- Park, E.H.; Joo, M.H.; Kim, S.H.; Lim, C.J. Antiangiogenic activity of Gardenia jasminoides fruit. Phytother. Res. 2003, 17, 961–962. [Google Scholar] [CrossRef]
- Chang, K.H.; Chen, W.L.; Wu, Y.R.; Lin, T.H.; Wu, Y.C.; Chao, C.Y.; Lin, J.Y.; Lee, L.C.; Chen, Y.C.; Lee-Chen, G.J.; et al. Aqueous extract of Gardenia jasminoides targeting oxidative stress to reduce polyQ aggregation in cell models of spinocerebellar ataxia 3. Neuropharmacology 2014, 81, 166–175. [Google Scholar] [CrossRef]
- Assimopoulou, A.N.; Sinakos, Z.; Papageorgiou, V.P. Radical scavenging activity of Crocus sativus L. extract and its bioactive constituents. Phytother. Res. 2005, 19, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Finley, J.W.; Gao, S. A Perspective on Crocus sativus L. (Saffron) constituent Crocin: A potent water-soluble antioxidant and potential therapy for Alzheimer’s disease. J. Agric. Food Chem. 2017, 65, 1005–1020. [Google Scholar] [CrossRef]
- Deslauriers, A.M.; Afkhami-Goli, A.; Paul, A.M.; Bhat, R.K.; Acharjee, S.; Ellestad, K.K.; Noorbakhsh, F.; Michalak, M.; Power, C. Neuroinflammation and endoplasmic reticulum stress are coregulated by crocin to prevent demyelination and neurodegeneration. J. Immunol. 2011, 187, 4788–4799. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Park, G.Y.; Bang, S.Y.; Park, S.Y.; Bae, S.K.; Kim, Y. Crocin suppresses LPS-stimulated expression of inducible nitric oxide synthase by upregulation of heme oxygenase-1 via calcium/calmodulin-dependent protein kinase 4. Mediators Inflamm. 2014, 2014, 728709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano, J.; Alonso, G.L.; Coca-Prados, M.; Fernandez, J.A. Crocin, safranal and picrocrocin from saffron (Crocus sativus L.) inhibit the growth of human cancer cells in vitro. Cancer Lett. 1996, 100, 23–30. [Google Scholar] [CrossRef] [PubMed]
- He, S.Y.; Qian, Z.Y.; Tang, F.T.; Wen, N.; Xu, G.L.; Sheng, L. Effect of crocin on experimental atherosclerosis in quails and its mechanisms. Life Sci. 2005, 77, 907–921. [Google Scholar] [CrossRef]
- Song, R.; Han, S.; Gao, H.; Jiang, H.; Li, X. Crocin alleviates cognitive impairment associated with atherosclerosis via improving neuroinflammation in LDLR−/− mice fed a high-fat/cholesterol diet. Phytother. Res. 2022, 36, 1284–1296. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ali Redha, A.; Snoeck, E.R.; Singh, S.; Simal-Gandara, J.; Ibrahim, S.A.; Jafari, S.M. Anti-depressant properties of crocin molecules in saffron. Molecules 2022, 27, 2076. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, J.; Shen, Y.; Pan, J.; Wang, C.; Cheng, L. Protective effect of crocin on immune checkpoint inhibitors-related myocarditis through inhibiting NLRP3 mediated pyroptosis in cardiomyocytes via NF-κB pathway. J. Inflamm. Res. 2022, 15, 1653–1666. [Google Scholar] [CrossRef]
- Lin, J.K.; Wang, C.J. Protection of crocin dyes on the acute hepatic damage induced by aflatoxin B1 and dimethylnitrosamine in rats. Carcinogenesis 1986, 7, 595–599. [Google Scholar] [CrossRef]
- Lv, B.; Chen, T.; Xu, Z.; Huo, F.; Wei, Y.; Yang, X. Crocin protects retinal ganglion cells against H2O2-induced damage through the mitochondrial pathway and activation of NF-κB. Int. J. Mol. Med. 2016, 37, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Previn, R.; Lu, L.; Liao, R.F.; Jin, Y.; Wang, R.K. Crocin, a natural product attenuates lipopolysaccharide-induced anxiety and depressive-like behaviors through suppressing NF-κB and NLRP3 signaling pathway. Brain Res. Bull. 2018, 142, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Petrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, S.Y.; Kim, J.-H.; Kim, H.-Y.; Lee, Y.J.; Park, S.Y.; Lee, S.J.; Kim, Y. Achyranthes japonica exhibits anti-inflammatory effect via NF-κB suppression and HO-1 induction in macrophages. J. Ethnopharmacol. 2012, 144, 109–117. [Google Scholar] [CrossRef]
- Fic, E.; Kedracka-Krok, S.; Jankowska, U.; Pirog, A.; Dziedzicka-Wasylewska, M. Comparison of protein precipitation methods for various rat brain structures prior to proteomic analysis. Electrophoresis 2010, 31, 3573–3579. [Google Scholar] [CrossRef]
- Lugrin, J.; Martinon, F. Detection of ASC oligomerization by western blotting. Bio-Protoc. 2017, 7, e2292. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, K. Inflammasome-associated cell death: Pyroptosis, apoptosis, and physiological implications. Microbiol. Immunol. 2020, 64, 252–269. [Google Scholar] [CrossRef]
- Duan, L.; Chen, J.; Razavi, M.; Wei, Y.; Tao, Y.; Rao, X.; Zhong, J. Alpha2B-adrenergic receptor regulates neutrophil recruitment in MSU-induced peritoneal inflammation. Front. Immunol. 2019, 10, 501. [Google Scholar] [CrossRef]
- Getting, S.J.; Di Filippo, C.; Lam, C.W.; Rossi, F.; D’Amico, M. Investigation into the potential anti-inflammatory effects of endothelin antagonists in a murine model of experimental monosodium urate peritonitis. J. Pharmacol. Exp. Ther. 2004, 310, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Jhang, J.-J.; Lu, C.-C.; Yen, G.-C. Epigallocatechin gallate inhibits urate crystals-induced peritoneal inflammation in C57BL/6 mice. Mol. Nutr. Food Res. 2016, 60, 2297–2303. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, M.; Liu, Q. Crocin alleviates the inflammation and oxidative stress responses associated with diabetic nephropathy in rats via NLRP3 inflammasomes. Life Sci. 2021, 278, 119542. [Google Scholar] [CrossRef]
- Souza, J.G.; Starobinas, N.; Ibañez, O.C.M. Unknown/enigmatic functions of extracellular ASC. Immunology 2021, 163, 377–388. [Google Scholar] [CrossRef]
- Franklin, B.S.; Latz, E.; Schmidt, F.I. The intra- and extracellular functions of ASC specks. Immunol. Rev. 2018, 281, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Franklin, B.S.; Bossaller, L.; De Nardo, D.; Ratter, J.M.; Stutz, A.; Engels, G.; Brenker, C.; Nordhoff, M.; Mirandola, S.R.; Al-Amoudi, A.; et al. The adaptor ASC has extracellular and ’prionoid’ activities that propagate inflammation. Nat. Immunol. 2014, 15, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Baroja-Mazo, A.; Martín-Sánchez, F.; Gomez, A.I.; Martínez, C.M.; Amores-Iniesta, J.; Compan, V.; Barberà-Cremades, M.; Yagüe, J.; Ruiz-Ortiz, E.; Antón, J.; et al. The NLRP3 inflammasome is released as a particulate danger signal that amplifies the inflammatory response. Nat. Immunol. 2014, 15, 738–748. [Google Scholar] [CrossRef]
- Li, X.; Jiang, C.; Zhu, W. Crocin reduces the inflammation response in rheumatoid arthritis. Biosc. Biotechnol. Biochem. 2017, 81, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Liu, G.; Yang, L. Crocin improves cognitive behavior in rats with Alzheimer’s disease by regulating endoplasmic reticulum stress and apoptosis. Biomed. Res. Int. 2019, 2019, 9454913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadipour, M.; Meftahi, G.H.; Afarinesh, M.R.; Jahromi, G.P.; Hatef, B. Crocin attenuates the granular cells damages on the dentate gyrus and pyramidal neurons in the CA3 regions of the hippocampus and frontal cortex in the rat model of Alzheimer’s disease. J. Chem. Neuroanat. 2021, 113, 101837. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Lu, L.; Wang, Q.; Liu, H.; Xue, W.; Zhou, T.; Xu, L.; Wang, K.; Wu, D.; Wei, F.; et al. Crocin reverses depression-like behavior in Parkinson disease mice via VTA-mPFC Pathway. Mol. Neurobiol. 2020, 57, 3158–3170. [Google Scholar] [CrossRef] [PubMed]
- Holley, C.L.; Schroder, K. The rOX-stars of inflammation: Links between the inflammasome and mitochondrial meltdown. Clin. Transl. Immunol. 2020, 9, e1109. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhang, D.; Hu, D.; Zhou, X.; Zhou, Y. The role of mitochondria in NLRP3 inflammasome activation. Mol. Immunol. 2018, 103, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhai, Y.; Liang, S.; Mori, Y.; Han, R.; Sutterwala, F.S.; Qiao, L. TRPM2 links oxidative stress to NLRP3 inflammasome activation. Nat. Commun. 2013, 4, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, H.; Yang, Z.; Ou, Y.; Liang, S.; Deng, W.; Chen, H.; Zhang, C.; Hua, L.; Hu, W.; Sun, P. Tanshinones inhibit NLRP3 inflammasome activation by alleviating mitochondrial damage to protect against septic and gouty inflammation. Int. Immunopharmacol. 2021, 97, 107819. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, X.; Zong, B.; Yuan, H.; Wang, Z.; Wei, Y.; Wang, X.; Liu, G.; Zhang, J.; Li, S.; et al. Gypenosides improve diabetic cardiomyopathy by inhibiting ROS-mediated NLRP3 inflammasome activation. J. Cell. Mol. Med. 2018, 22, 4437–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelblanco, M.; Lugrin, J.; Ehirchiou, D.; Nasi, S.; Ishii, I.; So, A.; Martinon, F.; Busso, N. Hydrogen sulfide inhibits NLRP3 inflammasome activation and reduces cytokine production both in vitro and in a mouse model of inflammation. J. Biol. Chem. 2018, 293, 2546–2557. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.; Deng, S.; Zhang, L.; Chen, S.; Lu, Q.; Peng, H. Crocin protects against dexamethasone-induced osteoblast apoptosis by inhibiting the ROS/Ca2+-mediated mitochondrial pathway. Mol. Med. Rep. 2019, 20, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Suh, K.S.; Chon, S.; Jung, W.W.; Choi, E.M. Crocin attenuates methylglyoxal-induced osteoclast dysfunction by regulating glyoxalase, oxidative stress, and mitochondrial function. Food Chem. Toxicol. 2019, 124, 367–373. [Google Scholar] [CrossRef]
- Dagvadorj, J.; Mikulska-Ruminska, K.; Tumurkhuu, G.; Ratsimandresy, R.A.; Carriere, J.; Andres, A.M.; Marek-Iannucci, S.; Song, Y.; Chen, S.; Lane, M.; et al. Recruitment of pro-IL-1α to mitochondrial cardiolipin, via shared LC3 binding domain, inhibits mitophagy and drives maximal NLRP3 activation. Proc. Natl. Acad. Sci. USA 2021, 118, e2015632118. [Google Scholar] [CrossRef] [PubMed]
- Xian, H.; Watari, K.; Sanchez-Lopez, E.; Offenberger, J.; Onyuru, J.; Sampath, H.; Ying, W.; Hoffman, H.M.; Shadel, G.S.; Karin, M. Oxidized DNA fragments exit mitochondria via mPTP- and VDAC-dependent channels to activate NLRP3 inflammasome and interferon signaling. Immunity 2022, 55, 1370–1385.e1378. [Google Scholar] [CrossRef]
- Zhou, Y.; Tong, Z.; Jiang, S.; Zheng, W.; Zhao, J.; Zhou, X. The roles of endoplasmic reticulum in NLRP3 inflammasome activation. Cells 2020, 9, 1219. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangare, R.; Madhi, I.; Kim, J.-H.; Kim, Y. Crocin Attenuates NLRP3 Inflammasome Activation by Inhibiting Mitochondrial Reactive Oxygen Species and Ameliorates Monosodium Urate-Induced Mouse Peritonitis. Curr. Issues Mol. Biol. 2023, 45, 2090-2104. https://doi.org/10.3390/cimb45030134
Sangare R, Madhi I, Kim J-H, Kim Y. Crocin Attenuates NLRP3 Inflammasome Activation by Inhibiting Mitochondrial Reactive Oxygen Species and Ameliorates Monosodium Urate-Induced Mouse Peritonitis. Current Issues in Molecular Biology. 2023; 45(3):2090-2104. https://doi.org/10.3390/cimb45030134
Chicago/Turabian StyleSangare, Ruth, Iskander Madhi, Ji-Hee Kim, and YoungHee Kim. 2023. "Crocin Attenuates NLRP3 Inflammasome Activation by Inhibiting Mitochondrial Reactive Oxygen Species and Ameliorates Monosodium Urate-Induced Mouse Peritonitis" Current Issues in Molecular Biology 45, no. 3: 2090-2104. https://doi.org/10.3390/cimb45030134
APA StyleSangare, R., Madhi, I., Kim, J. -H., & Kim, Y. (2023). Crocin Attenuates NLRP3 Inflammasome Activation by Inhibiting Mitochondrial Reactive Oxygen Species and Ameliorates Monosodium Urate-Induced Mouse Peritonitis. Current Issues in Molecular Biology, 45(3), 2090-2104. https://doi.org/10.3390/cimb45030134