

Serratiopeptidase Attenuates Lipopolysaccharide-Induced Vascular Inflammation by Inhibiting the Expression of Monocyte Chemoattractant Protein-1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Method
2.1. Experimental Animals
2.2. Oxidative Stress Parameters in Aorta
2.3. Tumor Necrosis Factor-α and Interleukin in Aorta
2.4. Immunohistochemistry of Aortic Tissue
2.5. Histological Analysis
2.6. Statistical Analysis
3. Results
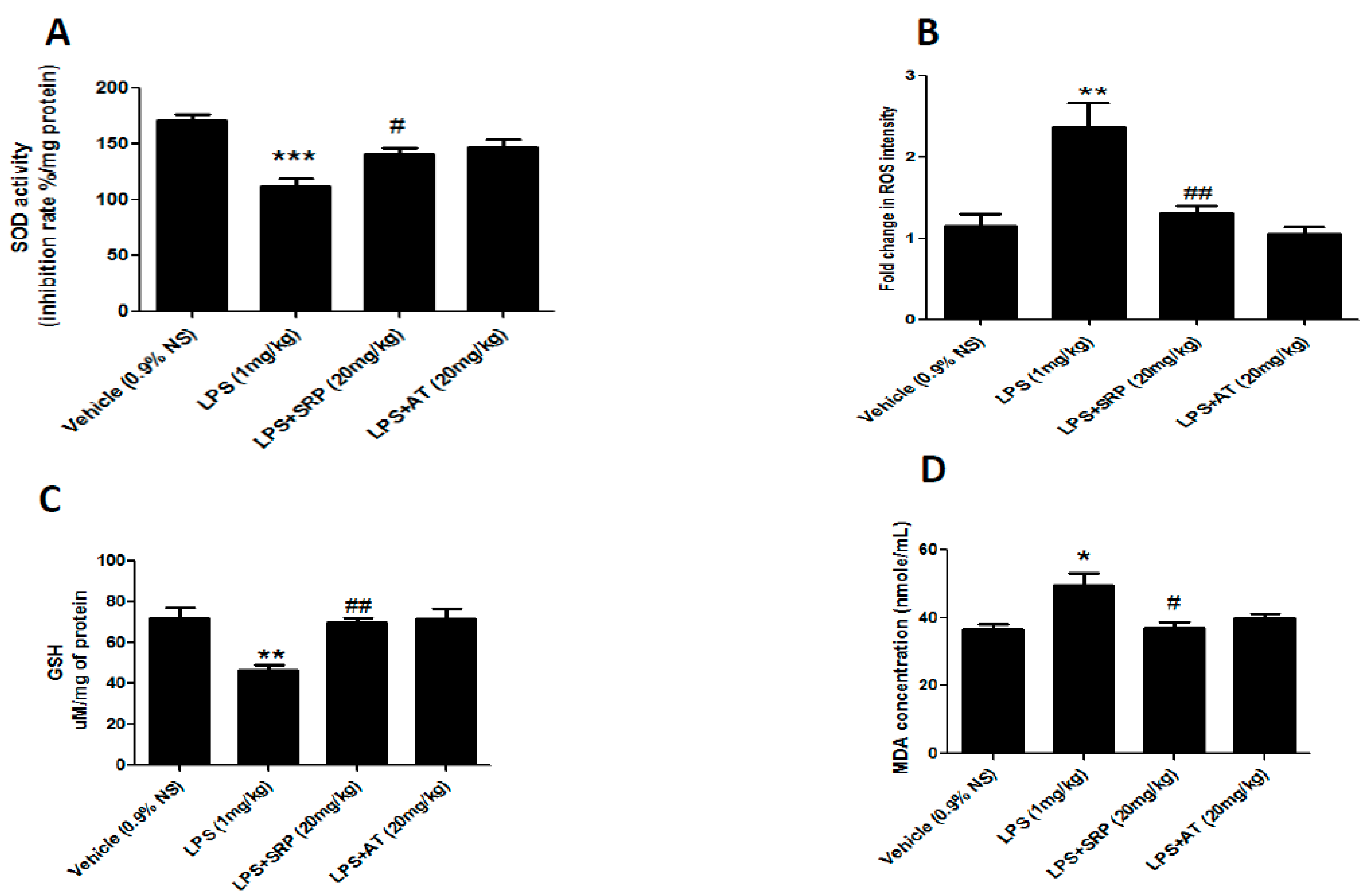
3.1. Effect of SRP against Oxidative Stress in Aorta
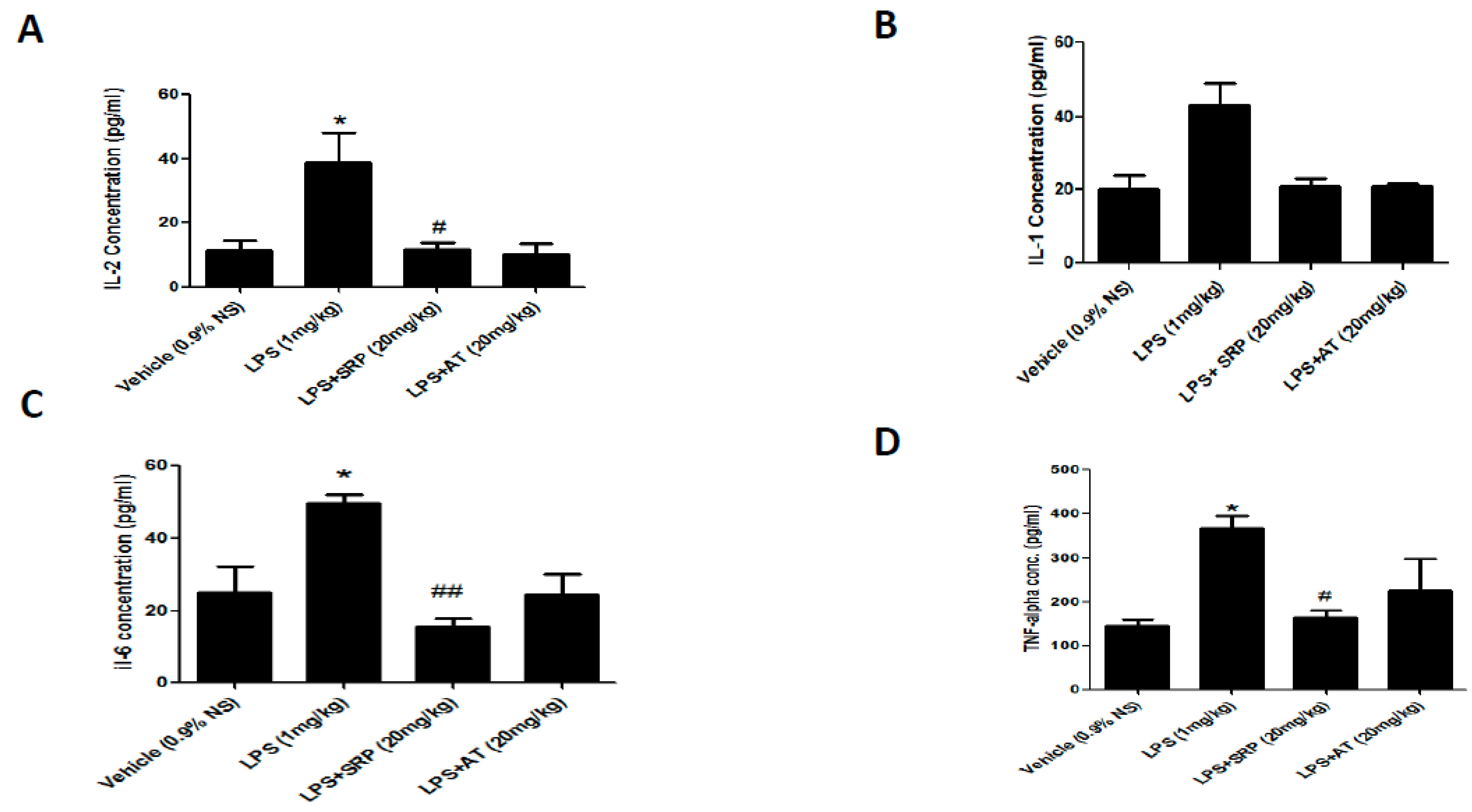
3.2. Effect on Saortic IL-2, IL-4, IL-6, and TNF-α Levels
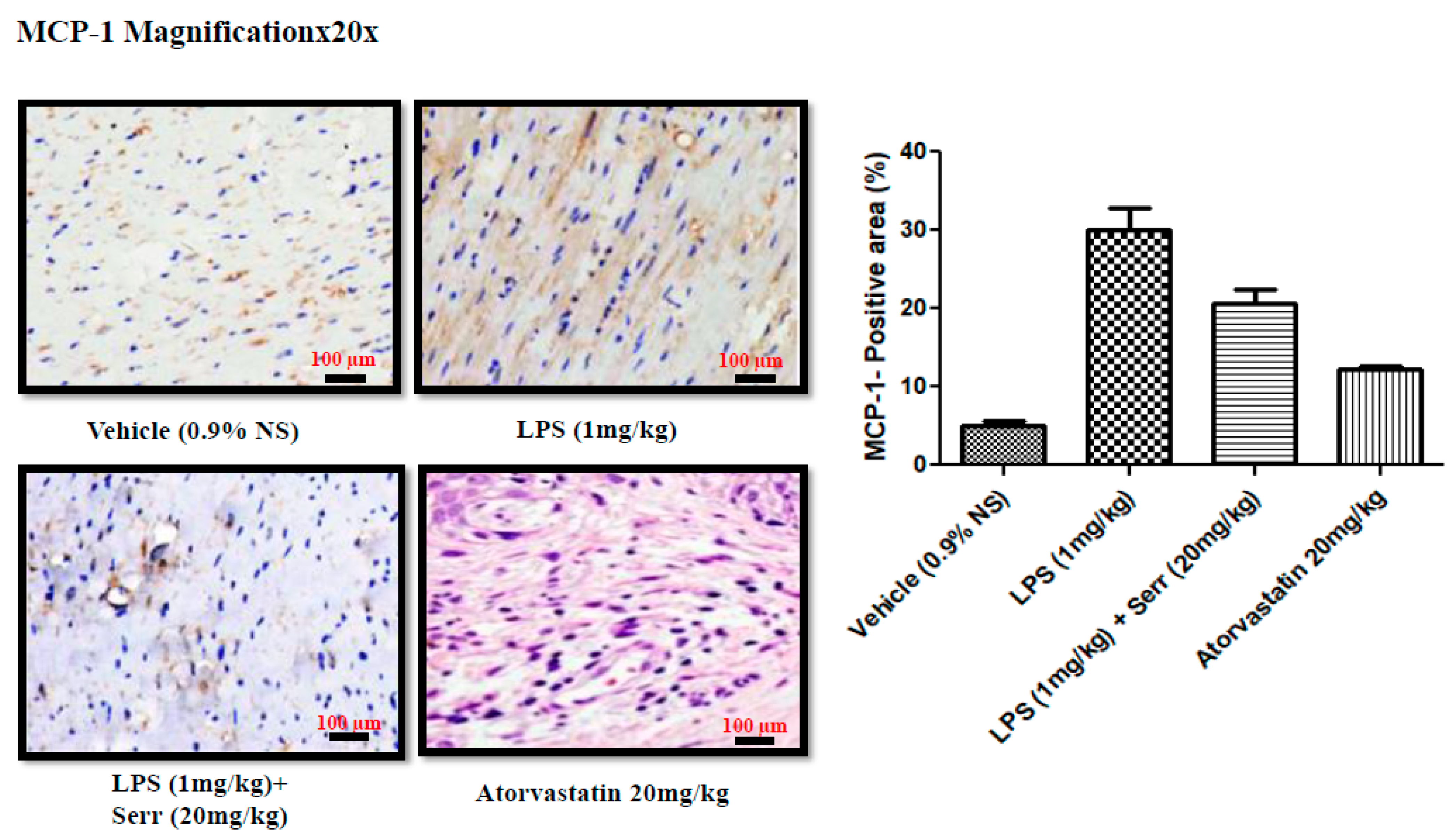
3.3. Effect on Production of MCP-1 in Aorta
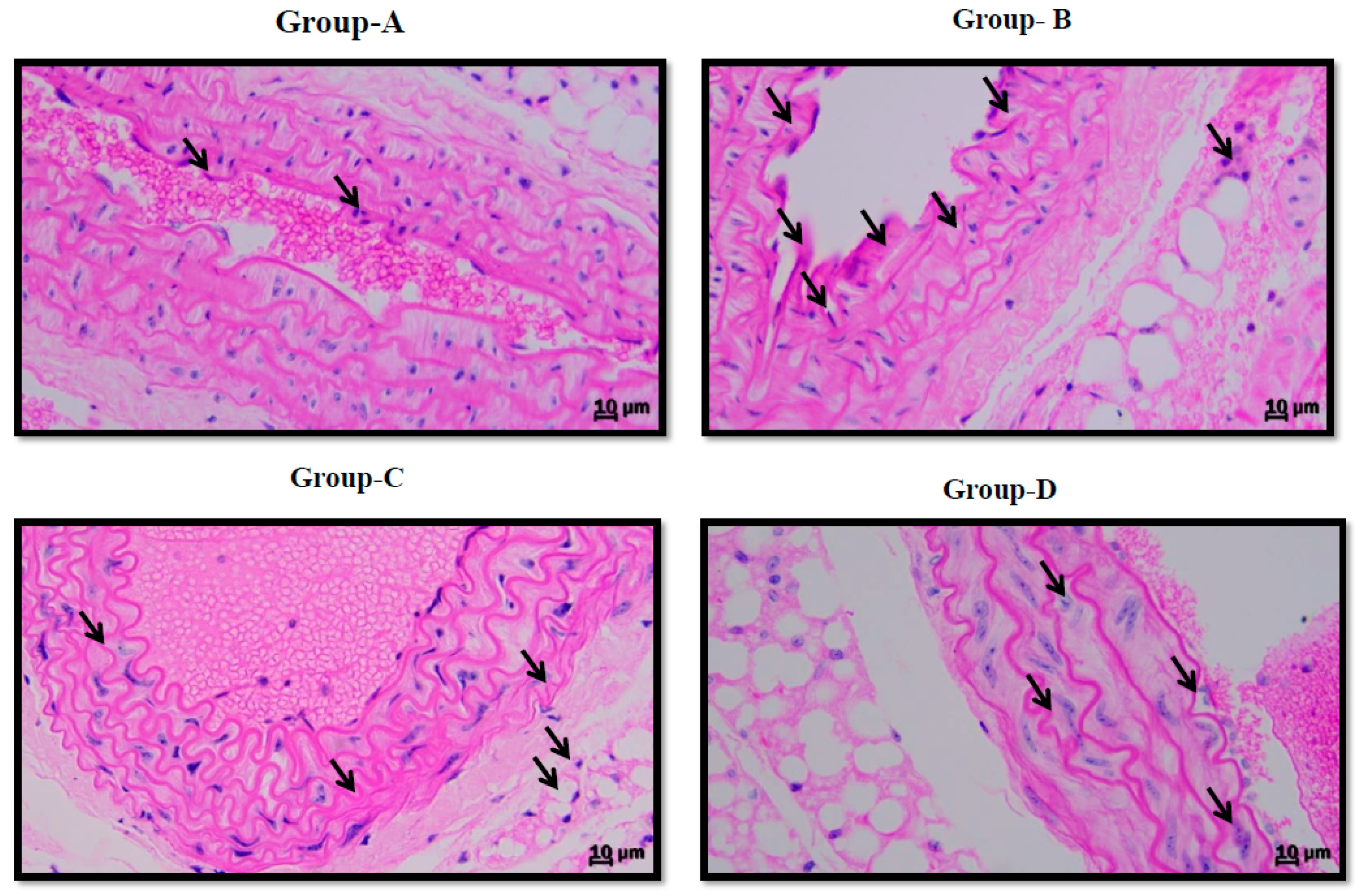
3.4. Effect on Vascular Endothelium in Aorta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Youness, R.A.; Assal, R.A.; Motaal, A.A.; Gad, M.Z. A novel role of sONE/NOS3/NO signaling cascade in mediating hydrogen sulphide bilateral effects on triple negative breast cancer progression. Nitric Oxide 2018, 80, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Ojha, S.; Patil, C.R. Animal Models of Inflammation for Screening of Anti-inflammatory Drugs: Implications for the Discovery and Development of Phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.J.; Lee, H.S.; Shin, J.S.; Lee, S.H.; Park, B.M.; Youn, Y.S.; Lee, S.K. Modulation of acute and chronic inflammatory processes by a traditional medicine preparation GCSB-5 both in vitro and in vivo animal models. J. Ethnopharmacol. 2010, 130, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Achaiah, G.; Sastry, G.N. Novel Targets for Antiinflammatory and Antiarthritic Agents. Curr. Pharm. Des. 2006, 12, 2437–2454. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhao, J.Y.; Zhao, Z.L.; Ye, J.; Li, S.F.; Fang, H.H.; Gu, M.N.; Hu, Y.W.; Qin, Z.S. Propofol Attenuates Lipopolysaccharide-Induced Monocyte Chemoattractant Protein-1 Production Through Enhancing apoM and foxa2 Expression in HepG2 Cells. Inflammation 2015, 38, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-B.; Yao, X.-X.; Xiu, J.-C.; Hu, Y.-W. MicroRNA-125b-5p attenuates lipopolysaccharide-induced monocyte chemoattractant protein-1 production by targeting inhibiting LACTB in THP-1 macrophages. Arch. Biochem. Biophys. 2016, 590, 64–71. [Google Scholar] [CrossRef]
- Anand, A.R.; Bradley, R.; Ganju, R.K. LPS-induced MCP-1 expression in human microvascular endothelial cells is mediated by the tyrosine kinase, Pyk2 via the p38 MAPK/NF-κB-dependent pathway. Mol. Immunol. 2009, 46, 962–968. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, F.; Finn, A.V.; Yazdani, S.K.; Nakano, M.; Kolodgie, F.D.; Virmani, R. The importance of the endothelium in atherothrombosis and coronary stenting. Nat. Rev. Cardiol. 2012, 9, 439–453. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr. Endothelial dysfunction, hemodynamic forces, and atherosclerosis. Thromb. Haemost. 1999, 82, 722–726. [Google Scholar] [CrossRef]
- Zhang, C. The role of inflammatory cytokines in endothelial dysfunction. Basic Res. Cardiol. 2008, 103, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Castellon, X.; Bogdanova, V. Chronic Inflammatory Diseases and Endothelial Dysfunction. Aging Dis. 2016, 7, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fosse, J.H.; Haraldsen, G.; Falk, K.; Edelmann, R. Endothelial Cells in Emerging Viral Infections. Front. Cardiovasc. Med. 2021, 8, 619690. [Google Scholar] [CrossRef] [PubMed]
- Sandoo, A.; Veldhuijzen van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Gautam, A.S. Upregulated LOX-1 Receptor: Key Player of the Pathogenesis of Atherosclerosis. Curr. Atheroscler. Rep. 2019, 21, 38. [Google Scholar] [CrossRef]
- Fusco, R.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Cordaro, M.; Crupi, R.; Impellizzeri, D.; Gomiero, C.; Cuzzocrea, S. Micro Composite Palmitoylethanolamide/Rutin Reduces Vascular Injury through Modulation of the Nrf2/HO− 1 and NF-kB Pathways. Curr. Med. Chem. 2021, 28, 6287–6302. [Google Scholar] [CrossRef]
- Daci, A.; Da Dalt, L.; Alaj, R.; Shurdhiqi, S.; Neziri, B.; Ferizi, R.; Norata, G.D.; Krasniqi, S. Rivaroxaban improves vascular response in LPS-induced acute inflammation in experimental models. PLoS ONE 2020, 15, e0240669. [Google Scholar] [CrossRef]
- D’Amico, R.; Monaco, F.; Siracusa, R.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Cuzzocrea, S.; Di Paola, R.; et al. Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. Int. J. Mol. Sci. 2021, 22, 11388. [Google Scholar] [CrossRef] [PubMed]
- Mangana, C.; Lorigo, M.; Cairrao, E. Implications of Endothelial Cell-Mediated Dysfunctions in Vasomotor Tone Regulation. Biol. 2021, 1, 231–251. [Google Scholar] [CrossRef]
- Singh, S. Herbal Approach for Management of Atherosclerosis: A Review. Curr. Atheroscler. Rep. 2019, 21, 12. [Google Scholar] [CrossRef]
- Sousa, J.; Diniz, C. Vascular sympathetic neurotransmission and endothelial dysfunction. In Endothelial Dysfunction—Old Concepts and New Challenges; IntechOpen: London, UK, 2018; pp. 327–344. [Google Scholar] [CrossRef] [Green Version]
- Badhe, R.V.; Nanda, R.K.; Kulkarni, M.B.; Bhujbal, M.N.; Patil, P.S.; Badhe, S.R. Media optimization studies for Serratiopeptidase production from Serratia marcescens ATCC 13880. Hindustan Antibiot. Bull. 2009, 51, 17–23. [Google Scholar]
- Bromke, B.J.; Hammel, J.M. Regulation of extracellular protease formation by Serratia marcescens. Can. J. Microbiol. 1979, 25, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Selan, L.; Papa, R.; Ermocida, A.; Cellini, A.; Ettorre, E.; Vrenna, G.; Campoccia, D.; Montanaro, L.; Arciola, C.R.; Artini, M. Serratiopeptidase reduces the invasion of osteoblasts by Staphylococcus aureus. Int. J. Immunopathol. Pharmacol. 2017, 30, 423–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadl, N.N.; Ahmed, H.H.; Booles, H.F.; Sayed, A.H. Serrapeptase and nattokinase intervention for relieving Alzheimer’s disease pathophysiology in rat model. Hum. Exp. Toxicol. 2013, 32, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Longhi, C.; Scoarughi, G.L.; Poggiali, F.; Cellini, A.; Carpentieri, A.; Seganti, L.; Pucci, P.; Amoresano, A.; Cocconcelli, P.S.; Artini, M.; et al. Protease treatment affects both invasion ability and biofilm formation in Listeria monocytogenes. Microb. Pathog. 2008, 45, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Mishra, P. Co-delivery of curcumin and serratiopeptidase in HeLa and MCF-7 cells through nanoparticles show improved anti-cancer activity. Mater. Sci. Eng. C 2018, 92, 673–684. [Google Scholar] [CrossRef]
- Metkar, S.K.; Girigoswami, A.; Murugesan, R.; Girigoswami, K. In vitro and in vivo insulin amyloid degradation mediated by Serratiopeptidase. Mater. Sci. Eng. C 2017, 70, 728–735. [Google Scholar] [CrossRef]
- Metkar, S.K.; Girigoswami, A.; Vijayashree, R.; Girigoswami, K. Attenuation of subcutaneous insulin induced amyloid mass in vivo using Lumbrokinase and Serratiopeptidase. Int. J. Biol. Macromol. 2020, 163, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef]
- van der Velde, A.R.; Meijers, W.C.; de Boer, R.A. Cardiovascular biomarkers: Translational aspects of hypertension, atherosclerosis, and heart failure in drug development. In Principles of Translational Science in Medicine; Elsevier: Amsterdam, The Netherlands, 2015; pp. 167–183. [Google Scholar]
- Singh, S.; Gautam, U.; Manvi, F.V. Protective Impact of Edaravone Against ZnO NPs-induced Oxidative Stress in the Human Neuroblastoma SH-SY5Y Cell Line. Cell. Mol. Neurobiol. 2020, 42, 1189–1210. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, A. Protective effect of edaravone on cyclophosphamide induced oxidative stress and neurotoxicity in rats. Curr. Drug Saf. 2019, 14, 209–216. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, J.; Morgan, S.; Liu, Z.; Dou, C.; Liu, B. Monocyte Chemoattractant Protein-1 (MCP-1) Regulates Macrophage Cytotoxicity in Abdominal Aortic Aneurysm. PLoS ONE 2014, 9, e92053. [Google Scholar] [CrossRef] [PubMed]
- Boudahna, L.; Benbrahim, Z.; Amaadour, L.; Mazouz, A.; Benhayoune, K.; Tahiri, Y.; Farih, M.H.; Amarti, A.; Arifi, S.; Mellas, N. Para testicular rhabdomyosarcoma in adults: Three case reports and review of literature. Pan Afr. Med. J. 2014, 19, 279. [Google Scholar] [CrossRef] [PubMed]
- Eelen, G.; De Zeeuw, P.; Simons, M.; Carmeliet, P. Endothelial Cell Metabolism in Normal and Diseased Vasculature. Circ. Res. 2015, 116, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Hu, T.; Hao, H.; Hill, M.A.; Xu, C.; Liu, Z. Inflammatory bowel disease and cardiovascular diseases: A concise review. Eur. Heart J. Open 2021, 2, oeab029. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M. The role of serratiopeptidase in the resolution of inflammation. Asian J. Pharm. Sci. 2017, 12, 209–215. [Google Scholar] [CrossRef]
- Al-Khateeb, T.; Nusair, Y. Effect of the proteolytic enzyme serrapeptase on swelling, pain and trismus after surgical extraction of mandibular third molars. Int. J. Oral Maxillofac. Surg. 2008, 37, 264–268. [Google Scholar] [CrossRef]
- Cines, D.B.; Pollak, E.S.; Buck, C.A.; Loscalzo, J.; Zimmerman, G.A.; McEver, R.P.; Pober, J.S.; Wick, T.; Konkle, B.A.; Schwartz, B.S.; et al. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood 1998, 91, 3527–3561. [Google Scholar] [PubMed]
- Uguccioni, M.; D’Apuzzo, M.; Loetscher, M.; Dewald, B.; Baggiolini, M. Actions of the chemotactic cytokines MCP-1, MCP-2, MCP-3, RANTES, MIP-1α and MIP-1β on human monocytes. Eur. J. Immunol. 1995, 25, 64–68. [Google Scholar] [CrossRef]
- Baggiolini, M.; Dewald, B.; Moser, B. Human Chemokines: An Update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef]
- Haimovitz-Friedman, A.; Cordon-Cardo, C.; Bayoumy, S.; Garzotto, M.; McLoughlin, M.; Gallily, R.; Edwards, C.K.; Schuchman, E.H.; Fuks, Z.; Kolesnick, R. Lipopolysaccharide Induces Disseminated Endothelial Apoptosis Requiring Ceramide Generation. J. Exp. Med. 1997, 186, 1831–1841. [Google Scholar] [CrossRef] [Green Version]
- Penn, M.S.; Chisolm, G.M. Relation between lipopolysaccharide-induced endothelial cell injury and entry of macromolecules into the rat aorta in vivo. Circ. Res. 1991, 68, 1259–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnaswamy, G.; Kelley, J.L.; Yerra, L.; Smith, J.K.; Chi, D.S. Human Endothelium as a Source of Multifunctional Cytokines: Molecular Regulation and Possible Role in Human Disease. J. Interf. Cytokine Res. 1999, 19, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Pu, J.; He, B. Tanshinol protects human umbilical vein endothelial cells against hydrogen peroxide-induced apoptosis. Mol. Med. Rep. 2014, 10, 2764–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, V.; Sharma, S.; Kumar, A.; Singh, S.; Ravichandiran, V. Serratiopeptidase Attenuates Lipopolysaccharide-Induced Vascular Inflammation by Inhibiting the Expression of Monocyte Chemoattractant Protein-1. Curr. Issues Mol. Biol. 2023, 45, 2201-2212. https://doi.org/10.3390/cimb45030142
Yadav V, Sharma S, Kumar A, Singh S, Ravichandiran V. Serratiopeptidase Attenuates Lipopolysaccharide-Induced Vascular Inflammation by Inhibiting the Expression of Monocyte Chemoattractant Protein-1. Current Issues in Molecular Biology. 2023; 45(3):2201-2212. https://doi.org/10.3390/cimb45030142
Chicago/Turabian StyleYadav, Vikas, Satyam Sharma, Ashutosh Kumar, Sanjiv Singh, and V. Ravichandiran. 2023. "Serratiopeptidase Attenuates Lipopolysaccharide-Induced Vascular Inflammation by Inhibiting the Expression of Monocyte Chemoattractant Protein-1" Current Issues in Molecular Biology 45, no. 3: 2201-2212. https://doi.org/10.3390/cimb45030142
APA StyleYadav, V., Sharma, S., Kumar, A., Singh, S., & Ravichandiran, V. (2023). Serratiopeptidase Attenuates Lipopolysaccharide-Induced Vascular Inflammation by Inhibiting the Expression of Monocyte Chemoattractant Protein-1. Current Issues in Molecular Biology, 45(3), 2201-2212. https://doi.org/10.3390/cimb45030142