
Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Salmonella Isolation
2.3. Antibiotic Susceptibility Test
2.4. Genomic DNA Extraction, Whole-Genome Sequencing, and De Novo Assembly
2.5. Annotation
3. Results
3.1. Prevalence of Salmonella spp.
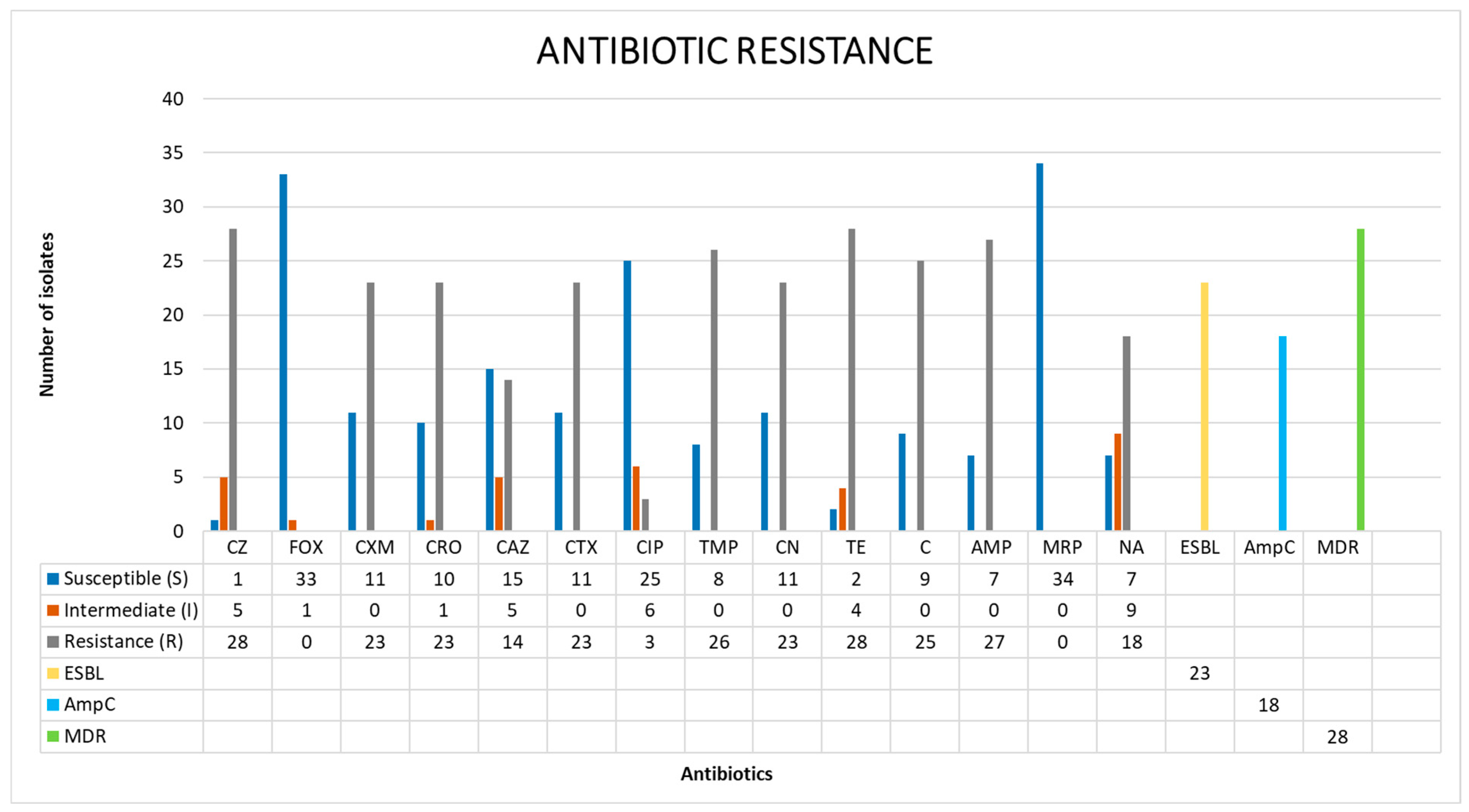
3.2. Antibiotic Resistance Profiles of the Salmonella Isolates
3.2.1. Whole-Genome Sequencing and Genome Characteristics
3.2.2. Antibiotic Resistance Gene Profile
Aminoglycoside
Beta-Lactam
Quinolone
Other Genes
3.3. Plasmid Replicons and Virulence Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Isolation of Samples | CXM (mm) | CRO (mm) | FOX (mm) | CZ (mm) | CTX (mm) | CAZ (mm) | TM (mm) | TE (mm) | C (mm) | CN (mm) | NA (mm) | CIP (mm) | AMP (mm) | MRP (mm) | Resistance Number |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ba Đinh District | R (8) | R (6) | S (25) | R (6) | R (6) | R (6) | R (6) | R (8) | S (26) | R (6) | I (18) | S (27) | R (6) | S (35) | 9 |
| S (22) | I (21) | S (24) | I (20) | S (33) | S (25) | S (33) | I (12) | R (8) | S (20) | S (24) | S (39) | S (21) | S (37) | 1 | |
| R (9) | R (6) | S (24) | R (6) | R (6) | R (6) | R (6) | R (11) | R (6) | R (6) | S (20) | S (31) | R (6) | S (35) | 10 | |
| R (6) | R (9) | S (27) | R (6) | R (6) | R (6) | R (6) | R (6) | R (6) | S (12) | I (17) | S (25) | R (6) | S (30) | 10 | |
| S (27) | S (32) | S (27) | S (23) | S (38) | S (27) | S (32) | S (18) | S (30) | S (20) | I (18) | I (28) | S (21) | S (25) | 0 | |
| Cau Giay District | R (10) | R (10) | S (21) | R (6) | R (8) | R (13) | R (6) | R (6) | R (6) | R (12) | R (6) | S (30) | R (6) | S (34) | 11 |
| R (10) | R (6) | S (18) | R (6) | R (6) | R (6) | R (6) | R (6) | R (6) | R (6) | R (12) | S (27) | R (6) | S (37) | 11 | |
| R (6) | R (6) | S (26) | R (6) | R (6) | R (6) | R (6) | R (10) | R (6) | R (6) | S (20) | S (35) | R (6) | S (35) | 10 | |
| Dong Da District | S (22) | S (34) | S (23) | I (21) | S (31) | S (23) | R (6) | R (11) | R (6) | R (6) | S (21) | S (29) | S (19) | S (32) | 4 |
| R (6) | R (6) | S (25) | R (6) | R (6) | R (6) | R (6) | R (6) | S (25) | R (8) | I (17) | I (23) | R (6) | S (30) | 9 | |
| S (26) | S (37) | S (28) | I (21) | S (34) | S (27) | S (33) | R (11) | S (30) | S (21) | S (27) | S (41) | S (21) | S (26) | 1 | |
| R (6) | R (8) | S (19) | R (6) | R (10) | I (20) | R (6) | R (6) | R (6) | R (11) | R (6) | S (30) | R (6) | S (32) | 10 | |
| R (6) | R (11) | S (25) | R (6) | R (16) | S (22) | R (6) | R (6) | R (6) | R (12) | R (6) | S (32) | R (6) | S (35) | 10 | |
| R (6) | R (12) | S (23) | R (6) | R (19) | S (27) | R (6) | R (6) | R (6) | R (11) | R (6) | S (31) | R (6) | S (35) | 10 | |
| R (6) | R (8) | S (29) | R (6) | R (7) | I (19) | R (6) | R (6) | R (6) | R (10) | R (6) | S (34) | R (6) | S (37) | 10 | |
| R (6) | R (12) | S (25) | R (6) | R (19) | S (21) | S (30) | R (6) | R (6) | R (12) | R (6) | S (32) | R (6) | S (35) | 9 | |
| R (6) | R (9) | I (17) | R (6) | R (9) | R (16) | S (28) | R (6) | R (6) | R (10) | R (6) | S (27) | R (6) | S (31) | 10 | |
| Hoang Mai District | S (22) | S (32) | S (24) | R (17) | S (34) | S (25) | R (6) | R (8) | S (27) | S (20) | I (15) | S (34) | R (6) | S (35) | 4 |
| R (6) | R (6) | S (24) | R (6) | R (6) | R (15) | R (6) | R (11) | R (6) | R (9) | R (6) | R (13) | R (6) | S (35) | 10 | |
| R (6) | R (8) | S (21) | R (6) | R (8) | R (12) | R (6) | R (6) | R (6) | R | R (6) | R (8) | R (6) | S (36) | 10 | |
| R (6) | R (6) | S (33) | R (6) | R (8) | S (26) | R (6) | I (12) | R (6) | R (6) | R (6) | R (18) | R (6) | S (30) | 8 | |
| R (6) | R (8) | S (21) | R (6) | R (9) | S (21) | R (6) | R (6) | R (6) | R (12) | R (6) | S (32) | R (6) | S (36) | 9 | |
| S (21) | S (35) | S (19) | R (15) | S (36) | S (25) | R (6) | R (8) | R (6) | S (17) | R (11) | I (23) | R (6) | S (30) | 5 | |
| S (25) | S (38) | S (27) | I (21) | S (39) | S (27) | S (36) | R (10) | S (32) | S (22) | I (16) | S (34) | S (22) | S (26) | 1 | |
| S (22) | S (33) | S (25) | I (20) | S (35) | S (27) | R (6) | I (14) | S (29) | S (20) | I (15) | S (28) | S (22) | S (23) | 1 | |
| Thanh Xuan District | S (21) | S (31) | S (23) | R (17) | S (32) | I (20) | R (6) | R (8) | R (6) | S (17) | I (15) | S (30) | R (6) | S (30) | 5 |
| R (9) | R (9) | S (23) | R (6) | R (9) | R (13) | R (6) | R (11) | S (24) | R (12) | I (15) | I (24) | R (6) | S (29) | 9 | |
| R (6) | R (6) | S (20) | R (6) | R (6) | R (6) | R (6) | R (6) | R (6) | R (6) | R (12) | I (25) | R (6) | S (34) | 11 | |
| R (10) | R (6) | S (21) | R (6) | R (6) | R (6) | R (6) | I (14) | R (6) | R (6) | R (6) | S (26) | R (6) | S (37) | 10 | |
| S (22) | S (31) | S (24) | R (17) | S (33) | S (23) | R (6) | R (8) | R (6) | S (17) | R (11) | I (23) | R (6) | S (30) | 6 | |
| R (6) | R (11) | S (30) | R (6) | R (9) | R (6) | R (6) | R (10) | R (6) | R (6) | S (20) | S (30) | R (6) | S (35) | 10 | |
| R (6) | R (13) | S (23) | R (6) | R (12) | I (20) | R (6) | R (6) | R (6) | R (12) | R (6) | S (29) | R (6) | S (32) | 10 | |
| R (6) | R (9) | S (20) | R (6) | R (13) | I (19) | S (28) | R (11) | R (6) | S (20) | R (6) | S (31) | R (6) | S (33) | 8 | |
| S (20) | S (31) | S (20) | R (18) | S (30) | S (23) | S (30) | S (19) | S (27) | S (19) | S (22) | S (36) | S (18) | S (35) | 1 | |
| E. coli ATCC 25922 (negative control) | S (28) | S (27) | S (26) | S (34) | S (35) | S (29) | S (30) | S (30) | S (32) | S (30) | S (29) | S (31) | S (32) | S (30) | - |
| Drug Classes | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic Resistance | Code Strain | Strains | Aminoglycoside | Beta-Lactam | Chloramphenicol | Quinolone | Macrolides | Tetracycline | Sulfonamides | Fosfomycin | Diaminopyrimidine | Rifampin | Lincosamide | Polypeptide | Multidrug Classes |
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-AMP | 68_S20 | Kentucky | aac(3)-Iia; aac(3)-Id; aac(3)-IId_1; aac(6)-Iaa_1; aadA17; aadA7_1; ant(3)-Ia_1; aph(3)-Ia_3; rmtB_1; | blaCTX-M-55; blaTEM-206; blaTEM-1B; blaTEM-141; blaTEM-214 | floR_2 | qnrS1_1 | mph(A)-2 | Tet(A)_6; TetR | sul1_5; | fosA3_1 | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; Mrx; | |
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-C-AMP | 43_S11 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(3)-Ia_3; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2 | floR_2 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | |||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-C-AMP | 45_S12 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2 | floR_2 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | |||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-C-AMP | 51_S13 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2; blaTEM-214; blaTEM-206; blaTEM-33; blaTEM-216; blaTEM-209; blaTEM-34; blaTEM-210; blaTEM-141 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | ||||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-AMP | 31_S7 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | ||||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-C-AMP | 42_S10 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2 | floR_2 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | |||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-C-AMP | 57_S16 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(3)-Ia_3; aph(6)-Id_1; | blaCTX-M-55; blaTEM-1B_1 | floR_2 | qnrS1_1 | Tet(A)_6; TetR | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | |||
| CMX-CRO-CZ-CTX-CAZ-TM-CN-TE-C-AMP | 60_S17 | Muenster | aac(3)-Iia; aac(3)-IId_1; aac(6)-Iaa_1; aac(6)-Iy; aadA22; ant(3)-Ia_1; aph(3)-Ia_3; | blaCTX-M-55; blaTEM-1B_1; blaLAP-2 | floR_2 | qnrS1_1 | sul3_2; | dfrA14_5 | ARR-3_4; ARR-2; | lnu(F)_1; linG; | golS; mdsA; mdsB; mdsC; mdtK; sdiA; | ||||
References
- FAO; WHO. M I C R O B I O Salmonella and Campylobacter in Chicken Meat; Center for Emerging: Geneva, Switzerland, 2009. [Google Scholar]
- EFSA; ECDC. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef] [Green Version]
- Center for Emerging; CDCN; Infectious Diseases, Z. National Enteric Disease Surveillance: Salmonella Annual Report 2016; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2016.
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States 2019; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. [CrossRef] [Green Version]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Juniora, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-Analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA; ECDC. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Appleby, M.C. Whom Should We Eat? Why Veal Can Be Better for Welfare than Chicken; Appleby, M.C., Weary, D.M., Sandoe, P., Eds.; CABI: Wallingford, UK, 2014. [Google Scholar] [CrossRef]
- Tran, T.P.; Ly, T.L.K.; Nguyen, T.T.; Akiba, M.; Ogasawara, N.; Shinoda, D.; Okatani, A.T.; Hayashidani, H. Prevalence of Salmonella Spp. in Pigs, Chickens and Ducks in the Mekong Delta, Vietnam. J. Vet. Med. Sci. 2004, 66, 1011–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.Y.; Kang, M.; Yoon, R.H.; Park, C.K.; Moon, O.K.; Jang, H.K. Prevalence and Antimicrobial Susceptibility of Salmonella Isolates in Pekin Ducks from South Korea. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 473–479. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Venkitanarayanan, K.; Johny, A.K. Antibiotic-Resistant Salmonella in the Food Supply and the Potential Role of Antibiotic Alternatives for Control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; de Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348. [Google Scholar] [CrossRef] [Green Version]
- Castro-Vargas, R.E.; Herrera-Sánchez, M.P.; Rodríguez-Hernández, R.; Rondón-Barragán, I.S. Antibiotic Resistance in Salmonella Spp. Isolated from Poultry: A Global Overview. Vet. World 2020, 13, 2070–2084. [Google Scholar] [CrossRef]
- Hagag, A.; Naguib, D.; Mohamed, A.A.; El-Gohary, A.H. Prevalence, Virulence Genes, and Antibiotic Resistance of Escherichia Coli and Salmonella Spp. Isolated from Pigeons and Humans ARTICLE HISTORY ABSTRACT. Mansoura. Vet. Med. J. 2022, 23, 24–30. [Google Scholar] [CrossRef]
- Carrique-Mas, J.J.; Trung, N.V.; Hoa, N.T.; Mai, H.H.; Thanh, T.H.; Campbell, J.I.; Wagenaar, J.A.; Hardon, A.; Hieu, T.Q.; Schultsz, C. Antimicrobial Usage in Chicken Production in the Mekong Delta of Vietnam. Zoonoses Public Health 2014, 62 (Suppl. 1), 70–78. [Google Scholar] [CrossRef] [Green Version]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6, 261–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.; Chiu, C.-H. Forum on Antimicrobial Resistance Evolution of the Virulence Plasmids of Non-Typhoid Salmonella and Its Association with Antimicrobial Resistance. Microbes Infect. 2006, 8, 1931–1936. [Google Scholar] [CrossRef] [PubMed]
- Thi Phan, T.; Thi Lien Khai, L.; Ogasawara, N.; Thu Tam, N.; Tomomitsu Okatani, A.; Akiba, M.; Hayashidani, H. Contamination of Salmonella in Retail Meats and Shrimps in the Mekong Delta, Vietnam. J. Food Prot. 2005, 68, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, T. Salmonella in Domestic Animals; Wray, C., Wray, A., Eds.; CABI International: Oxford, UK, 2000. [Google Scholar]
- Löfström, C.; Hansen, T.; Maurischat, S.; Malorny, B. Salmonella: Salmonellosis. Encycl. Food Health 2015, 701–705. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Nguyen, L.T.; Chau, T.T.H.; Nguyen, T.T.; Tran, B.N.; Taniguchi, T.; Hayashidani, H.; Ly, K.T.L. Prevalence and Antibiotic Resistance of Salmonella Isolated from Poultry and Its Environment in the Mekong Delta, Vietnam. Vet. World 2021, 14, 3216. [Google Scholar] [CrossRef]
- Tsai, H.J.; Hsiang, P.H. The Prevalence and Antimicrobial Susceptibilities of Salmonella and Campylobacter in Ducks in Taiwan. J. Vet. Med. Sci. 2005, 67, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Scaltriti, E.; Sassera, D.; Comandatore, F.; Morganti, C.M.; Mandalari, C.; Gaiarsa, S.; Bandi, C.; Zehender, G.; Bolzoni, L.; Casadei, G.; et al. Differential Single Nucleotide Polymorphism-Based Analysis of an Outbreak Caused by Salmonella Enterica Serovar Manhattan Reveals Epidemiological Details Missed by Standard Pulsed-Field Gel Electrophoresis. J. Clin. Microbiol. 2015, 53, 1227. [Google Scholar] [CrossRef] [Green Version]
- Achtman, M.; Wain, J.; Weill, F.X.; Nair, S.; Zhou, Z.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Multilocus Sequence Typing as a Replacement for Serotyping in Salmonella Enterica. PLoS Pathog. 2012, 8, e1009040. [Google Scholar] [CrossRef] [Green Version]
- Gilchrist, C.A.; Turner, S.D.; Riley, M.F.; Petri, W.A.; Hewlett, E.L. Whole-Genome Sequencing in Outbreak Analysis. Clin. Microbiol. Rev. 2015, 28, 541. [Google Scholar] [CrossRef] [Green Version]
- Köser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-Genome Sequencing to Control Antimicrobial Resistance. Trends Genet. 2014, 30, 401. [Google Scholar] [CrossRef] [Green Version]
- La, T.M.; Kim, T.; Lee, H.J.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Lee, S.W. Whole-Genome Analysis of Multidrug-Resistant Salmonella Enteritidis Strains Isolated from Poultry Sources in Korea. Pathogens 2021, 10, 1615. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; den Bakker, H.C.; Hendriksen, R.S. Genomic Epidemiology: Whole-Genome-Sequencing-Powered Surveillance and Outbreak Investigation of Foodborne Bacterial Pathogens. Annu. Rev. Food Sci. Technol. 2016, 7, 16–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pornsukarom, S.; van Vliet, A.H.M.; Thakur, S. Whole Genome Sequencing Analysis of Multiple Salmonella Serovars Provides Insights into Phylogenetic Relatedness, Antimicrobial Resistance, and Virulence Markers across Humans, Food Animals and Agriculture Environmental Sources. BMC Genom. 2018, 19, 801. [Google Scholar] [CrossRef] [Green Version]
- USDA. Isolation and Identification of Salmonella from Meat, Poultry, Pasteurized Egg, and Siluriformes (Fish) Products and Carcass and Environmental Sponges. Revision 10. 2019. Available online: https://content.govdelivery.com/accounts/USFSIS/bulletins/2ed04a9 (accessed on 24 August 2022).
- Hasman, H.; Agersø, Y.; Hendriksen, R.; Cavaco, L.M.; Guerra-Roman, B. Isolation of ESBL-, AmpC-and Carbapenemase-Producing E. coli from Caecal Samples. LAB. PROTOCOL. 2019, 1–18. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility. Clin. Lab. Stand. Inst. 2022, 32, 1–16. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Assefa, S.; Keane, T.M.; Otto, T.D.; Newbold, C.; Berriman, M. ABACAS: Algorithm-Based Automatic Contiguation of Assembled Sequences. Bioinformatics 2009, 25, 1968–1969. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, C.E.; Kruczkiewicz, P.; Laing, C.R.; Lingohr, E.J.; Gannon, V.P.J.; Nash, J.H.E.; Taboada, E.N. The Salmonella in Silico Typing Resource (SISTR): An Open Web-Accessible Tool for Rapidly Typing and Subtyping Draft Salmonella Genome Assemblies. PLoS ONE 2016, 11, e0147101. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antiobiotic Resistance Genes. Available online: https://github.com/tseemann/abricate (accessed on 24 August 2022).
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of Mobile Genetic Elements Associated with Antibiotic Resistance in Salmonella Enterica Using a Newly Developed Web Tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Flament, A.; Soubbotina, A.; Mainil, J.; Marlier, D. Prevalence of Salmonella Serotypes in Male Mule Ducks in Belgium. Vet. Rec. 2012, 170, 311. [Google Scholar] [CrossRef]
- Adzitey, F.; Huda, N.; Rahmat Ali, G.R. Prevalence and Antibiotic Resistance of Campylobacter, Salmonella, and L. Monocytogenes in Ducks: A Review. Foodborne Pathog. Dis. 2012, 9, 498–505. [Google Scholar] [CrossRef]
- Chen, Z.; Bai, J.; Wang, S.; Zhang, X.; Zhan, Z.; Shen, H.; Zhang, H.; Wen, J.; Gao, Y.; Liao, M.; et al. Prevalence, Antimicrobial Resistance, Virulence Genes and Genetic Diversity of Salmonella Isolated from Retail Duck Meat in Southern China. Microorganisms 2020, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Lai, J.; Wang, Y.; Liu, S.; Li, Y.; Liu, K.; Shen, J.; Wu, C. Prevalence and Characterization of Salmonella Species Isolated from Pigs, Ducks and Chickens in Sichuan Province, China. Int. J. Food Microbiol. 2013, 163, 14–18. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly Prevalent Multidrug-Resistant Salmonella From Chicken and Pork Meat at Retail Markets in Guangdong, China. Front. Microbiol. 2018, 9, 2104. [Google Scholar] [CrossRef] [Green Version]
- Trongjit, S.; Angkititrakul, S.; Tuttle, R.E.; Poungseree, J.; Padungtod, P.; Chuanchuen, R. Prevalence and Antimicrobial Resistance in Salmonella Enterica Isolated from Broiler Chickens, Pigs and Meat Products in Thailand–Cambodia Border Provinces. Microbiol. Immunol. 2017, 61, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, T.H.A.; Hai, C.V.; Ha, H.Y.; Tu, N.H.K. Antibiotic Resistance in Salmonella Isolated from Ho Chi Minh City (Vietnam) and Difference of Sulfonamide Resistance Gene Existence in Serovars. J. Pure. Appl. Microbiol. 2021, 15, 2244–2251. [Google Scholar] [CrossRef]
- Van, T.T.H.; Moutafis, G.; Istivan, T.; Tran, L.T.; Coloe, P.J. Detection of Salmonella Spp. in Retail Raw Food Samples from Vietnam and Characterization of Their Antibiotic Resistance. Appl. Environ. Microbiol. 2007, 73, 6885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Peng, J.; Guan, X.; Li, J.; Huang, X.; Liu, S.; Wen, Y.; Zhao, Q.; Huang, X.; Yan, Q.; et al. Genetic and Antimicrobial Resistance Profiles of Salmonella Spp. Isolated from Ducks along the Slaughter Line in Southwestern China. Food Control 2020, 107, 106805. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.Y.; Wang, Y.; Sun, F.; Li, W.; Wu, H.; Shen, P.C.; Pan, Z.M.; Jiao, X. Emergence of 16S RRNA Methylase Gene RmtB in Salmonella Enterica Serovar London and Evolution of RmtB-Producing Plasmid Mediated by IS26. Front. Microbiol. 2021, 11, 604278. [Google Scholar] [CrossRef]
- Zhou, D.; Li, X.-Z.; Canada, H.; Pina Fratamico, C.; Chen, W.; Fang, T.; Zhou, X.; Zhang, D.; Shi, X.; Shi, C. IncHI2 Plasmids Are Predominant in Antibiotic-Resistant Salmonella Isolates. Front. Microbiol. 2016, 7, 1566. [Google Scholar] [CrossRef] [Green Version]
- Atrice Blanc-Potard, A.-B.; Solomon, F.; Kayser, J.; Groisman, E.A. The SPI-3 Pathogenicity Island of Salmonella Enterica. J. Bacteriol. 1999, 181, 998–1004. [Google Scholar] [CrossRef] [Green Version]

| Sample | Serovar | Serogroup | H1 | H2 | O Antigen | MLST |
|---|---|---|---|---|---|---|
| 31_S7 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 42_S10 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 43_S11 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 45_S12 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 51_S13 | Muenster | E1 | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 57_S16 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 60_S17 | Muenster | - | e,h | 1,5 | 3,{10}{15}{15,34} | 321 |
| 68_S20 | Kentucky | C2-C3 | i | z6 | 8,20 | 198 |
| Sample | Reads | Average Read Length (bp) | Contigs | Genome Length (bp) | Average Contig Length (bp) | N50 (bp) | GC (%) | SRA Accession Number |
|---|---|---|---|---|---|---|---|---|
| 31_S7 | 808,752 | 238 | 305 | 4,701,119 | 129,321 | 34,942 | 52.29 | SRR21051813 |
| 42_S10 | 683,258 | 237 | 425 | 4,671,790 | 103,647 | 28,050 | 52.45 | SRR21051784 |
| 43_S11 | 677,248 | 238 | 493 | 4,905,076 | 101,196 | 23,437 | 52.15 | SRR21051783 |
| 45_S12 | 749,140 | 239 | 334 | 4,744,687 | 160,345 | 37,074 | 52.27 | SRR21051781 |
| 51_S13 | 783,124 | 239 | 329 | 4,766,796 | 157,729 | 36,997 | 52.32 | SRR21051780 |
| 57_S16 | 591,734 | 237 | 382 | 4,655,893 | 156,597 | 30,153 | 52.41 | SRR21051777 |
| 60_S17 | 655,310 | 238 | 493 | 4,682,797 | 87,054 | 22,011 | 52.51 | SRR21051776 |
| 68_S20 | 910,644 | 237 | 464 | 4,804,148 | 181,020 | 24,436 | 52.47 | SRR21051782 |
| Samples | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 68_S20 | 43_S11 | 45_S12 | 51_S13 | 31_S7 | 42_S10 | 57_S16 | 60_S17 | ||||
| Serovar | Kentucky | Muenster | Muenster | Muenster | Muenster | Muenster | Muenster | Muenster | |||
| Genes/Number of genes identified | 33 | 27 | 26 | 33 | 25 | 26 | 26 | 24 | |||
| Drug classes | Rifampin | arr-3_4 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| arr2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Aminoglycoside | aac(3)-Iia | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| aac(3)-IId_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| aac(3)-Id_1 | 1 | Absence (Negative) | |||||||||
| aac(6)-Iaa_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| aac(6)-Iy | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| aadA17 | 1 | ||||||||||
| aadA22 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| aadA7_1 | 1 | ||||||||||
| ant(3)-Ia_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| aph(3)-Ia_3 | 1 | 1 | 1 | 1 | |||||||
| aph(6)-Id_1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||
| rmtB_1 | 1 | ||||||||||
| Beta-lactam | blaCTX-M-55_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| blaLAP-2 | 1 | 1 | 1 | 1 | 1 | 1 | |||||
| blaTEM-141 | 1 | 1 | |||||||||
| blaTEM-1B_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| blaTEM-206 | 1 | 1 | |||||||||
| blaTEM-209 | 1 | ||||||||||
| blaTEM-210 | 1 | ||||||||||
| blaTEM-214 | 1 | 1 | |||||||||
| blaTEM-216 | 1 | ||||||||||
| blaTEM-33 | 1 | presence (Positive) | |||||||||
| blaTEM-34 | 1 | ||||||||||
| Fosfomycin | fosA3_1 | 1 | |||||||||
| Chloramphenicol | floR_2 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| Diaminopyrimidine | dfrA14_5 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Lincosamide | linG | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Inu(F)_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Quinolone | qnrS1_1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Macrolides | mph(A)-2 | 1 | |||||||||
| Sulfonamides | sul1_5 | 1 | |||||||||
| sul3_2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| Tetracyclin | tet(A)_6 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| tetR | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| Multi-drug classes | golS | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| mdsA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| mdsB | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| mdsC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| mdtK | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Mrx | 1 | ||||||||||
| sdiA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Strains | Serotype | Plasmid | Number of Virulence Factors | Number of Virulence Genes | SPI |
|---|---|---|---|---|---|
| 31_S7 | Muenster | 24 | 82 | SPI-1, SPI-2, SPI-3, SPI-5, SPI-9, SPI-13, SPI-14 | |
| 42_S10 | Muenster | 23 | 79 | C63PI, SPI-1, SPI-2, SPI-3, SPI-5, SPI-9, SPI-13, SPI-14 | |
| 43_S11 | Muenster | IncHI2_1 IncHI2A_1 | 20 | 75 | Not_named, SPI-1, SPI-2, SPI-3, SPI-9, SPI-13, SPI-14 |
| 45_S12 | Muenster | 21 | 79 | SPI-1, SPI-2, SPI-3, SPI-5, SPI-13, SPI-14 | |
| 51_S13 | Muenster | 23 | 84 | C63PI, SPI-1, SPI-2, SPI-3, SPI-5, SPI-9, SPI-13 | |
| 57_S16 | Muenster | 23 | 81 | C63PI, SPI-1, SPI-2, SPI-3, SPI-13 | |
| 60_S17 | Muenster | IncL/M(pMU407)_1_pMU407 | 21 | 72 | Not_named, SPI-1, SPI-2, SPI-3, SPI-13, SPI-14 |
| 68_S20 | Kentucky | 23 | 83 | SPI-1, SPI-2, SPI-3, SPI-9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.T.; Le, H.V.; Vu Thi Hai, H.; Nguyen Tuan, T.; Nguyen, H.M.; Pham Xuan, D.; Tran Thi Thanh, H.; Le Thi, H.H. Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam. Curr. Issues Mol. Biol. 2023, 45, 2213-2229. https://doi.org/10.3390/cimb45030143
Nguyen TT, Le HV, Vu Thi Hai H, Nguyen Tuan T, Nguyen HM, Pham Xuan D, Tran Thi Thanh H, Le Thi HH. Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam. Current Issues in Molecular Biology. 2023; 45(3):2213-2229. https://doi.org/10.3390/cimb45030143
Chicago/Turabian StyleNguyen, Trung Thanh, Hoa Vinh Le, Ha Vu Thi Hai, Thanh Nguyen Tuan, Huong Minh Nguyen, Da Pham Xuan, Huyen Tran Thi Thanh, and Hao Hong Le Thi. 2023. "Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam" Current Issues in Molecular Biology 45, no. 3: 2213-2229. https://doi.org/10.3390/cimb45030143
APA StyleNguyen, T. T., Le, H. V., Vu Thi Hai, H., Nguyen Tuan, T., Nguyen, H. M., Pham Xuan, D., Tran Thi Thanh, H., & Le Thi, H. H. (2023). Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam. Current Issues in Molecular Biology, 45(3), 2213-2229. https://doi.org/10.3390/cimb45030143