

Change in Long Non-Coding RNA Expression Profile Related to the Antagonistic Effect of Clostridium perfringens Type C on Piglet Spleen

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Total RNA Isolation
2.3. Library Preparation for lncRNA Sequencing
2.4. Quality Control and Mapping
2.5. Transcriptome Assembly and Expression Level Quantification
2.6. Coding Potential Analysis and Target Gene Prediction
2.7. Enrichment Analysis of GO and KEGG
2.8. Heat Map Construction and lncRNA Secondary Structure Prediction
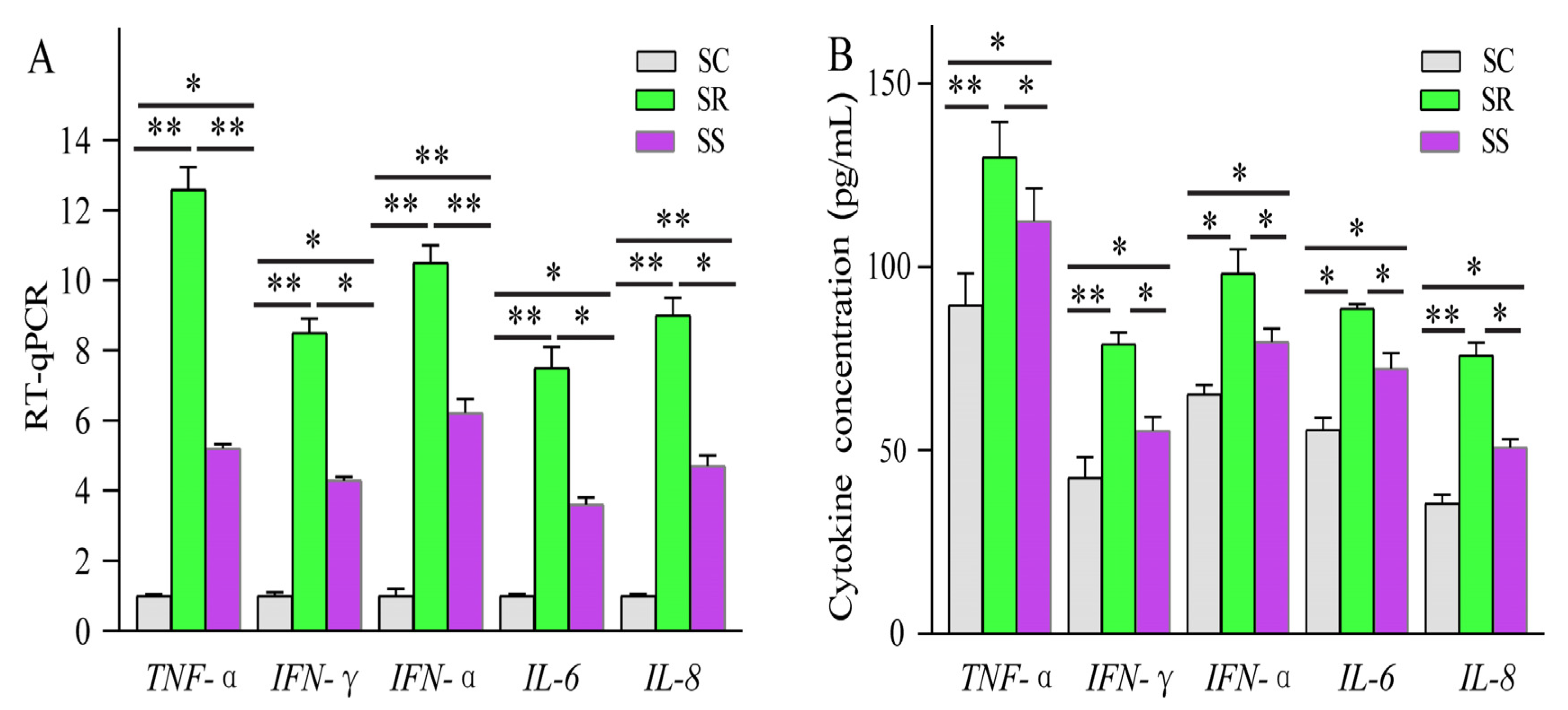
2.9. RT-qPCR Assay and ELISA Detection
3. Results
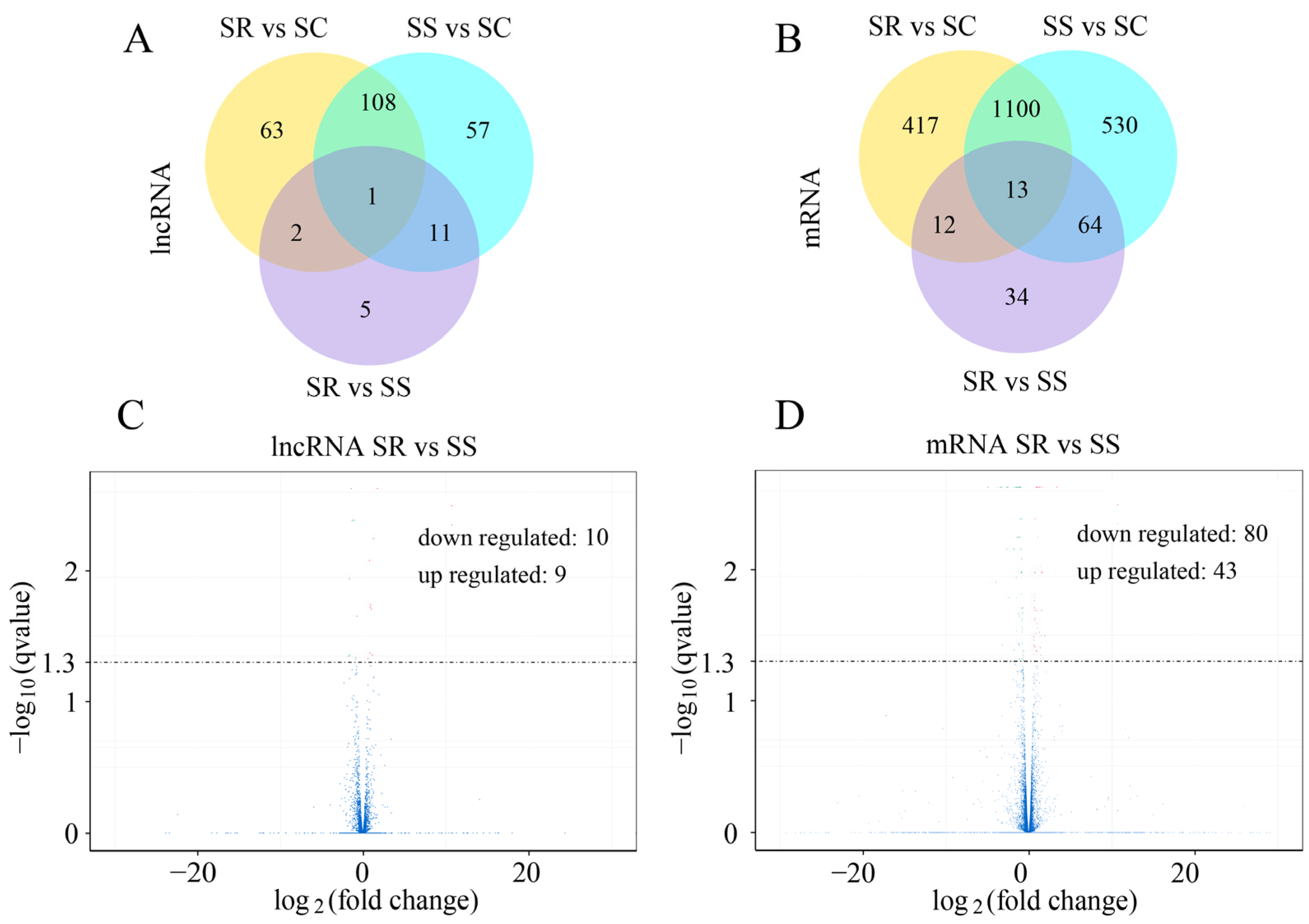
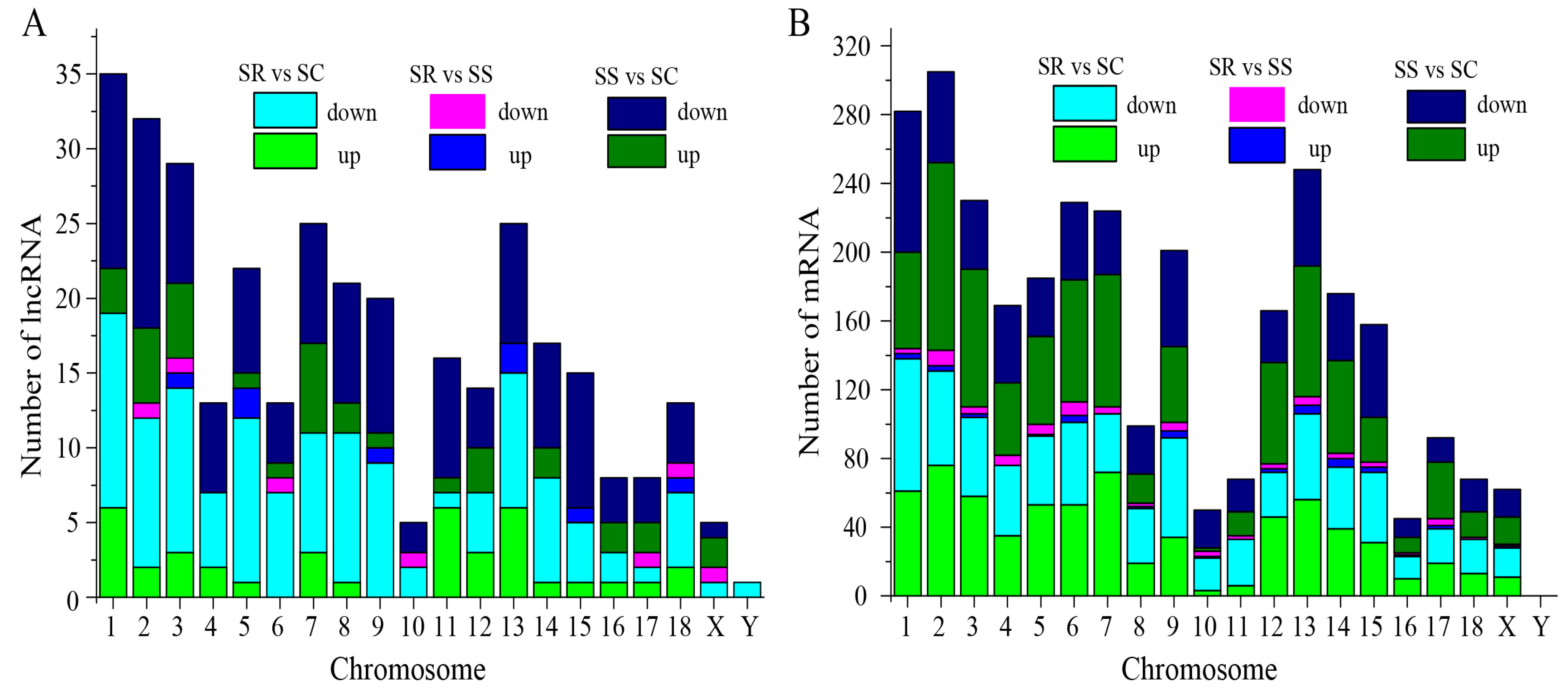
3.1. Analyses of Differentially Expressed lncRNAs
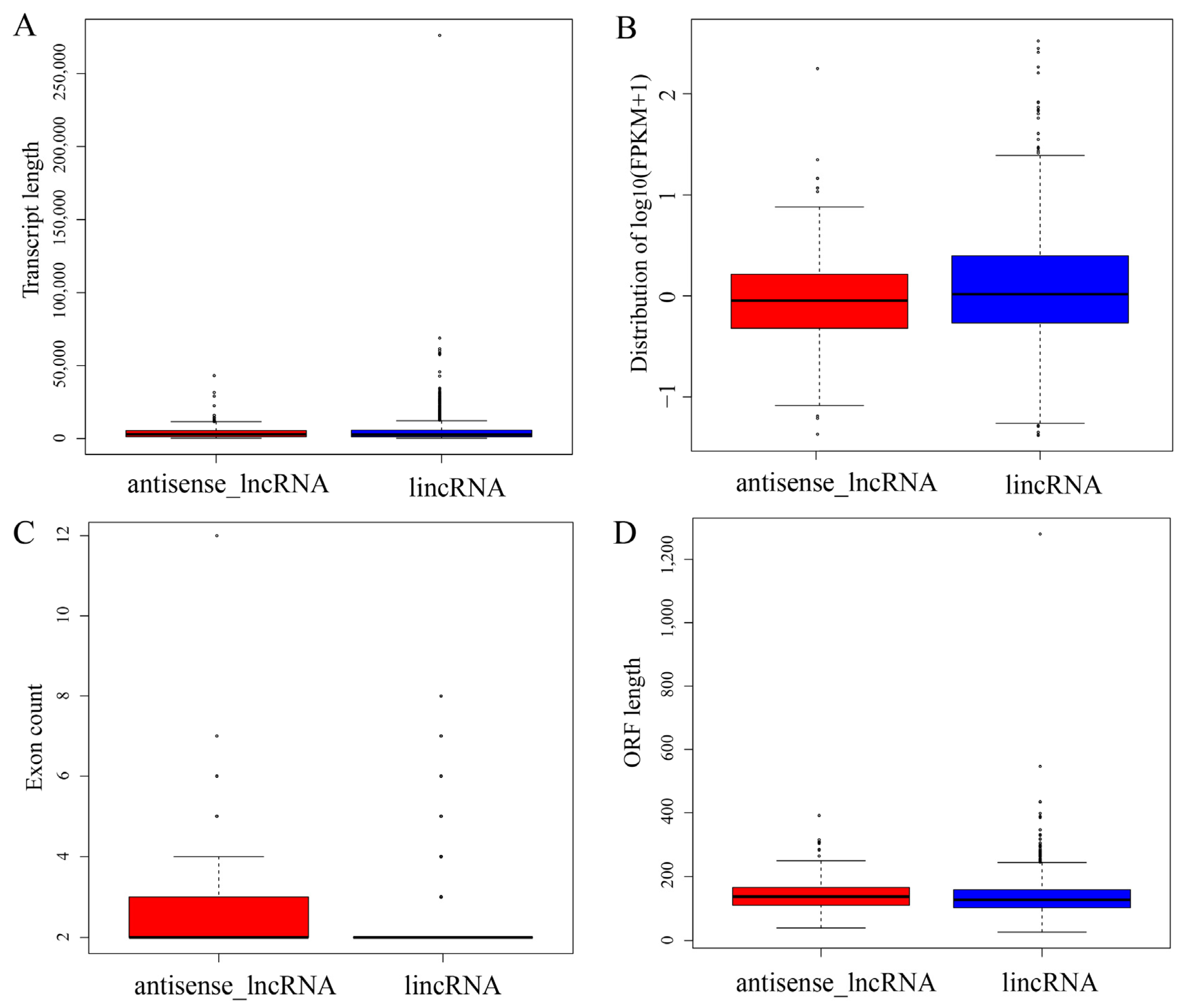
3.2. Characterization of lncRNA Subtypes
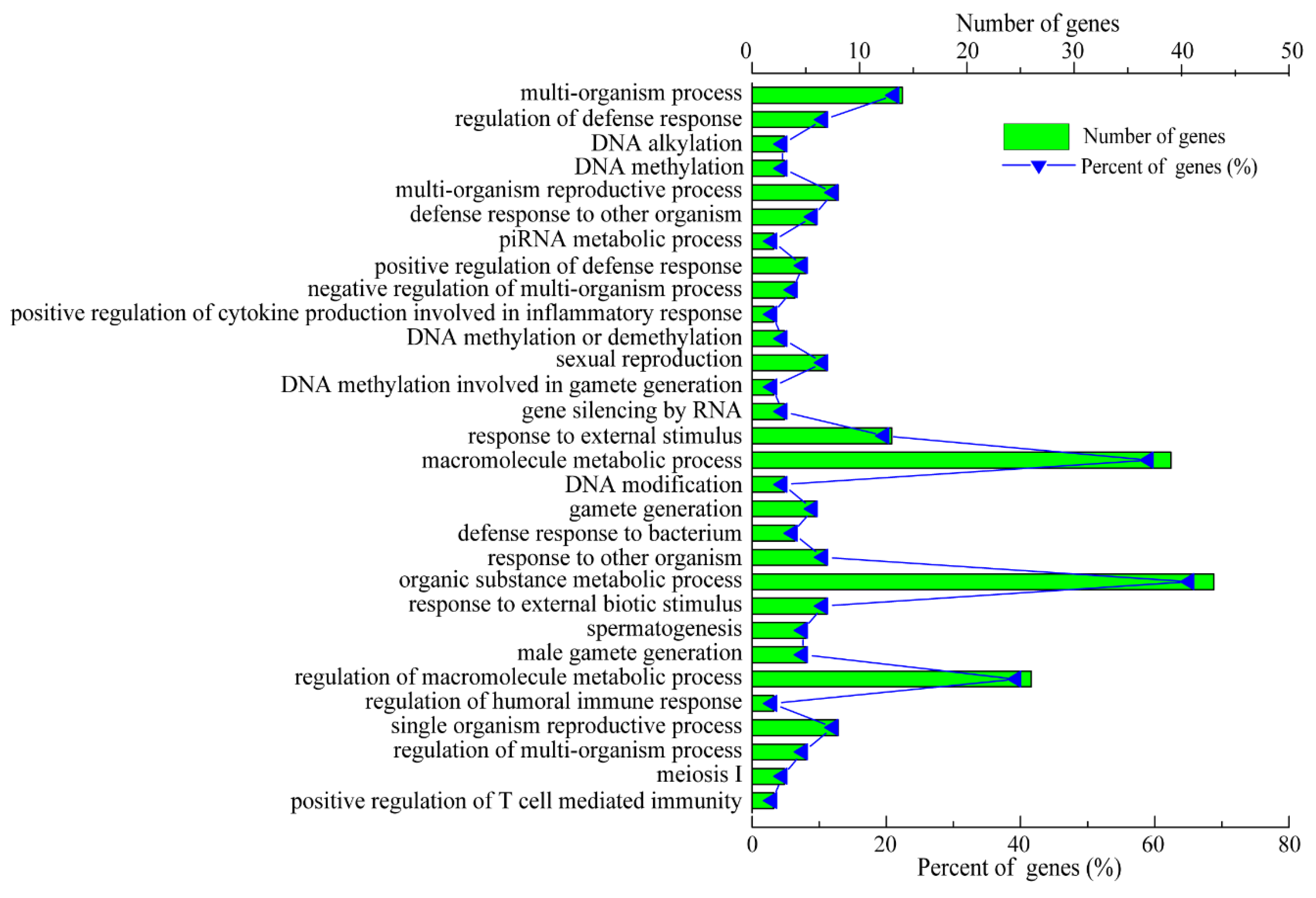
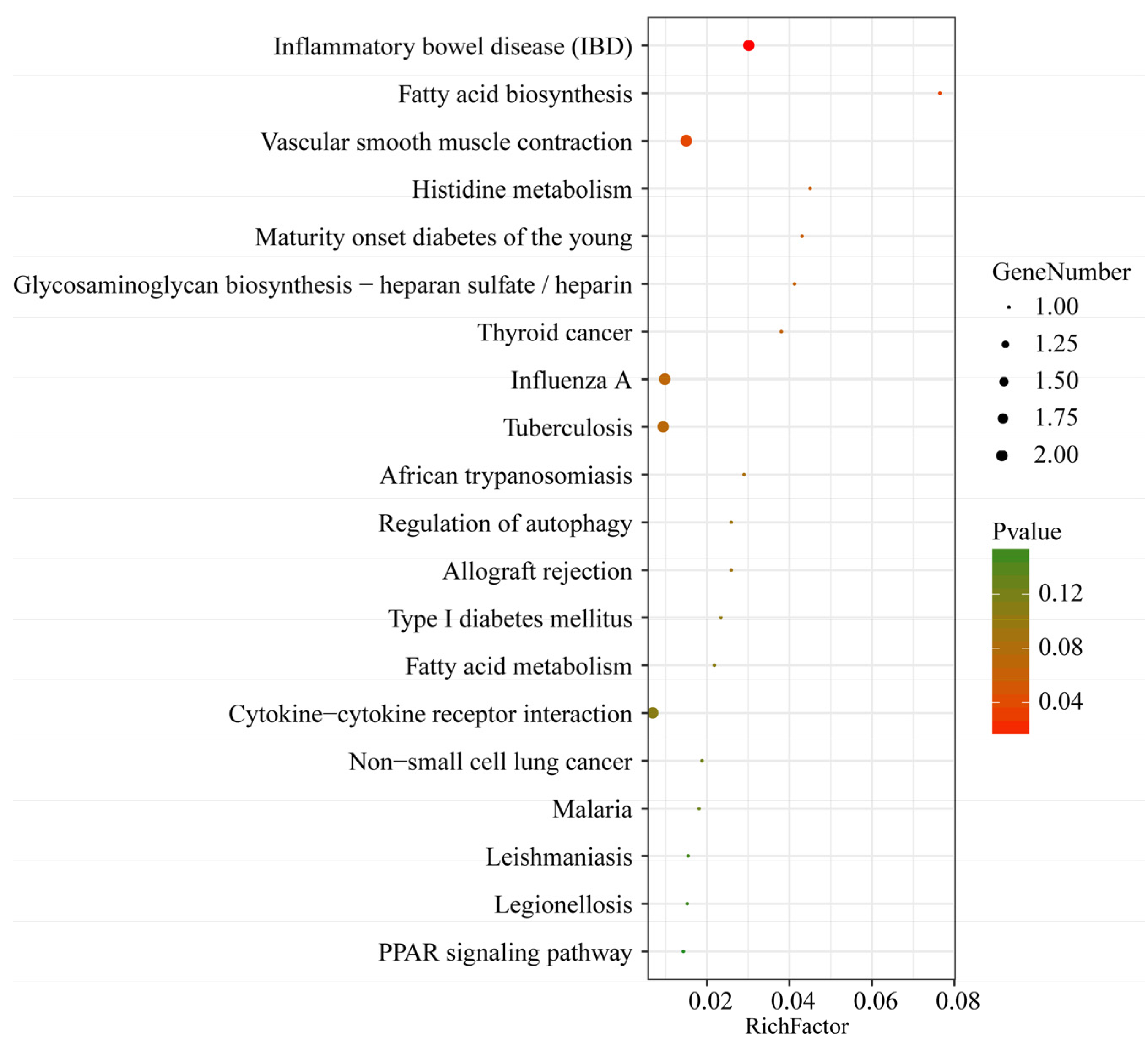
3.3. Functional Analyses of C. perfringens Type C-Responsive lncRNAs and mRNAs
3.4. RT-qPCR Validation and Tissue Expression Profiling of LNC_001595
3.5. The Identification of lncRNAs Antagonistic to C. perfringens Type C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Songer, J.G. Clostridia as agents of zoonotic disease. Vet. Microbiol. 2010, 140, 399–404. [Google Scholar] [CrossRef]
- Songer, J.G.; Uzal, F.A. Clostridial enteric infections in pigs. J. Vet. Diagn. Investig. 2005, 17, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Miclard, J.; Jaggi, M.; Sutter, E.; Wyder, M.; Grabscheid, B.; Posthaus, H. Clostridium perfringens beta-toxin targets endothelial cells in necrotizing enteritis in piglets. Vet. Microbiol. 2009, 137, 320–325. [Google Scholar] [CrossRef]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Uzal, F.A.; McClane, B.A. Recent progress in understanding the pathogenesis of Clostridium perfringens type C infections. Vet. Microbiol. 2011, 153, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, O.; Okabe, A. Clostridial hydrolytic enzymes degrading extracellular components. Toxicon 2001, 39, 1769–1780. [Google Scholar] [CrossRef]
- Sakurai, J.; Nagahama, M.; Oda, M. Clostridium perfringens alpha-toxin: Characterization and mode of action. J. Biochem. 2004, 136, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, V.L.; Martel, A.; Pasmans, F.; Van Immerseel, F.; Posthaus, H. Endothelial binding of beta toxin to small intestinal mucosal endothelial cells in early stages of experimentally induced Clostridium perfringens type C enteritis in pigs. Vet. Pathol. 2013, 50, 626–629. [Google Scholar] [CrossRef]
- Lawrence, G.; Cooke, R. Experimental pigbel: The production and pathology of necrotizing enteritis due to Clostridium welchii type C in the guinea-pig. Br. J. Exp. Pathol. 1980, 61, 261–271. [Google Scholar]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Health Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.; Ma, X.; Liu, Y.; Qiao, S.; Hou, Y.; Zhang, G. Dietary modulation of endogenous host defense peptide synthesis as an alternative approach to in-feed antibiotics. Anim. Nutr. 2018, 4, 160–169. [Google Scholar] [CrossRef]
- Worley, M.J. Immune evasion and persistence in enteric bacterial pathogens. Gut Microbes 2023, 15, 2163839. [Google Scholar] [CrossRef]
- Plotniece, A.; Sobolev, A.; Supuran, C.T.; Carta, F.; Bjoerkling, F.; Franzyk, H.; Yli-Kauhaluoma, J.; Augustyns, K.; Cos, P.; De Vooght, L.; et al. Selected strategies to fight pathogenic bacteria. J. Enzym. Inhib. Med. Chem. 2023, 38, 2155816. [Google Scholar] [CrossRef]
- Vogeli, P.; Meijerink, E.; Fries, R.; Neuenschwander, S.; Vorlander, N.; Stranzinger, G.; Bertschinger, H.U. A molecular test for the detection of E. Coli F18 receptors: A breakthrough in the struggle against edema disease and post-weaning diarrhea in swine. Schweiz. Arch. Tierheilkd. 1997, 139, 479–484. [Google Scholar]
- Anh Duc, T.; Hong, Y.H.; Lillehoj, H.S. RNA-Seq profiles of immune related genes in the spleen of necrotic enteritis-afflicted chicken lines. Asian-Australas. J. Anim. Sci. 2015, 28, 1496–1511. [Google Scholar]
- Jie, Z.; Chun-Yan, L.; Jing-Ping, L.; Ren, G.; Hui, W.; Juan, P.; Sheng-Lan, L. Immunoregulation on mice of low immunity and effects on five kinds of human cancer cells of panax japonicus polysaccharide. Evid.-Based Complement. Altern. Med. 2015, 2015, 839697. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yu, Y.; Zhang, X.; Huang, B.; Xu, C.; Zhang, B.; Bai, P.; Liu, C. Phenotypic, genomic, and transcriptomic changes in an acinetobacter baumannii strain after spaceflight in China’s tiangong-2 space laboratory. Braz. J. Microbiol. 2022, 53, 1447–1464. [Google Scholar] [CrossRef]
- Wu, W.-H.; Hui, T.-C.; Wu, Q.-Q.; Xu, C.-A.; Zhou, Z.-W.; Wang, S.-H.; Zheng, W.; Yin, Q.-Q.; Li, X.; Pan, H.-Y. Pneumocystis jirovecii and legionella pneumophila coinfection in a patient with diffuse large B-cell lymphoma: A case report. World J. Clin. Cases 2021, 9, 8595–8601. [Google Scholar] [CrossRef]
- Kosinska-Selbi, B.; Mielczarek, M.; Szyda, J. Review: Long non-coding RNA in livestock. Animal 2020, 14, 2003–2013. [Google Scholar] [CrossRef]
- Goudarzi, M.; Berg, K.; Pieper, L.M.; Schier, A.F. Individual long non-coding RNAs have no overt functions in zebrafish embryogenesis, viability and fertility. Elife 2019, 8, e40815. [Google Scholar] [CrossRef]
- Iempridee, T.; Wiwithaphon, S.; Piboonprai, K.; Pratedrat, P.; Khumkhrong, P.; Japrung, D.; Temisak, S.; Laiwejpithaya, S.; Chaopotong, P.; Dharakul, T. Identification of reference genes for circulating long noncoding RNA analysis in serum of cervical cancer patients. FEBS Open Bio 2018, 8, 1844–1854. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Sirbe, C.; Simu, G.; Szabo, I.; Grama, A.; Pop, T.L. Pathogenesis of autoimmune hepatitis-cellular and molecular mechanisms. Int. J. Mol. Sci. 2021, 22, 13578. [Google Scholar] [CrossRef]
- Zhao, C.-N.; Mao, Y.-M.; Liu, L.-N.; Li, X.-M.; Wang, D.-G.; Pan, H.-F. Emerging role of lncRNAs in systemic lupus erythematosus. Biomed. Pharmacother. 2018, 106, 584–592. [Google Scholar] [CrossRef]
- Marete, A.; Ariel, O.; Ibeagha-Awemu, E.; Bissonnette, N. Identification of long non-coding RNA isolated from naturally infected macrophages and associated with bovine johne’s disease in Canadian holstein using a combination of neural networks and logistic regression. Front. Vet. Sci. 2021, 8, 639053. [Google Scholar] [CrossRef]
- Lodde, V.; Floris, M.; Muroni, M.R.; Cucca, F.; Idda, M.L. Non-coding RNAs in malaria infection. Wiley Interdiscip. Rev.-RNA 2022, 13, e1697. [Google Scholar] [CrossRef]
- Guegan, F.; Rajan, K.S.; Bento, F.; Pinto-Neves, D.; Sequeira, M.; Guminska, N.; Mroczek, S.; Dziembowski, A.; Cohen-Chalamish, S.; Doniger, T.; et al. A long noncoding RNA promotes parasite differentiation in African trypanosomes. Sci. Adv. 2022, 8, eabn2706. [Google Scholar] [CrossRef]
- Bronte, V.; Pittet, M.J. The spleen in local and systemic regulation of immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Gawandi, S.; Vandna; Yadav, I.; Mohan, H.; Desai, V.G.; Kumar, S. Analysis of fluoro based pyrazole analogues as a potential therapeutics candidate against japanese encephalitis virus infection. Virus Res. 2023, 323, 198955. [Google Scholar] [CrossRef]
- Hosseindoust, A.; Ha, S.; Lokhande, A.; Mun, J.; Kim, Y.I.; Kim, J. The targeted anti-salmonella bacteriophage attenuated the inflammatory response of laying hens challenged with salmonella gallinarum. Poult. Sci. 2023, 102, 102296. [Google Scholar] [CrossRef]
- Jin, C.; Bao, J.; Wang, Y.; Chen, W.; Wu, T.; Wang, L.; Lv, X.; Gao, W.; Wang, B.; Zhu, G.; et al. Changes in long non-coding RNA expression profiles related to the antagonistic effects of escherichia coli F17 on lamb spleens. Sci. Rep. 2018, 8, 16514. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Bao, J.; Wang, Y.; Chen, W.; Zou, S.; Wu, T.; Wang, L.; Lv, X.; Gao, W.; Wang, B.; et al. Changes in circRNA expression profiles related to the antagonistic effects of escherichia coli F17 in lamb spleens. Sci. Rep. 2018, 8, 14524. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Hue, D.; Lillehoj, H.S.; Song, K.-D.; Oh, J.-D. Differential regulation of microRNA transcriptome in chicken lines resistant and susceptible to necrotic enteritis disease. Poult. Sci. 2014, 93, 1383–1395. [Google Scholar] [CrossRef]
- Gaur, U.; Xiong, Y.Y.; Luo, Q.P.; Yuan, F.Y.; Wu, H.Y.; Qiao, M.; Wimmers, K.; Li, K.; Mei, S.Q.; Liu, G.S. Breed-specific transcriptome response of spleen from six to eight week old piglet after infection with streptococcus suis type 2. Mol. Biol. Rep. 2014, 41, 7865–7873. [Google Scholar] [CrossRef]
- Yan, Z.; Huang, X.; Sun, W.; Yang, Q.; Shi, H.; Jiang, T.; Li, S.; Wang, P.; Gun, S. Analyses of long non-coding RNA and mRNA profiling in the spleen of diarrheic piglets caused by Clostridium perfringens type C. PeerJ 2018, 6, e5997. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Sun, W.Y.; Yan, Z.Q.; Shi, H.R.; Yang, Q.L.; Wang, P.F.; Li, S.G.; Liu, L.X.; Zhao, S.G.; Gun, S.B. Novel insights reveal anti-microbial gene regulation of piglet intestine immune in response to Clostridium perfringens infection. Sci. Rep. 2019, 9, 1963. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-Seq experiments with Tophat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Guttman, M.; Garber, M.; Levin, J.Z.; Donaghey, J.; Robinson, J.; Adiconis, X.; Fan, L.; Koziol, M.J.; Gnirke, A.; Nusbaum, C.; et al. Ab initio reconstruction of cell type-specific transcriptomes in mouse reveals the conserved multi-exonic structure of lincRNAs. Nat. Biotechnol. 2010, 28, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-Seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG orthology (ko) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Broadbent, K.M.; Broadbent, J.C.; Ribacke, U.; Wirth, D.; Rinn, J.L.; Sabeti, P.C. Strand-specific RNA sequencing in plasmodium falciparum malaria identifies developmentally regulated long non-coding RNA and circular RNA. BMC Genom. 2015, 16, 454. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Guo, Y.; Zhao, C.; Yuan, S.; Wang, Y.; Tang, G.; Zhou, W.; Sun, S. Hepatitis b virus X protein (HBx)-related long noncoding RNA (lncRNA) down-regulated expression by HBx (Dreh) inhibits hepatocellular carcinoma metastasis by targeting the intermediate filament protein vimentin. Hepatology 2013, 57, 1882–1892. [Google Scholar] [CrossRef]
- Li, M.; Cui, J.; Niu, W.; Huang, J.; Feng, T.; Sun, B.; Yao, H. Long non-coding PCED1B-AS1 regulates macrophage apoptosis and autophagy by sponging miR-155 in active tuberculosis. Biochem. Biophys. Res. Commun. 2019, 509, 803–809. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, S.; Liu, X.; Liu, H.; Hu, T.; Qiu, X.; Zhang, J.; Lei, M. Analyses of long non-coding RNA and mRNA profiling using RNA sequencing during the pre-implantation phases in pig endometrium. Sci. Rep. 2016, 6, 20238. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Wang, G.; Chen, L.; Jiang, J.; Liu, L.; Li, N.; Zhao, J.; Sun, X.; Zhou, P. Genome-wide analysis of long non-coding RNAs at early stage of skin pigmentation in goats (capra hircus). BMC Genom. 2016, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.-M.; Codogno, P. Autophagic cell death loch ness monster or endangered species? Autophagy 2011, 7, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Long, Y.C.; Shen, H.-M. Differential regulatory functions of three classes of phosphatidylinositol and phosphoinositide 3-kinases in autophagy. Autophagy 2015, 11, 1711–1728. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, L.; Greshock, J.; Colligon, T.A.; Wang, Y.; Ward, R.; Katsaros, D.; Lassus, H.; Butzow, R.; Godwin, A.K.; et al. Frequent genetic abnormalities of the PI3K/AKT pathway in primary ovarian cancer predict patient outcome. Genes Chromosomes Cancer 2011, 50, 606–618. [Google Scholar] [CrossRef] [Green Version]
- El-Diwany, R.; Soliman, M.; Sugawara, S.; Breitwieser, F.; Skaist, A.; Coggiano, C.; Sangal, N.; Chattergoon, M.; Bailey, J.R.; Siliciano, R.F.; et al. CMPK2 and BCL-G are associated with type 1 interferon-induced hiv restriction in humans. Sci. Adv. 2018, 4, eaat0843. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-L.; Lin, D.-W.; Chang, Z.-F. Identification of a putative human mitochondrial thymidine monophosphate kinase associated with monocytic/macrophage terminal differentiation. Genes Cells 2008, 13, 679–689. [Google Scholar] [CrossRef]
- Ebrahimi, K.H. A unifying view of the broad-spectrum antiviral activity of RSAD2 (viperin) based on its radical-sam chemistry. Metallomics 2018, 10, 539–552. [Google Scholar] [CrossRef]
- Chin, K.C.; Cresswell, P. Viperin (cig5), an ifn-inducible antiviral protein directly induced by human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2001, 98, 15125–15130. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhang, X.; Wang, S.; Wang, J.; Du, B.; Wang, Z.; Liu, M.; Jiang, W.; Qian, M.; Ren, H. GPR97 is dispensable for metabolic syndrome but is involved in macrophage inflammation in high-fat diet-induced obesity in mice. Sci. Rep. 2016, 6, 24649. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.; Chen, X.; Lu, S.; Kuang, Y.; Fei, J.; Wang, Z. GPR97/ADGRG3 ameliorates experimental autoimmune encephalomyelitis by regulating cytokine expression. Acta Biochim. Biophys. Sin. 2018, 50, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Hoffmeyer, A.; Piekorz, R.; Moriggl, R.; Ihle, J.N. GADD45G is dispensable for normal mouse development and T-cell proliferation. Mol. Cell. Biol. 2001, 21, 3137–3143. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Yu, H.; Chow, C.; Li, B.; Zheng, W.; Davis, R.J.; Flavell, R.A. GADD45G mediates the activation of the p38 and jnk map kinase pathways and cytokine production in effector TH1 cells. Immunity 2001, 14, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Lu, B. The molecular mechanisms that control function and death of effector CD4+ T cells. Immunol. Res. 2006, 36, 275–282. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
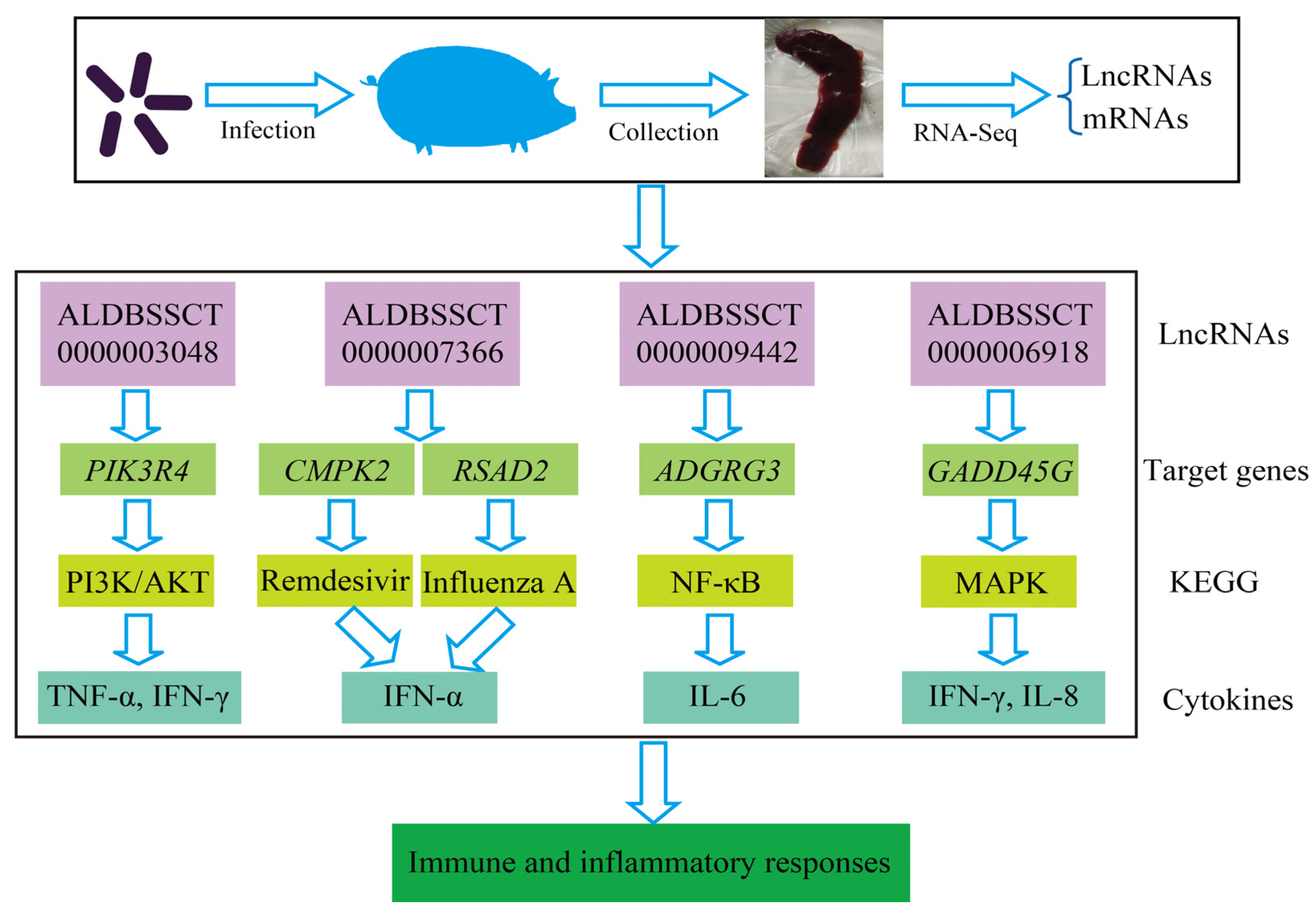{kind=link}
| Transcript_Id | Length | Gene_Id | SR | SS | p | Q |
|---|---|---|---|---|---|---|
| LNC_001595 | 6577 | XLOC_093069 | 1.36 | 0.67 | 0.0009 | 0.0196 |
| ALDBSSCT0000003048 | 519 | ALDBSSCG0000001826 | 18.27 | 5.33 | 0.0001 | 0.0023 |
| LNC_000191 | 2965 | XLOC_011580 | 2.11 | 3.81 | 0.0030 | 0.0469 |
| ALDBSSCT0000009442 | 233 | ALDBSSCG0000005758 | 56.02 | 171.52 | 0.0004 | 0.0114 |
| LNC_001065 | 2890 | XLOC_064212 | 0.33 | 1.06 | 0.0028 | 0.0449 |
| LNC_001987 | 20,029 | XLOC_118880 | 2.89 | 4.73 | 0.0010 | 0.0220 |
| LNC_000042 | 4774 | XLOC_002451 | 1.00 | 0.45 | 0.0027 | 0.0438 |
| ALDBSSCT0000006918 | 1302 | ALDBSSCG0000004208 | 14.20 | 33.32 | 0.0001 | 0.0041 |
| LNC_001097 | 8758 | XLOC_065996 | 2.21 | 1.13 | 0.0008 | 0.0181 |
| LNC_000263 | 280 | XLOC_016001 | 12.34 | 23.03 | 0.0001 | 0.0023 |
| LNC_002009 | 1490 | XLOC_120225 | 3.13 | 7.56 | 0.0001 | 0.0041 |
| ALDBSSCT0000007366 | 504 | ALDBSSCG0000004471 | 27.97 | 13.09 | 0.0002 | 0.0056 |
| LNC_001253 | 13,487 | XLOC_074796 | 0.59 | 1.53 | 0.0001 | 0.0023 |
| LNC_001985 | 31,206 | XLOC_118644 | 2.25 | 1.45 | 0.0030 | 0.0469 |
| LncRNA Transcript_Id | Trans Target Genes | Cis Target Genes |
|---|---|---|
| ALDBSSCT0000003048 | PIK3R4 | |
| LNC_000191 | GTPBP8 | |
| ALDBSSCT0000009442 | ADGRG3 | SPOCD1 |
| LNC_001987 | ZNF648 | |
| ALDBSSCT0000006918 | FAM167B, IL17RD, GADD45G, LY6G5B, ZGRF1, IQCJ, LRFN5, DDR1, NCALD, IL17F, POU4F3, IL17RA, SLC2A13, CCDC110, LIN28A, DEFB123, MUSK, CD63, TTYH1, RXRB, RBFOX1, MCAT, GGT1, CA7, SYCN, MRC2, CALCRL, PDZD4, HIST1H2BA, MROH1, SPINK5, IL12A, PGPEP1L, PAG6, PNCK, ICAM2, MARCH3, ADAMTS4, SSMEM1, PLAC8L1, LCN8, HRASLS5, SLC39A14, ONECUT2 | |
| LNC_001097 | TFEC | |
| LNC_000263 | RPL8, SPO11, MAEL, BTG4, WEE2, NLRP9, OTX2, H1FOO, HAL, FKBP6, PAX6, FOXR1, TCL1A, BCL2L10, ZP4, HMGB2, NLRP11 | ZC3H7A |
| ALDBSSCT0000007366 | RSAD2, RNF144A, CMPK2 | |
| LNC_001985 | ZNF398, PDIA4, ZNF425, ZNF786 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Z.; Wang, P.; Yang, Q.; Gao, X.; Gun, S.; Huang, X. Change in Long Non-Coding RNA Expression Profile Related to the Antagonistic Effect of Clostridium perfringens Type C on Piglet Spleen. Curr. Issues Mol. Biol. 2023, 45, 2309-2325. https://doi.org/10.3390/cimb45030149
Yan Z, Wang P, Yang Q, Gao X, Gun S, Huang X. Change in Long Non-Coding RNA Expression Profile Related to the Antagonistic Effect of Clostridium perfringens Type C on Piglet Spleen. Current Issues in Molecular Biology. 2023; 45(3):2309-2325. https://doi.org/10.3390/cimb45030149
Chicago/Turabian StyleYan, Zunqiang, Pengfei Wang, Qiaoli Yang, Xiaoli Gao, Shuangbao Gun, and Xiaoyu Huang. 2023. "Change in Long Non-Coding RNA Expression Profile Related to the Antagonistic Effect of Clostridium perfringens Type C on Piglet Spleen" Current Issues in Molecular Biology 45, no. 3: 2309-2325. https://doi.org/10.3390/cimb45030149
APA StyleYan, Z., Wang, P., Yang, Q., Gao, X., Gun, S., & Huang, X. (2023). Change in Long Non-Coding RNA Expression Profile Related to the Antagonistic Effect of Clostridium perfringens Type C on Piglet Spleen. Current Issues in Molecular Biology, 45(3), 2309-2325. https://doi.org/10.3390/cimb45030149