

Effects of Large Extracellular Vesicles from Kidney Cancer Patients on the Growth and Environment of Renal Cell Carcinoma Xenografts in a Mouse Model
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Patients Included
2.2. Primary Cell Isolation and Culture
2.3. Immunofluorescence
2.4. Doubling Time of Primary Cells
2.5. lEV Isolation
2.6. Animal Model
2.7. Immunofluorescent Labeling of Tumors
2.8. Western Blot Analysis of Mouse Native Kidney
2.9. Statistical Analysis
3. Results
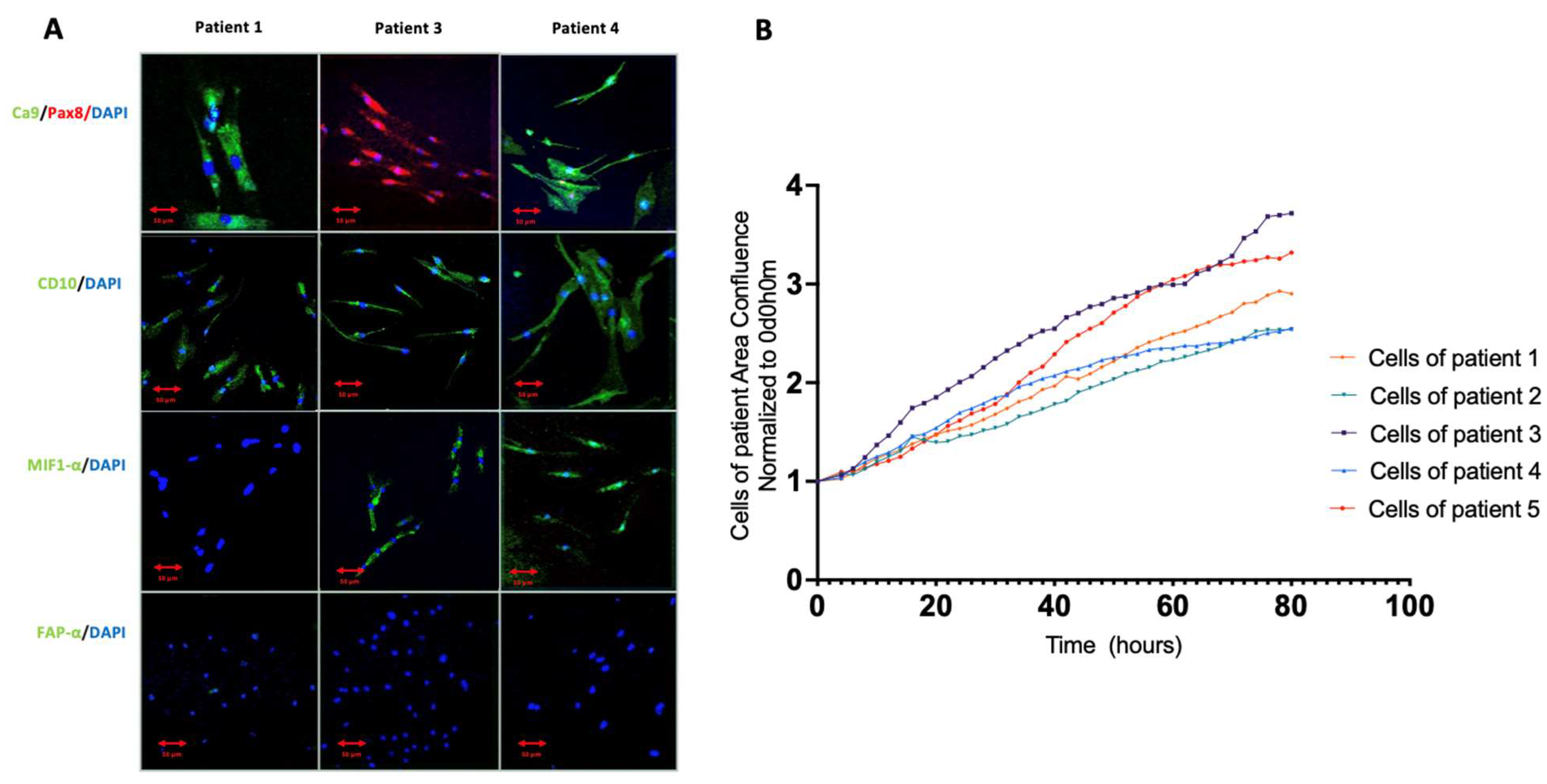
3.1. Primary Cell Line Characterization
3.2. Effect of lEVs on Tumor Growth and the Peritumoral Environment
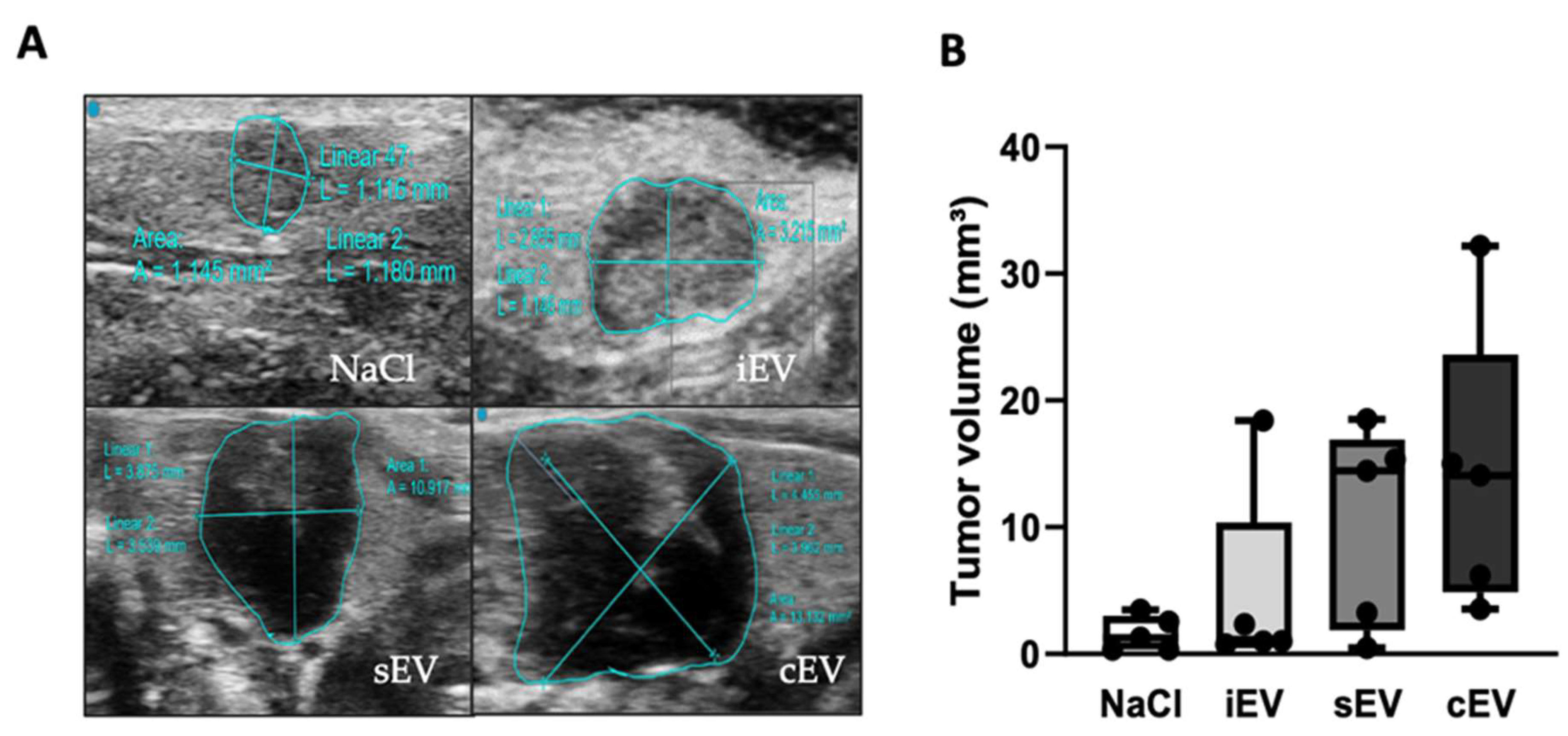
3.2.1. lEVs from Cancer Patients Tend to Increase Xenograft Growth
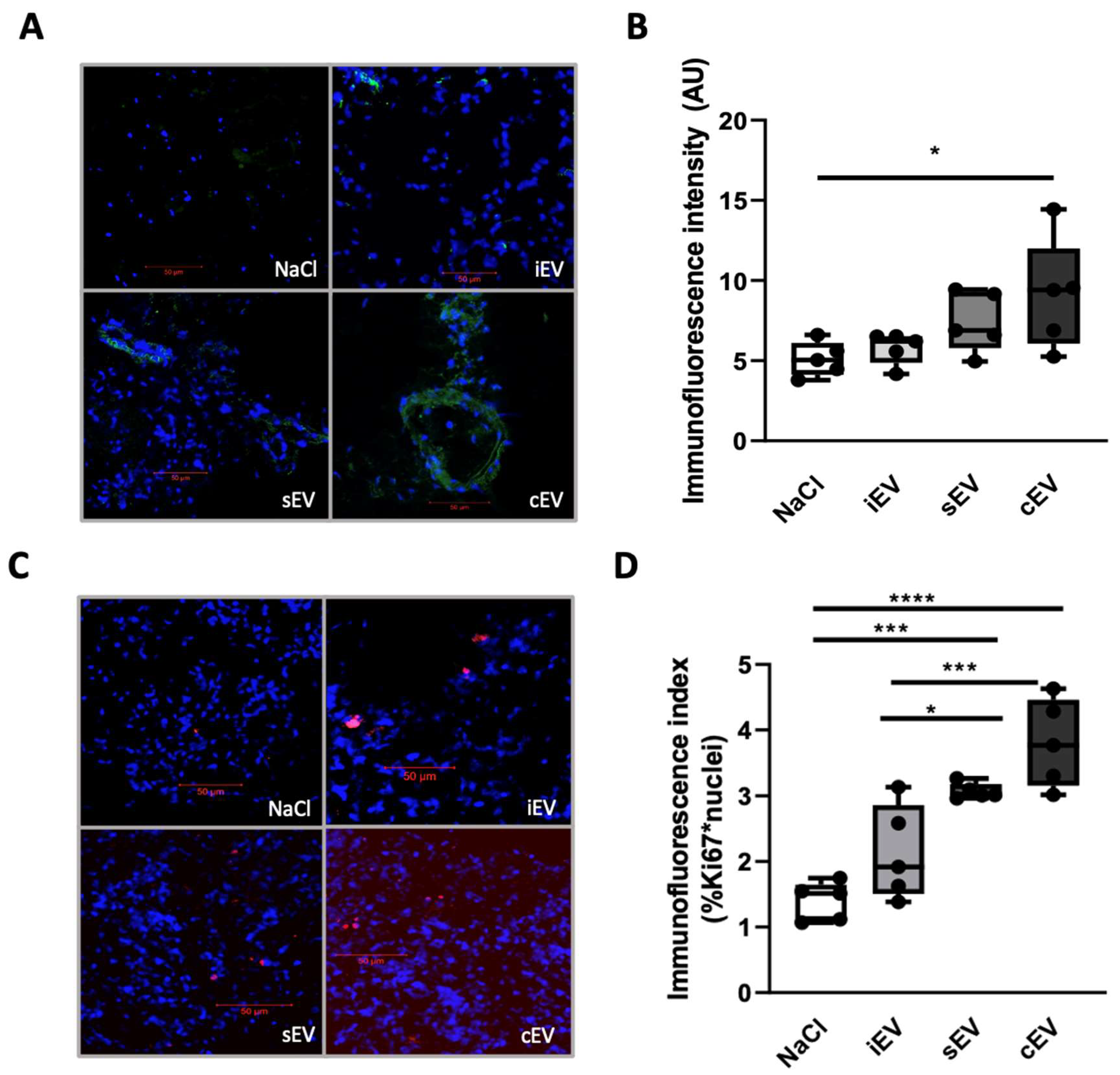
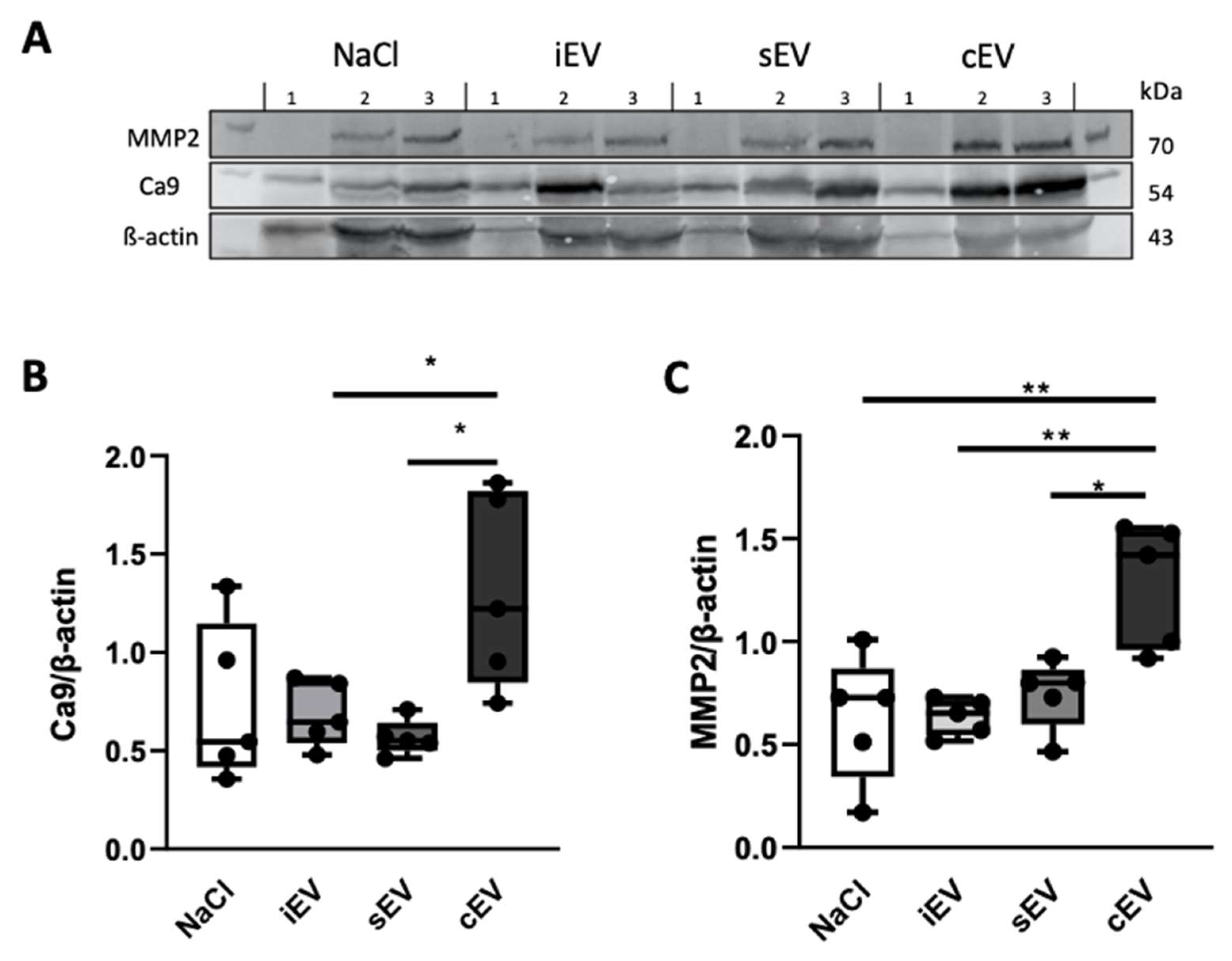
3.2.2. lEVs from Cancer Patients Altered Peritumoral Environment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Capitanio, U.; Montorsi, F. Renal Cancer. Lancet 2016, 387, 894–906. [Google Scholar] [CrossRef]
- Neuzillet, Y.; Tillou, X.; Mathieu, R.; Long, J.-A.; Gigante, M.; Paparel, P.; Poissonnier, L.; Baumert, H.; Escudier, B.; Lang, H.; et al. Renal Cell Carcinoma (RCC) in Patients with End-Stage Renal Disease Exhibits Many Favourable Clinical, Pathologic, and Outcome Features Compared with RCC in the General Population. Eur. Urol. 2011, 60, 366–373. [Google Scholar] [CrossRef]
- Patard, J.-J.; Kim, H.L.; Lam, J.S.; Dorey, F.J.; Pantuck, A.J.; Zisman, A.; Ficarra, V.; Han, K.-R.; Cindolo, L.; De La Taille, A.; et al. Use of the University of California Los Angeles Integrated Staging System to Predict Survival in Renal Cell Carcinoma: An International Multicenter Study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2004, 22, 3316–3322. [Google Scholar] [CrossRef] [PubMed]
- Zigeuner, R.; Hutterer, G.; Chromecki, T.; Imamovic, A.; Kampel-Kettner, K.; Rehak, P.; Langner, C.; Pummer, K. External Validation of the Mayo Clinic Stage, Size, Grade, and Necrosis (SSIGN) Score for Clear-Cell Renal Cell Carcinoma in a Single European Centre Applying Routine Pathology. Eur. Urol. 2010, 57, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Motzer, R.J.; McDermott, D.F.; Escudier, B.; Burotto, M.; Choueiri, T.K.; Hammers, H.J.; Barthélémy, P.; Plimack, E.R.; Porta, C.; George, S.; et al. Conditional Survival and Long-Term Efficacy with Nivolumab plus Ipilimumab versus Sunitinib in Patients with Advanced Renal Cell Carcinoma. Cancer 2022, 128, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.; Zanotti, G.; Kim, R.; Krulewicz, S.; Leith, A.; Bailey, A.; Liu, F.X.; Kearney, M. Real-World Symptoms, Disease Burden, Resource Use and Quality of Life in US Patients with Advanced Renal Cell Cancer. Future Oncol. 2021, 17, 2169–2182. [Google Scholar] [CrossRef]
- Rossi, S.H.; Klatte, T.; Stewart, G.D. Quality of Life Outcomes in Patients with Localised Renal Cancer: A Literature Review. World J. Urol. 2018, 36, 1961–1972. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Andriantsitohaina, R.; Gaceb, A.; Vergori, L.; Martínez, M.C. Microparticles as Regulators of Cardiovascular Inflammation. Trends Cardiovasc. Med. 2012, 22, 88–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malloci, M.; Perdomo, L.; Veerasamy, M.; Andriantsitohaina, R.; Simard, G.; Martínez, M.C. Extracellular Vesicles: Mechanisms in Human Health and Disease. Antioxid. Redox Signal. 2019, 30, 813–856. [Google Scholar] [CrossRef] [PubMed]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.-J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with Immune Modulatory Features Are Present in Human Breast Milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, M.M.; Campelo, F.; Liska, N.; Chernomordik, L.V.; Marrink, S.J.; McMahon, H.T. Mechanisms Shaping Cell Membranes. Curr. Opin. Cell Biol. 2014, 29, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junker, K.; Heinzelmann, J.; Beckham, C.; Ochiya, T.; Jenster, G. Extracellular Vesicles and Their Role in Urologic Malignancies. Eur. Urol. 2016, 70, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Grange, C.; Brossa, A.; Bussolati, B. Extracellular Vesicles and Carried MiRNAs in the Progression of Renal Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 1832. [Google Scholar] [CrossRef] [Green Version]
- Skog, J.; Wurdinger, T.; van Rijn, S.; Meijer, D.; Gainche, L.; Sena-Esteves, M.; Curry, W.T.; Carter, R.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma Microvesicles Transport RNA and Protein That Promote Tumor Growth and Provide Diagnostic Biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Du, T.; Ju, G.; Wu, S.; Cheng, Z.; Cheng, J.; Zou, X.; Zhang, G.; Miao, S.; Liu, G.; Zhu, Y. Microvesicles Derived from Human Wharton’s Jelly Mesenchymal Stem Cells Promote Human Renal Cancer Cell Growth and Aggressiveness through Induction of Hepatocyte Growth Factor. PloS ONE 2014, 9, e96836. [Google Scholar] [CrossRef] [Green Version]
- Lindoso, R.S.; Collino, F.; Camussi, G. Extracellular Vesicles Derived from Renal Cancer Stem Cells Induce a Pro-Tumorigenic Phenotype in Mesenchymal Stromal Cells. Oncotarget 2015, 6, 7959–7969. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, X.; Luo, C.; Chen, X.; Yang, L.; Tao, J.; Shi, J. The 786-0 Renal Cancer Cell-Derived Exosomes Promote Angiogenesis by Downregulating the Expression of Hepatocyte Cell Adhesion Molecule. Mol. Med. Rep. 2013, 8, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Grange, C.; Tapparo, M.; Collino, F.; Vitillo, L.; Damasco, C.; Deregibus, M.C.; Tetta, C.; Bussolati, B.; Camussi, G. Microvesicles Released from Human Renal Cancer Stem Cells Stimulate Angiogenesis and Formation of Lung Premetastatic Niche. Cancer Res. 2011, 71, 5346–5356. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to Build a Bridge from a Population-Based to a More “Personalized” Approach to Cancer Staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Vergori, L.; Martinez, M.C.; Bigot, P. Circulating Large Extracellular Vesicles Carrying CA9 in the Diagnosis and Prognosis of Clear-cell Renal Cell Carcinoma. Clin. Transl. Med. 2021, 11, e358. [Google Scholar] [CrossRef]
- Tomayko, M.M.; Reynolds, C.P. Determination of Subcutaneous Tumor Size in Athymic (Nude) Mice. Cancer Chemother. Pharmacol. 1989, 24, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, N.; Tanaka, A.; Ohe, C.; Nagashima, Y. Recent Advances of Immunohistochemistry for Diagnosis of Renal Tumors. Pathol. Int. 2013, 63, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Bond, K.H.; Chiba, T.; Wynne, K.P.H.; Vary, C.P.H.; Sims-Lucas, S.; Coburn, J.M.; Oxburgh, L. The Extracellular Matrix Environment of Clear Cell Renal Cell Carcinoma Determines Cancer Associated Fibroblast Growth. Cancers 2021, 13, 5873. [Google Scholar] [CrossRef]
- Inamura, K. Renal Cell Tumors: Understanding Their Molecular Pathological Epidemiology and the 2016 WHO Classification. Int. J. Mol. Sci. 2017, 18, 2195. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Wright, B.M.; Li, X.; Finke, J.; Rini, B.I.; Zhou, M.; He, H.; Lal, P.; Welford, S.M. Tumor-Derived Macrophage Migration Inhibitory Factor Promotes an Autocrine Loop That Enhances Renal Cell Carcinoma. Oncogene 2013, 32, 1469–1474. [Google Scholar] [CrossRef] [Green Version]
- Errarte, P.; Guarch, R.; Pulido, R.; Blanco, L.; Nunes-Xavier, C.E.; Beitia, M.; Gil, J.; Angulo, J.C.; López, J.I.; Larrinaga, G. The Expression of Fibroblast Activation Protein in Clear Cell Renal Cell Carcinomas Is Associated with Synchronous Lymph Node Metastases. PLoS ONE 2016, 11, e0169105. [Google Scholar] [CrossRef]
- Yap, N.Y.; Ong, T.A.; Morais, C.; Pailoor, J.; Gobe, G.C.; Rajandram, R. Establishment of Epithelial and Fibroblast Cell Cultures and Cell Lines from Primary Renal Cancer Nephrectomies. Cell Biol. Int. 2019, 43, 715–725. [Google Scholar] [CrossRef]
- Lobo, N.C.; Gedye, C.; Apostoli, A.J.; Brown, K.R.; Paterson, J.; Stickle, N.; Robinette, M.; Fleshner, N.; Hamilton, R.J.; Kulkarni, G.; et al. Efficient Generation of Patient-Matched Malignant and Normal Primary Cell Cultures from Clear Cell Renal Cell Carcinoma Patients: Clinically Relevant Models for Research and Personalized Medicine. BMC Cancer 2016, 16, 485. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Piwocka, O.; Musielak, M.; Piotrowski, I.; Suchorska, W.M.; Trzeciak, T. From Donor to the Lab: A Fascinating Journey of Primary Cell Lines. Front. Cell Dev. Biol. 2021, 9, 711381. [Google Scholar] [CrossRef] [PubMed]
- Kim, F.J.; Campagna, A.; Khandrika, L.; Koul, S.; Byun, S.-S.; vanBokhoven, A.; Moore, E.E.; Koul, H. Individualized Medicine for Renal Cell Carcinoma: Establishment of Primary Cell Line Culture from Surgical Specimens. J. Endourol. 2008, 22, 2361–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, H.; Béraud, C.; Bethry, A.; Danilin, S.; Lindner, V.; Coquard, C.; Rothhut, S.; Massfelder, T. Establishment of a Large Panel of Patient-Derived Preclinical Models of Human Renal Cell Carcinoma. Oncotarget 2016, 7, 59336–59359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Manley, B.J.; Becerra, M.F.; Redzematovic, A.; Casuscelli, J.; Tennenbaum, D.M.; Reznik, E.; Han, S.; Benfante, N.; Chen, Y.-B.; et al. Tumor Xenografts of Human Clear Cell Renal Cell Carcinoma but Not Corresponding Cell Lines Recapitulate Clinical Response to Sunitinib: Feasibility of Using Biopsy Samples. Eur. Urol. Focus 2017, 3, 590–598. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Xu, P.; Li, X.; Lu, Y.; Jin, D.; Yin, X.; Jiang, H.; Huang, J.; Xiong, H.; et al. PRMT1 Is a Novel Molecular Therapeutic Target for Clear Cell Renal Cell Carcinoma. Theranostics 2021, 11, 5387–5403. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Shangguan, W.; Chen, Z.; Zheng, Z.; Chen, Y.; Zhong, Q.; Zhang, Y.; Yang, J.; Zhu, D.; Xie, W. Establishment of Sunitinib-Resistant Xenograft Model of Renal Cell Carcinoma and the Identification of Drug-Resistant Hub Genes and Pathways. Drug Des. Dev. Ther. 2021, 15, 5061–5074. [Google Scholar] [CrossRef]
- Verhoest, G. Développement et Mise au Point de Modèles Murins de Xénogreffe de Carcinome Rénal à Cellules Claires, et Evaluation de la Réponse de l’Association d’un Antagoniste des Récepteurs à l’Angiotensine-II au Sunitinib. Ph.D. Thesis, University of Rennes, Rennes, France, 2014; 96p. [Google Scholar]
- Yorozu, A.; Yamamoto, E.; Niinuma, T.; Tsuyada, A.; Maruyama, R.; Kitajima, H.; Numata, Y.; Kai, M.; Sudo, G.; Kubo, T.; et al. Upregulation of Adipocyte Enhancer-binding Protein 1 in Endothelial Cells Promotes Tumor Angiogenesis in Colorectal Cancer. Cancer Sci. 2020, 111, 1631–1644. [Google Scholar] [CrossRef]
- Hosseini-Beheshti, E.; Choi, W.; Weiswald, L.-B.; Kharmate, G.; Ghaffari, M.; Roshan-Moniri, M.; Hassona, M.D.; Chan, L.; Chin, M.Y.; Tai, I.T.; et al. Exosomes Confer Pro-Survival Signals to Alter the Phenotype of Prostate Cells in Their Surrounding Environment. Oncotarget 2016, 7, 14639–14658. [Google Scholar] [CrossRef] [Green Version]
- Gobin, E.; Bagwell, K.; Wagner, J.; Mysona, D.; Sandirasegarane, S.; Smith, N.; Bai, S.; Sharma, A.; Schleifer, R.; She, J.-X. A Pan-Cancer Perspective of Matrix Metalloproteases (MMP) Gene Expression Profile and Their Diagnostic/Prognostic Potential. BMC Cancer 2019, 19, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Kang, H.; Gao, M.; Jin, L.; Zhang, F.; Chen, D.; Li, M.; Xiao, L. Exosome-Transmitted Circ_MMP2 Promotes Hepatocellular Carcinoma Metastasis by Upregulating MMP2. Mol. Oncol. 2020, 14, 1365–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courcier, J.; de la Taille, A.; Nourieh, M.; Leguerney, I.; Lassau, N.; Ingels, A. Carbonic Anhydrase IX in Renal Cell Carcinoma, Implications for Disease Management. Int. J. Mol. Sci. 2020, 21, E7146. [Google Scholar] [CrossRef] [PubMed]
- Takacova, M.; Bartosova, M.; Skvarkova, L.; Zatovicova, M.; Vidlickova, I.; Csaderova, L.; Barathova, M.; Breza, J.; Bujdak, P.; Pastorek, J.; et al. Carbonic Anhydrase IX Is a Clinically Significant Tissue and Serum Biomarker Associated with Renal Cell Carcinoma. Oncol. Lett. 2013, 5, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.-J.; Rho, S.B.; Jung, D.C.; Han, I.-O.; Oh, E.-S.; Kim, J.-Y. Carbonic Anhydrase IX (CA9) Modulates Tumor-Associated Cell Migration and Invasion. J. Cell Sci. 2011, 124, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Ji, X.; Liu, J.; Fan, D.; Zhou, Q.; Chen, C.; Wang, W.; Wang, G.; Wang, H.; Yuan, W.; et al. Effects of Exosomes on Pre-Metastatic Niche Formation in Tumors. Mol. Cancer 2019, 18, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-Derived Exosomal MiR-1247-3p Induces Cancer-Associated Fibroblast Activation to Foster Lung Metastasis of Liver Cancer. Nat. Commun. 2018, 9, 191. [Google Scholar] [CrossRef] [Green Version]
- Horie, K.; Kawakami, K.; Fujita, Y.; Sugaya, M.; Kameyama, K.; Mizutani, K.; Deguchi, T.; Ito, M. Exosomes Expressing Carbonic Anhydrase 9 Promote Angiogenesis. Biochem. Biophys. Res. Commun. 2017, 492, 356–361. [Google Scholar] [CrossRef]
- Urabe, F.; Kosaka, N.; Kimura, T.; Egawa, S.; Ochiya, T. Extracellular Vesicles: Toward a Clinical Application in Urological Cancer Treatment. Int. J. Urol. 2018, 25, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Yoshioka, Y.; Fujita, Y.; Ochiya, T. Versatile Roles of Extracellular Vesicles in Cancer. J. Clin. Investig. 2016, 126, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral Sphingomyelinase 2 (NSMase2)-Dependent Exosomal Transfer of Angiogenic MicroRNAs Regulate Cancer Cell Metastasis. J. Biol. Chem. 2013, 288, 10849–10859. [Google Scholar] [CrossRef] [Green Version]
- Nishida-Aoki, N.; Tominaga, N.; Takeshita, F.; Sonoda, H.; Yoshioka, Y.; Ochiya, T. Disruption of Circulating Extracellular Vesicles as a Novel Therapeutic Strategy against Cancer Metastasis. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 181–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5 Patients | |
|---|---|
| Gender | |
| 2 |
| 3 |
| Medianage (Year, SD) | 63.8 (12.4) |
| TStage | |
| 1 |
| 2 |
| 2 |
| 0 |
| 0 |
| 1 |
| Mediantumorsize (cm, SD) | 8.6 (1.7) |
| ISUPscore | |
| 2 |
| 3 |
| Tumoralnecrosis | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferragu, M.; Vergori, L.; Le Corre, V.; Bellal, S.; del Carmen Martinez, M.; Bigot, P. Effects of Large Extracellular Vesicles from Kidney Cancer Patients on the Growth and Environment of Renal Cell Carcinoma Xenografts in a Mouse Model. Curr. Issues Mol. Biol. 2023, 45, 2491-2504. https://doi.org/10.3390/cimb45030163
Ferragu M, Vergori L, Le Corre V, Bellal S, del Carmen Martinez M, Bigot P. Effects of Large Extracellular Vesicles from Kidney Cancer Patients on the Growth and Environment of Renal Cell Carcinoma Xenografts in a Mouse Model. Current Issues in Molecular Biology. 2023; 45(3):2491-2504. https://doi.org/10.3390/cimb45030163
Chicago/Turabian StyleFerragu, Matthieu, Luisa Vergori, Vincent Le Corre, Sarah Bellal, Maria del Carmen Martinez, and Pierre Bigot. 2023. "Effects of Large Extracellular Vesicles from Kidney Cancer Patients on the Growth and Environment of Renal Cell Carcinoma Xenografts in a Mouse Model" Current Issues in Molecular Biology 45, no. 3: 2491-2504. https://doi.org/10.3390/cimb45030163
APA StyleFerragu, M., Vergori, L., Le Corre, V., Bellal, S., del Carmen Martinez, M., & Bigot, P. (2023). Effects of Large Extracellular Vesicles from Kidney Cancer Patients on the Growth and Environment of Renal Cell Carcinoma Xenografts in a Mouse Model. Current Issues in Molecular Biology, 45(3), 2491-2504. https://doi.org/10.3390/cimb45030163