
Sucrose Transporter StSUT2 Affects Potato Plants Growth, Flowering Time, and Tuber Yield
Abstract
:1. Introduction
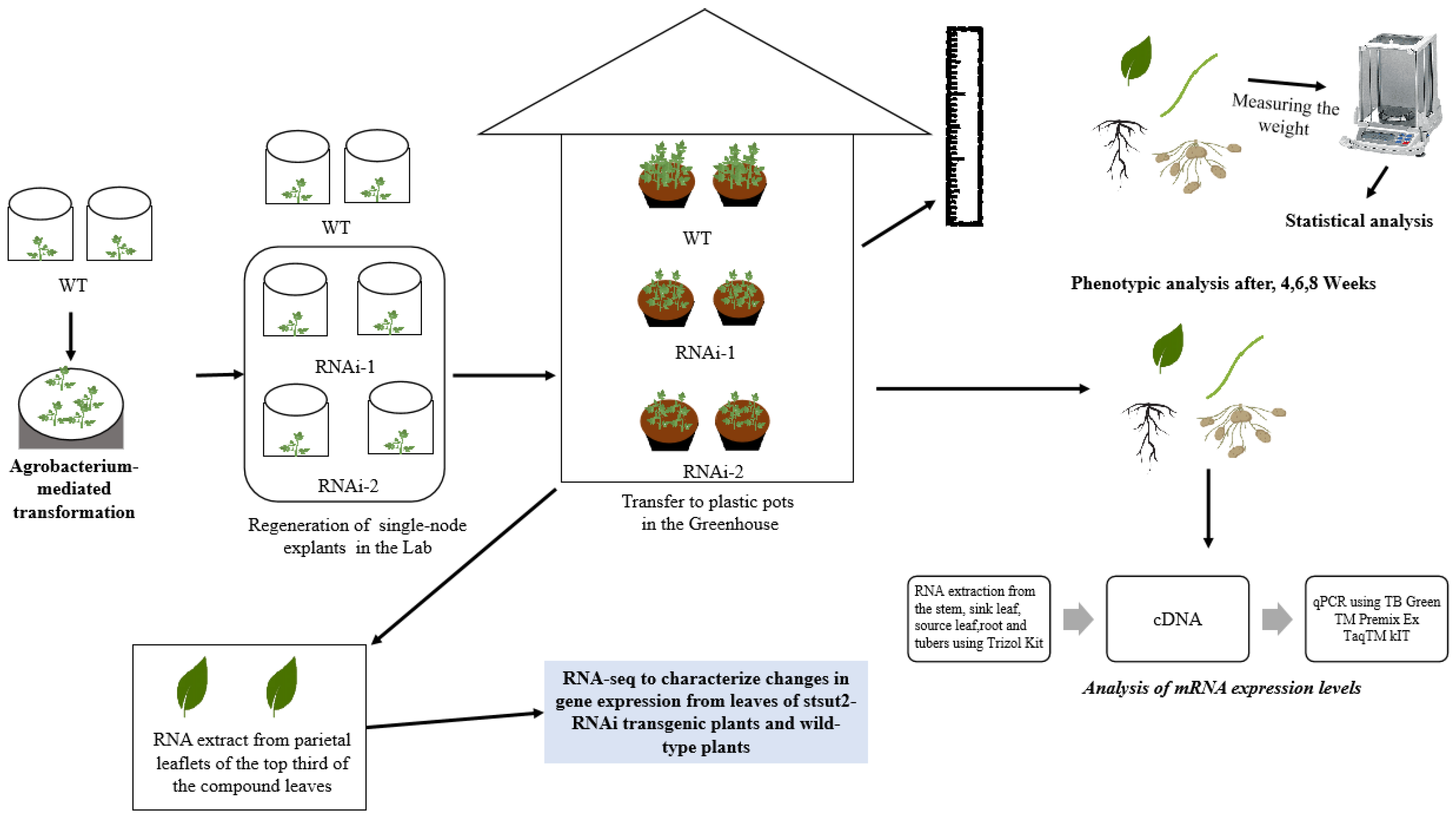
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Determination of Phenotypic Analysis of StSUT2-RNAi Plants
2.3. Analysis of Potato Yield
2.4. RNA-Seq
2.5. Analysis of mRNA Expression Levels
2.6. Statistical Analysis
3. Results
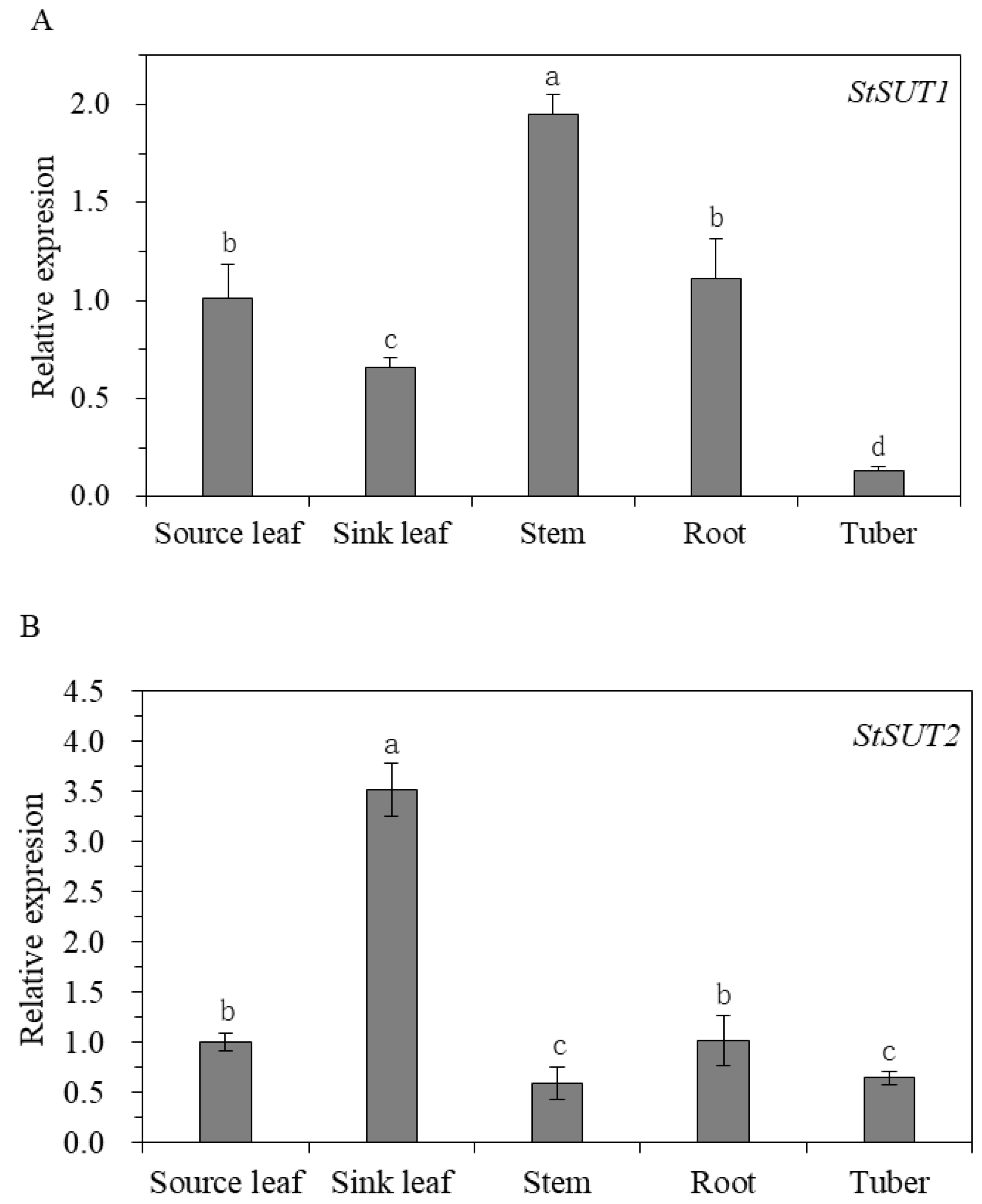
3.1. Gene Expression of StSUT1, StSUT2, and StSUT4 in Different Tissues
3.2. Phenotypic Analysis of StSUT2-RNA Interference Plants
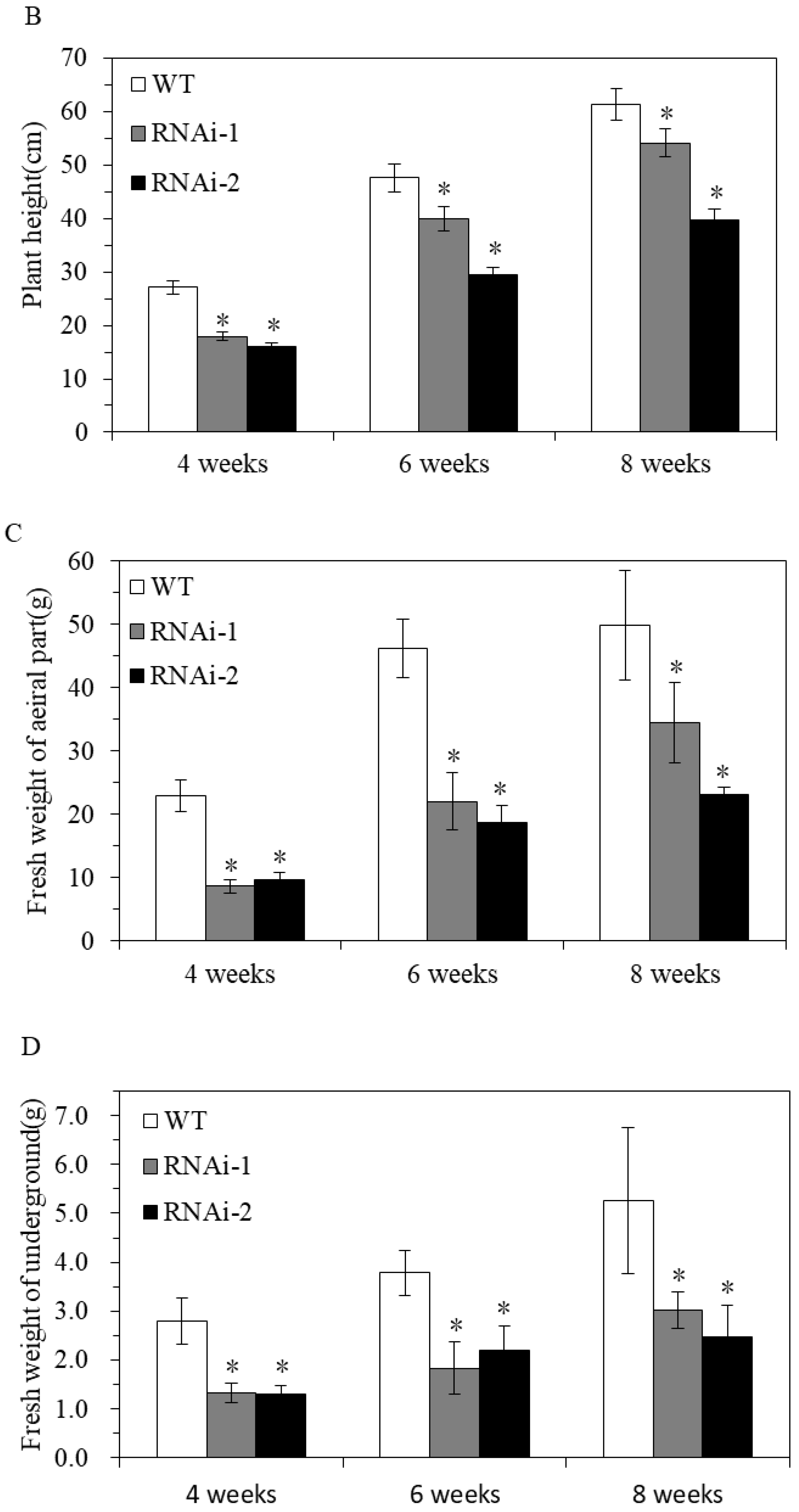
3.2.1. Effect of StSUT2-RNA Interference on Potato Vegetative Growth
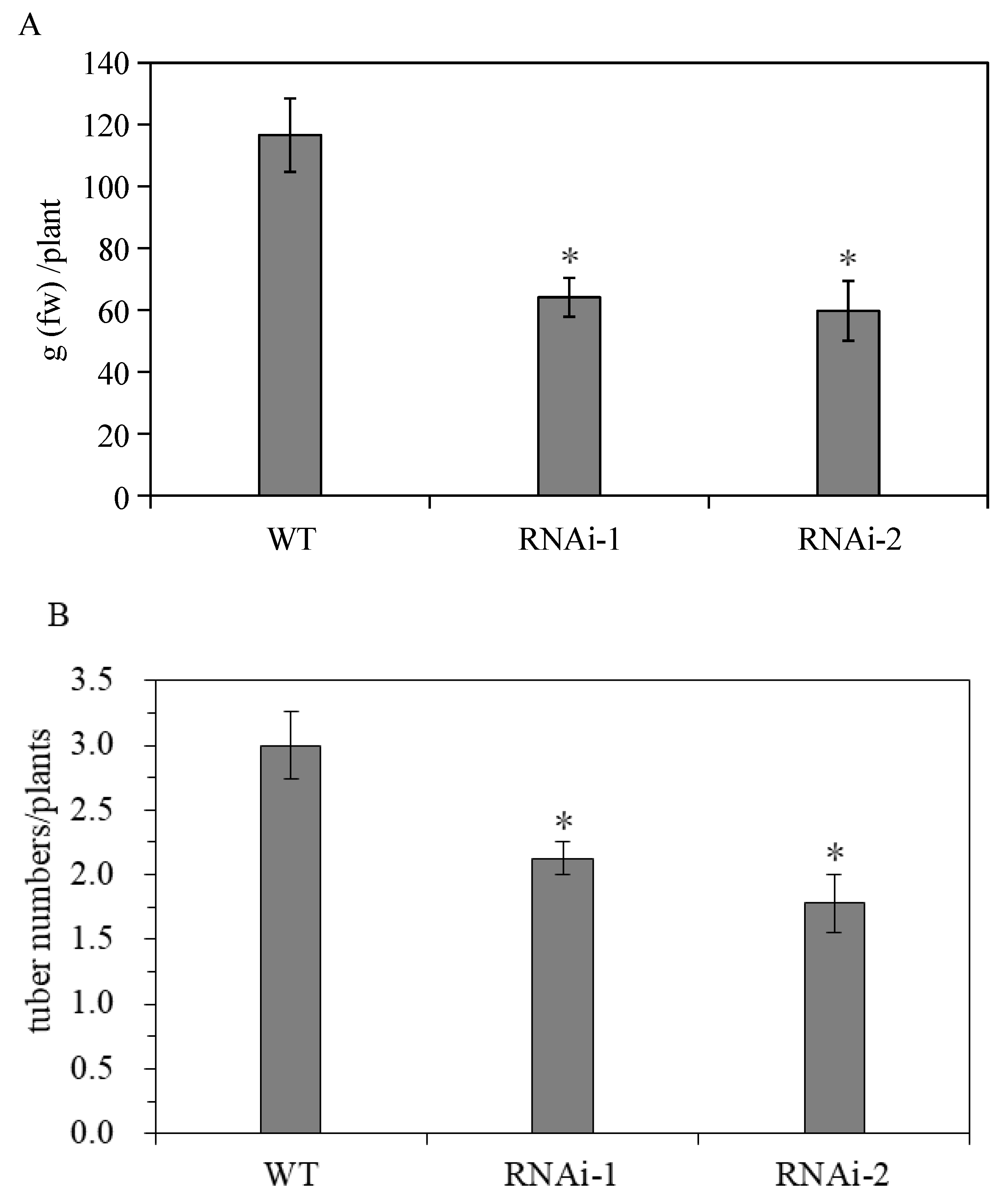
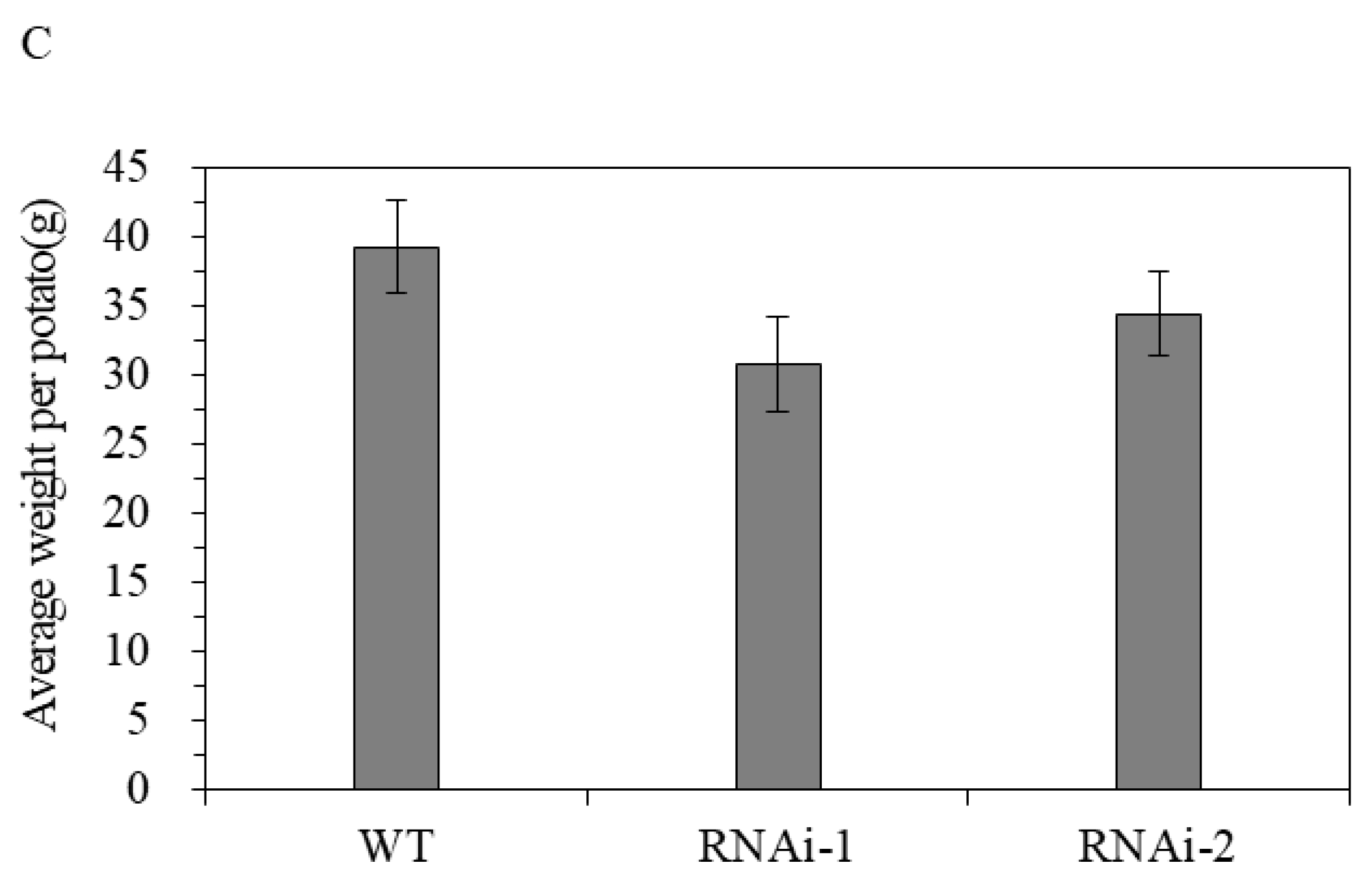
3.2.2. Effect of StSUT2-RNAi on Flowering and Tuber Yield
3.2.3. Effect of StSTU2-RNAi on Carbohydrate Content
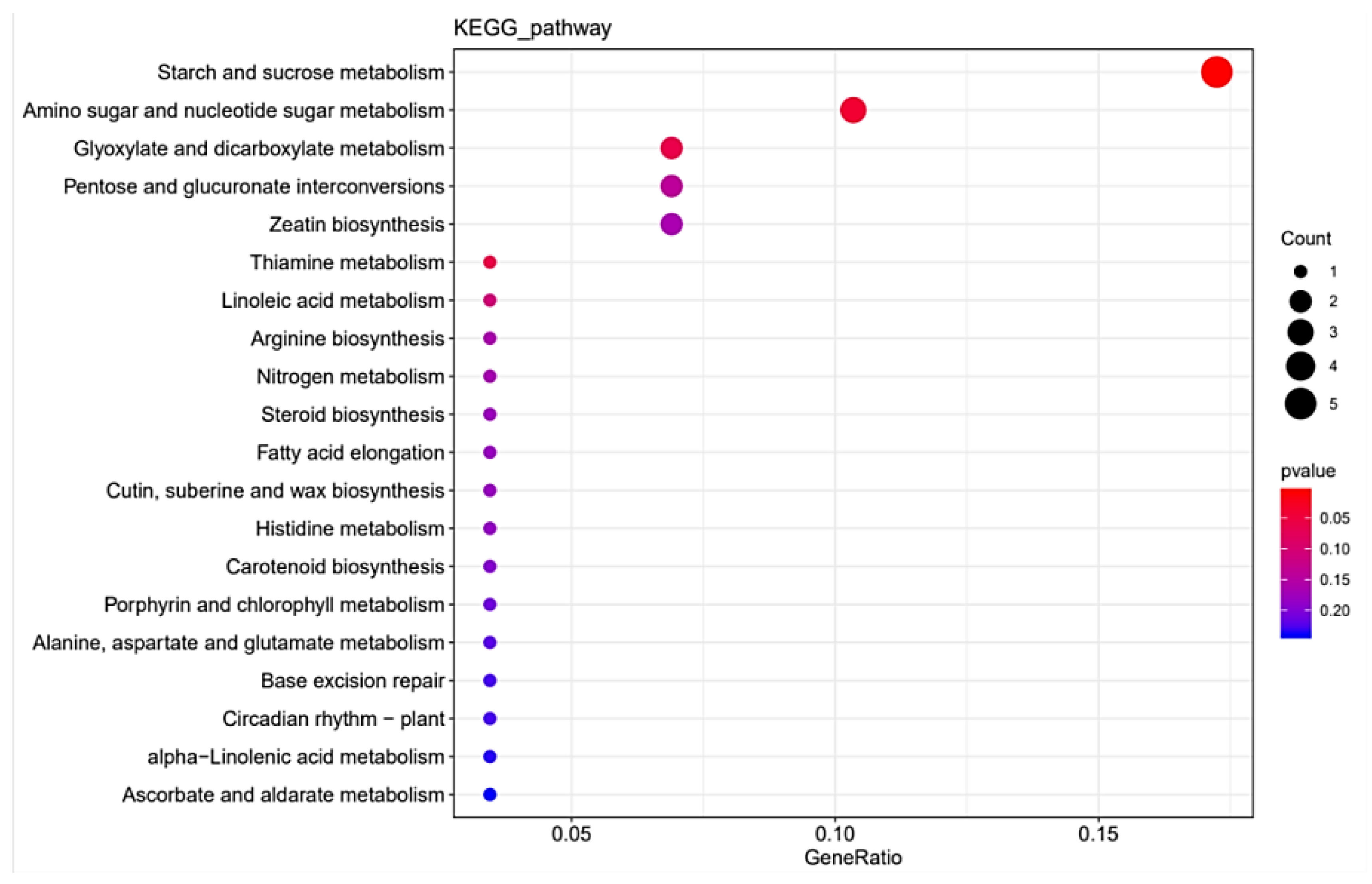
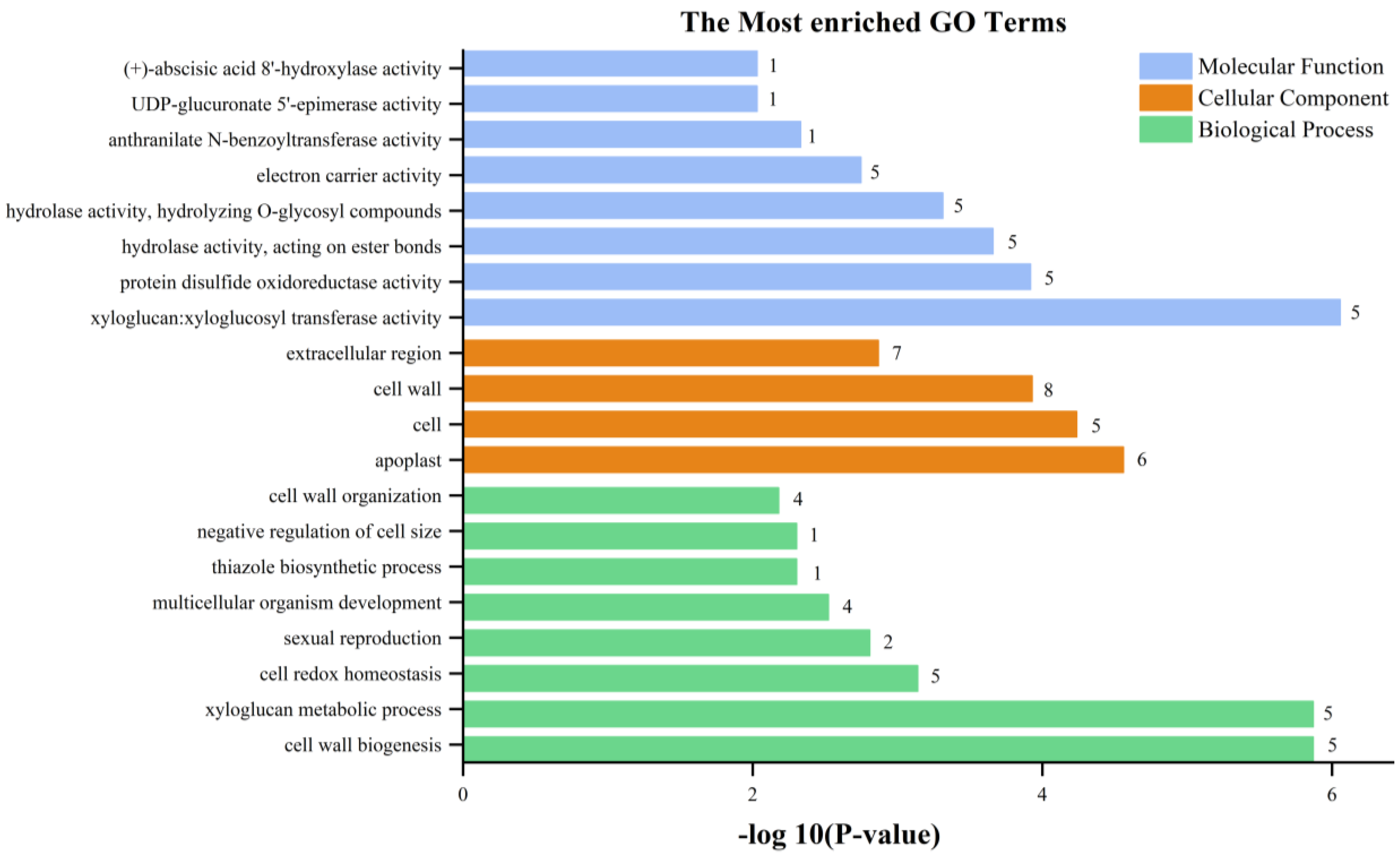
3.3. RNA-Seq Transcriptome Analysis between StSUT2-RNAi and WT Plants
4. Discussion
4.1. StSUT2 Is Involved in Different Physiological Characteristics of Potato Plants
4.2. Functional Annotation of Differentially Expressed Genes (DEGs)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lipavsk, H.; Ševčíková, H.; Mašková, P.; Tarkowská, D.; Mašek, T.; Lipavská, H.; Lipavsk, H. Carbohydrates and Gibberellins Relationship in Potato Tuberization. J. Plant Physiol. 2017, 214, 53–63. [Google Scholar] [CrossRef]
- Patrick, J.W.; Botha, F.C.; Birch, R.G. Metabolic Engineering of Sugars and Simple Sugar Derivatives in Plants. Plant Biotechnol. J. 2013, 11, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Kühn, C. A Comparison of the Sucrose Transporter Systems of Different Plant Species. Plant Biol. 2003, 5, 215–232. [Google Scholar] [CrossRef]
- Garg, V.; Kühn, C. Subcellular Dynamics and Protein-Protein Interactions of Plant Sucrose Transporters. J. Plant Physiol. 2022, 273, 153696. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, S.; Yunjuan, R.; Chen, S.; Liesche, J. Regulation of Sucrose Transporters and Phloem Loading in Response to Environmental Cues. Plant Physiol. 2018, 176, 930–945. [Google Scholar] [CrossRef] [Green Version]
- Chincinska, I.A.; Liesche, J.; Krügel, U.; Michalska, J.; Geigenberger, P.; Grimm, B.; Kühn, C.; Krügel, U.; Michalska, J.; Geigenberger, P.; et al. Sucrose Transporter StSUT4 from Potato Affects Flowering, Tuberization, and Shade Avoidance Response. Plant Physiol. 2008, 146, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Kühn, C.; Grof, C.P. Sucrose Transporters of Higher Plants. Curr. Opin. Plant Biol. 2010, 13, 287–297. [Google Scholar] [CrossRef]
- Sun, Y.; Reinders, A.; LaFleur, K.R.; Mori, T.; Ward, J.M. Transport Activity of Rice Sucrose Transporters OsSUT1 and OsSUT5. Plant Cell Physiol. 2010, 51, 114–122. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Chincinska, I.; Hackel, A.; Grimm, B.; Kuhn, C. Phloem Mobility and Stability of Sucrose Transporter Transcripts. Open Plant Sci. J. 2008, 2, 1–14. [Google Scholar] [CrossRef]
- Chincinska, I.; Gier, K.; Krügel, U.; Liesche, J.; He, H.; Grimm, B.; Harren, F.J.M.; Cristescu, S.M.; Kühn, C. Photoperiodic Regulation of the Sucrose Transporter StSUT4 Affects the Expression of Circadian-Regulated Genes and Ethylene Production. Front. Plant Sci. 2013, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.-H.; Lee, Y.-I.; Hsieh, S.-Y.; Lin, I.-S.; Wu, Y.-C.; Ko, H.-Y.; Klemens, P.A.; Neuhaus, H.E.; Chen, Y.-M.; Huang, T.-P.; et al. GeSUT4 Mediates Sucrose Import at the Symbiotic Interface for Carbon Allocation of Heterotrophic Gastrodia Elata (Orchidaceae). Plant. Cell Environ. 2021, 44, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Payyavula, R.S.; Tay, K.H.C.; Tsai, C.-J.; Harding, S.A. The Sucrose Transporter Family in Populus: The Importance of a Tonoplast PtaSUT4 to Biomass and Carbon Partitioning. Plant J. 2011, 65, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Y.; Han, H.; Ren, C.; Liu, S.; Long, J.; Peng, C. Cloning, Expression, and Localization Analysis of the Sucrose Transporter Gene JcSUT4 in the Biofuel Plant Jatropha Curcas. J. For. Res. 2018, 23, 297–303. [Google Scholar] [CrossRef]
- Wang, L.-F.; Qi, X.-X.; Huang, X.-S.; Xu, L.-L.; Jin, C.; Wu, J.; Zhang, S.-L. Overexpression of Sucrose Transporter Gene PbSUT2 from Pyrus Bretschneideri, Enhances Sucrose Content in Solanum Lycopersicum Fruit. Plant Physiol. Biochem. 2016, 105, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Hackel, A.; Schauer, N.; Carrari, F.; Fernie, A.R.; Grimm, B.; Kühn, C. Sucrose Transporter LeSUT1 and LeSUT2 Inhibition Affects Tomato Fruit Development in Different Ways. Plant J. 2006, 45, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Wang, J. Potato and Food Security in China. Am. J. Potato Res. 2019, 96, 100–101. [Google Scholar] [CrossRef] [Green Version]
- Viola, R.; Roberts, A.G.; Haupt, S.; Gazzani, S.; Hancock, R.D.; Marmiroli, N.; Machray, G.C.; Oparka, K.J. Tuberization in Potato Involves a Switch from Apoplastic to Symplastic Phloem Unloading. Plant Cell 2001, 13, 385–398. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.L.; Dusengemungu, L.; Igiraneza, C.; Rukundo, P. Molecular Regulation of Potato Tuber Dormancy and Sprouting: A Mini-Review. Plant Biotechnol. Rep. 2021, 15, 417–434. [Google Scholar] [CrossRef]
- Plantenga, F.D.M.; Bergonzi, S.; Bachem, C.W.B.; Visser, R.G.F.; Heuvelink, E.; Marcelis, L.F.M. High Light Accelerates Potato Flowering Independently of the FT-like Flowering Signal StSP3D. Environ. Exp. Bot. 2019, 160, 35–44. [Google Scholar] [CrossRef]
- Kühn, C.; Hajirezaei, M.-R.; Fernie, A.R.; Roessner-Tunali, U.; Czechowski, T.; Hirner, B.; Frommer, W.B. The Sucrose Transporter StSUT1 Localizes to Sieve Elements in Potato Tuber Phloem and Influences Tuber Physiology and Development. Plant Physiol. 2003, 131, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Horacio, P.; Martinez-Noel, G. Sucrose Signaling in Plants: A World yet to Be Explored. Plant Signal. Behav. 2013, 8, e23316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wu, L.; Foster, R.; Ruan, Y.-L. Molecular Regulation of Sucrose Catabolism and Sugar Transport for Development, Defence and Phloem Function. J. Integr. Plant Biol. 2017, 59, 322–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X. Construction of RNA Interference Vector of Potato Sucrose Transporter 2 and Its Genetic Transformation; Lanzhou University of Technology: Lanzhou, China, 2017. [Google Scholar]
- Gong, H.-L.; Chen, Q.-Q. Exogenous Sucrose Protects Potato Seedlings Against Heat Stress by Enhancing the Antioxidant Defense System. J. Soil Sci. Plant Nutr. 2021, 21, 1511–1519. [Google Scholar] [CrossRef]
- Mason, G.; Caciagli, P.; Accotto, G.P.; Noris, E. Real-Time PCR for the Quantitation of Tomato Yellow Leaf Curl Sardinia Virus in Tomato Plants and in Bemisia Tabaci. J. Virol. Methods 2008, 147, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Barker, L.; Kühn, C.; Weise, A.; Schulz, A.; Gebhardt, C.; Hirner, B.; Hellmann, H.; Schulze, W.; Ward, J.M.; Frommer, W.B. SUT2, a Putative Sucrose Sensor in Sieve Elements. Plant Cell 2000, 12, 1153–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansch, F.; Jaspar, H.; von Sivers, L.; Bitterlich, M.; Franken, P.; Kühn, C. Brassinosteroids and Sucrose Transport in Mycorrhizal Tomato Plants. Plant Signal. Behav. 2020, 15, 1714292. [Google Scholar] [CrossRef] [Green Version]
- Eom, J.-S.; Cho, J.-I.; Reinders, A.; Lee, S.-W.; Yoo, Y.; Tuan, P.Q.; Choi, S.-B.; Bang, G.; Park, Y.-I.; Cho, M.-H.; et al. Impaired Function of the Tonoplast-Localized Sucrose Transporter in Rice, OsSUT2, Limits the Transport of Vacuolar Reserve Sucrose and Affects Plant Growth. Plant Physiol. 2011, 157, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Bitterlich, M.; Krügel, U.; Boldt-Burisch, K.; Franken, P.; Kühn, C. The Sucrose Transporter SlSUT2 from Tomato Interacts with Brassinosteroid Functioning and Affects Arbuscular Mycorrhiza Formation. Plant J. 2014, 78, 877–889. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Annotation | RNA-Seq Fold Change | Regulation | RT-PCR Fold Change |
|---|---|---|---|---|
| PGSC0003DMG400021877 | Xyloglucan endotransglucosylase/hydrolase 2 | 3.11 | up | 1.98 |
| PGSC0003DMG400026189 | Xyloglucan endotransglucosylase/hydrolase protein 9 | 3.01 | up | 2.09 |
| PGSC0003DMG400004670 | Xyloglucan endotransglucosylase/hydrolase 2 | 2.72 | down | |
| PGSC0003DMG400000408 | Xyloglucan endotransglucosylase/hydrolase protein 31 | 2.66 | up | |
| PGSC0003DMG400004109 | Xyloglucan endotransglucosylase/hydrolase protein 33 | 2.99 | up | 1.47 |
| PGSC0003DMG400031731 | ω-hydroxypalmitate O-feruloyl transferase | 2.47 | down | |
| PGSC0003DMG400016249 | ω-hydroxypalmitate O-feruloyl transferase | 3.12 | up | |
| PGSC0003DMG400020589 | UDP-glucuronate 4-epimerase 6 | 2.35 | up | |
| PGSC0003DMG400011222 | UDP-glucuronate 4-epimerase 1 | 2.54 | up | 1.49 |
| PGSC0003DMG400023732 | UDP-glucoronosyl and UDP-glucosyl transferase | 2.29 | down | 1.33 |
| PGSC0003DMG400029738 | β-xylosidase/alpha-L-arabinofuranosidase 2 | 2.17 | up | |
| PGSC0003DMG400000827 | Galacturonosyltransferase-like 3 | 2.21 | up | |
| PGSC0003DMG401024140 | Pectin acetylesterase 8-like | 2.33 | up | 1.23 |
| PGSC0003DMG400015933 | pectinesterase/pectinesterase inhibitor PPE8B-like | 2.33 | up | 1.74 |
| PGSC0003DMG400031816 | Pectinesterase/pectinesterase inhibitor 41 | 2.87 | up | 2.53 |
| PGSC0003DMG400026220 | Expansin-like B1 | 2.91 | down | |
| PGSC0003DMG400018635 | Expansin-like A1 | 2.68 | up | 1.15 |
| PGSC0003DMG400014408 | Fasciclin-like arabinogalactan protein 1 | 2.25 | up | |
| PGSC0003DMG402013388 | Classical arabinogalactan protein 4-like | 2.51 | down | |
| PGSC0003DMG401027116 | Laccase-12 | 3.05 | down | 2.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, H.-L.; Liu, J.-B.; Igiraneza, C.; Dusengemungu, L. Sucrose Transporter StSUT2 Affects Potato Plants Growth, Flowering Time, and Tuber Yield. Curr. Issues Mol. Biol. 2023, 45, 2629-2643. https://doi.org/10.3390/cimb45030172
Gong H-L, Liu J-B, Igiraneza C, Dusengemungu L. Sucrose Transporter StSUT2 Affects Potato Plants Growth, Flowering Time, and Tuber Yield. Current Issues in Molecular Biology. 2023; 45(3):2629-2643. https://doi.org/10.3390/cimb45030172
Chicago/Turabian StyleGong, Hui-Ling, Jin-Bao Liu, Clement Igiraneza, and Leonce Dusengemungu. 2023. "Sucrose Transporter StSUT2 Affects Potato Plants Growth, Flowering Time, and Tuber Yield" Current Issues in Molecular Biology 45, no. 3: 2629-2643. https://doi.org/10.3390/cimb45030172
APA StyleGong, H. -L., Liu, J. -B., Igiraneza, C., & Dusengemungu, L. (2023). Sucrose Transporter StSUT2 Affects Potato Plants Growth, Flowering Time, and Tuber Yield. Current Issues in Molecular Biology, 45(3), 2629-2643. https://doi.org/10.3390/cimb45030172