

Characteristics, Comparative Analysis, and Phylogenetic Relationships of Chloroplast Genomes of Cultivars and Wild Relatives of Eggplant (Solanum melongena)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. DNA Extraction, Genome Sequencing, and Assembly
2.3. Genome Annotation
2.4. IR Expansion and Contraction
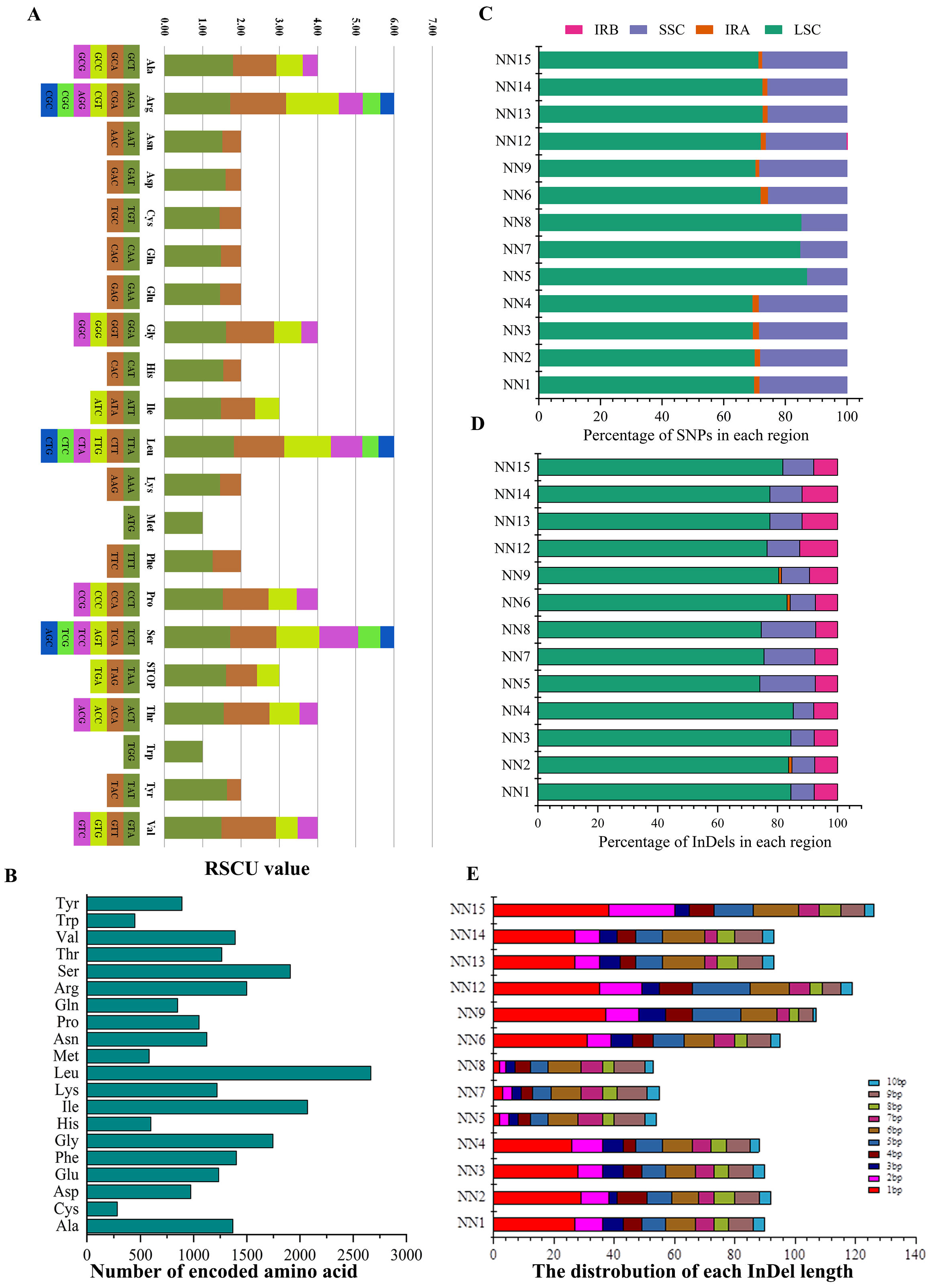
2.5. Codon Usage
2.6. Comparative Genome Analysis
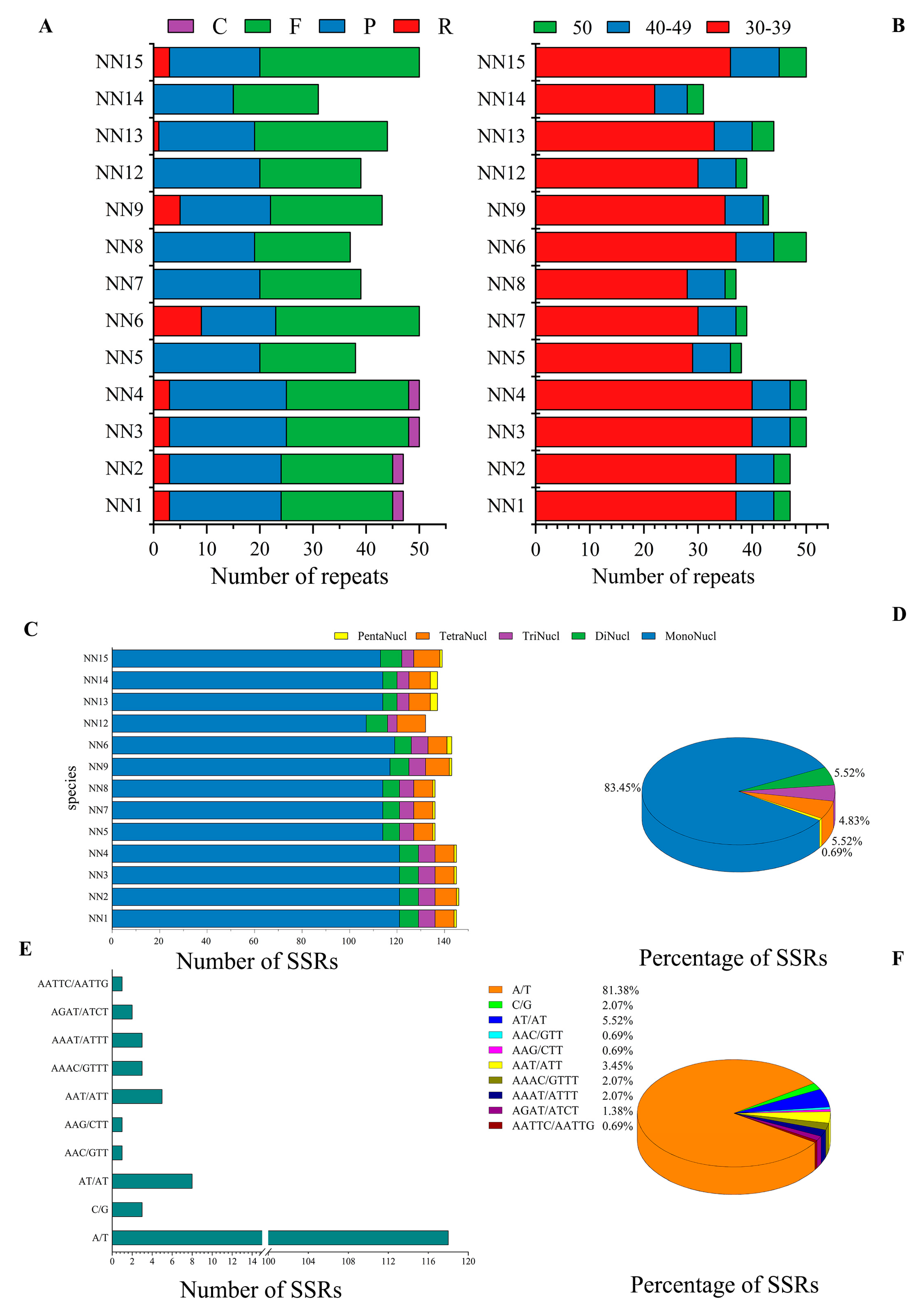
2.7. Repeat and SSR Analysis
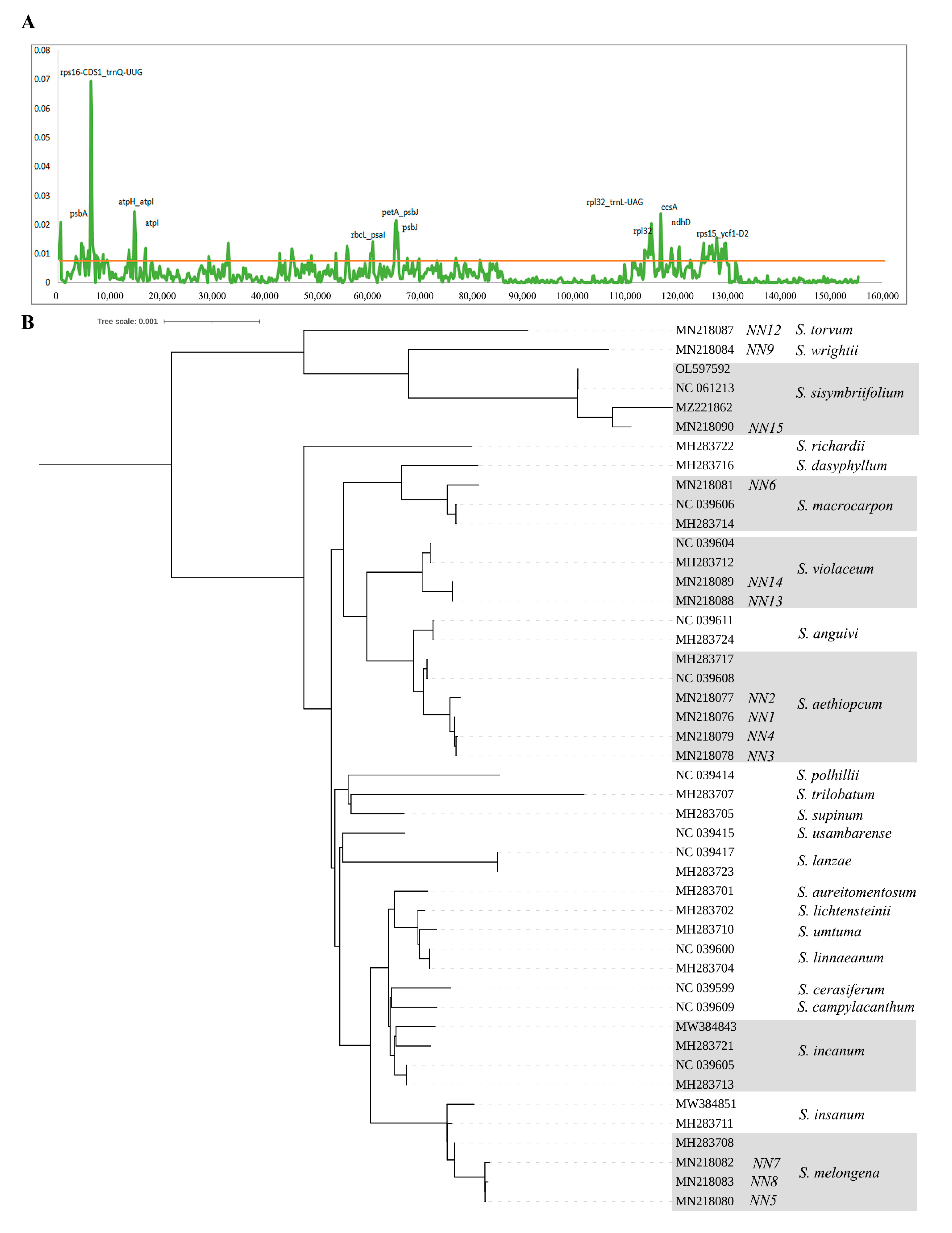
2.8. Identification of the Most Variable Regions
2.9. Phylogenetic Analysis
3. Results
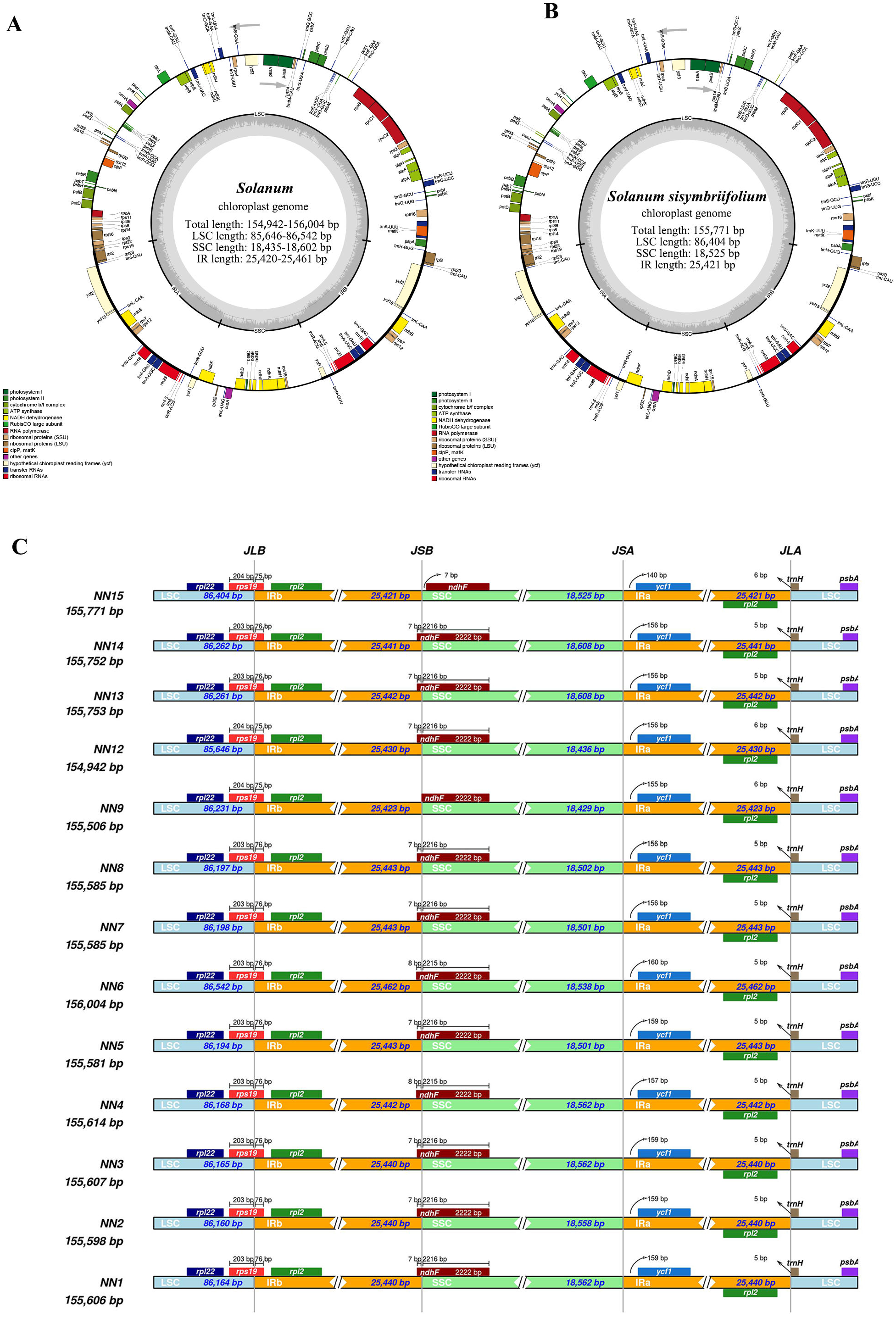
3.1. Characteristics of Cp Genomes of Leptostemonum Species
3.2. Comparative Genome Analysis
3.3. Repeat and SSR Analysis
3.4. Identification of the Most Variable Regions
3.5. Phylogenetic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vorontsova, M.S.; Stern, S.; Bohs, L.; Knapp, S. African spiny Solanum (subgenus Leptostemonum, Solanaceae): A thorny phylogenetic tangle. Bot. J. Linn. Soc. 2013, 173, 176–193. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, R.; Van Deynze, A.; Portis, E.; Rotino, G.L.; Toppino, L.; Hill, T.; Ashrafi, H.; Barchi, L.; Lanteri, S. New Insights on Eggplant/Tomato/Pepper Synteny and Identification of Eggplant and Pepper Orthologous QTL. Front. Plant Sci. 2016, 7, 1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Särkinen, T.; Bohs, L.; Olmstead, R.G.; Knapp, S. A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): A dated 1000-tip tree. BMC Evol. Biol. 2013, 13, 214. [Google Scholar] [CrossRef] [Green Version]
- Olmstead, R.G.; Bohs, L.; Migid, H.A.; Santiago-Valentin, E.; Garcia, V.F.; Collier, S.M. A molecular phylogeny of the Solanaceae. TAXON 2008, 57, 1159–1181. [Google Scholar] [CrossRef]
- Wu, F.; Tanksley, S.D. Chromosomal evolution in the plant family Solanaceae. BMC Genom. 2010, 11, 182. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Vorontsova, M.S.; Prohens, J. Wild Relatives of the Eggplant (Solanum melongena L.: Solanaceae): New Understanding of Species Names in a Complex Group. PLoS ONE 2013, 8, e57039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taher, D.; Solberg, S.; Prohens, J.; Chou, Y.-Y.; Rakha, M.; Wu, T.-H. World Vegetable Center Eggplant Collection: Origin, Composition, Seed Dissemination and Utilization in Breeding. Front. Plant Sci. 2017, 8, 1484. [Google Scholar] [CrossRef] [Green Version]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herraiz, F.J.; Ranil, R.; Fonseka, R.; Niran, L.; Fonseka, H.; et al. Interspecific Hybridization between Eggplant and Wild Relatives from Different Genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Afful, N.T.; Nyadanu, D.; Akromah, R.; Amoatey, H.M.; Annor, C.; Diawouh, R.G. Evaluation of crossability studies between selected eggplant accessions with wild relatives S. torvum, S. anguivi and S. aethopicum (Shum group). J. Plant Breed. Crop Sci. 2018, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Karihaloo, J.L.; Gottlieb, L.D. Allozyme variation in the eggplant, Solanum melongena L. (Solanaceae). Theor. Appl. Genet. 1995, 90, 578–583. [Google Scholar] [CrossRef]
- Ranil, R.H.G.; Prohens, J.; Aubriot, X.; Niran, H.M.L.; Plazas, M.; Fonseka, R.M.; Vilanova, S.; Fonseka, H.H.; Gramazio, P.; Knapp, S. Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): A review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet. Resour. Crop. Evol. 2016, 64, 1707–1722. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Juez, E.; Pyke, K. Plastids unleashed: Their development and their integration in plant development. Int. J. Dev. Biol. 2005, 49, 557–577. [Google Scholar] [CrossRef] [Green Version]
- Peeters, N.M.; Hanson, M.R.; Halter, C.P. RNA editing in ribosome-less plastids of iojap maize. Curr. Genet. 2004, 45, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; DePamphilis, C.W.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J.; et al. Methods for Obtaining and Analyzing Whole Chloroplast Genome Sequences. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2005; Volume 395, pp. 348–384. [Google Scholar]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Gao, H.; Wang, Y.; Song, J.; Henry, R.; Wu, H.; Hu, Z.; Yao, H.; Luo, H.; Luo, K.; et al. Complete chloroplast genome sequence of Magnolia grandiflora and comparative analysis with related species. Sci. China Life Sci. 2013, 56, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Sagona, E.; Carbonell, A.K.; Chiarini, F. A revision of the Solanum elaeagnifolium clade (Elaeagnifolium clade; subgenus Leptostemonum, Solanaceae). Phytokeys 2017, 84, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Aubriot, X.; Knapp, S.; Syfert, M.M.; Poczai, P.; Buerki, S. Shedding new light on the origin and spread of the brinjal eggplant (Solanum melongena L.) and its wild relatives. Am. J. Bot. 2018, 105, 1175–1187. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, S.; Manzur, J.P.; Prohens, J. Development and characterization of genomic simple sequence repeat markers in eggplant and their application to the study of diversity and relationships in a collection of different cultivar types and origins. Mol. Breed. 2011, 30, 647–660. [Google Scholar] [CrossRef]
- McPherson, H.; Van Der Merwe, M.; Delaney, S.K.; Edwards, M.A.; Henry, R.J.; McIntosh, E.; Rymer, P.D.; Milner, M.L.; Siow, J.; Rossetto, M. Capturing chloroplast variation for molecular ecology studies: A simple next generation sequencing approach applied to a rainforest tree. BMC Ecol. 2013, 13, 8. [Google Scholar] [CrossRef] [Green Version]
- Borgström, E.; Lundin, S.; Lundeberg, J. Large Scale Library Generation for High Throughput Sequencing. PLoS ONE 2011, 6, e19119. [Google Scholar] [CrossRef] [Green Version]
- Cronn, R.; Liston, A.; Parks, M.; Gernandt, D.S.; Shen, R.; Mockler, T. Multiplex sequencing of plant chloroplast genomes using Solexa sequencing-by-synthesis technology. Nucleic Acids Res. 2008, 36, e122. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Magrane, M. Consortium UniProt Knowledgebase: A hub of integrated protein data. Database: J. Biol. Databases Curation 2011, 2011, bar009. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Makałowska, I. ChloroplastDB: The Chloroplast Genome Database. Nucleic Acids Res. 2006, 34, D692–D696. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Ogban, F.; Udensi, O.; Inyang, G.; Osang, F. Diversity in Content and Analytical Algorithms of Biologist-Centric Software(s) for DNA and Sequence Data: Mega, DNASp, GenAlex and ARLEQUIN. Int. J. Nat. Appl. Sci. 2019, 12, 47–60. [Google Scholar]
- Katoh, K.; Kuma, K.I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Ding, Q.-X.; Liu, J.; Gao, L.-Z. The complete chloroplast genome of eggplant (Solanum melongena L.). Mitochondrial DNA Part B 2016, 1, 843–844. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Gan, G.; Li, W.; Li, W.; Jiang, Y.; Liang, X.; Yu, N.; Chen, R.; Wang, Y. Inheritance of Solanum chloroplast genomic DNA in interspecific hybrids. Mitochondrial DNA Part B 2021, 6, 351–357. [Google Scholar] [CrossRef]
- Yin, M.; Yu, Y.; Gong, Y.; Gui, M.; Li, Z.; Bao, R.; Cheng, J.; Du, G.; Wu, L. The complete chloroplast genome of Solanum sisymbriifolium (Solanaceae), the wild eggplant. Mitochondrial DNA Part B 2022, 7, 886–888. [Google Scholar] [CrossRef]
- Devi, C.P.; Munshi, A.D.; Behera, T.K.; Choudhary, H.; Gurung, B.V.; Saha, P. Cross compatibility in interspecific hybridization of eggplant, Solanum melongena, with its wild relatives. Sci. Hortic. 2015, 193, 353–358. [Google Scholar] [CrossRef]
- Prohens, J.; Plazas, M.; Raigón, M.D.; Seguí-Simarro, J.M.; Stommel, J.R.; Vilanova, S. Characterization of interspecific hybrids and first backcross generations from crosses between two cultivated eggplants (Solanum melongena and S. aethiopicum Kumba group) and implications for eggplant breeding. Euphytica 2012, 186, 517–538. [Google Scholar] [CrossRef]
- Bletsos, F.; Roupakias, D.; Tsaktsira, M.; Scaltsoyjannes, A. Production and characterization of interspecific hybrids between three eggplant (Solanum melongena L.) cultivars and Solanum macrocarpon L. Sci. Hortic. 2004, 101, 11–21. [Google Scholar] [CrossRef]
- Stedje, B.; Bukenya-Ziraba, R. RAPD variation in Solanum anguivi Lam. and S. aethiopicum L. (Solanaceae) in Uganda. Euphytica 2003, 131, 293–297. [Google Scholar] [CrossRef]
- Hassan, S.M.; Lester, R.N. Origin and domestication of the Brinjal eggplant, Solanum melongena L., from Solanum incanum L., in Africa and Asia. In Proceedings of the III Congreso Mundial De Plantas Solanaceas International Solanaceae Congress, Bogota, Columbia, 25–30 July 1988; pp. 25–30. [Google Scholar]
- Anaso, H.U. Comparative cytological study of Solanum aethiopicum Gilo group, Solanum aethiopicum Shum group and Solanum anguivi. Euphytica 1991, 53, 81–85. [Google Scholar] [CrossRef]
- Rakha, M.; Prohens, J.; Taher, D.; Wu, T.-h.; Solberg, S.Ø. Advances in Plant Breeding Strategies: Vegetable Crops: Volume 9: Fruits and Young Shoots; Jameel, M., Al-Khayri, S., Mohan, J., Johnson, D.V., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 163–203. [Google Scholar]
- Bletsos, F.; Roupakias, D.; Tsaktsira, M.; Scaltsoyjannes, A.; Thanassoulopoulos, C.C. Interspecific hybrids between three eggplant (Solanum melongena L.) cultivars and two wild species (Solanum torvum Sw. and Solanum sisymbriifolium Lam.). Plant Breed. 1998, 117, 159–164. [Google Scholar] [CrossRef]
- Kumchai, J.; Wei, Y.-C.; Lee, C.-Y.; Chen, F.-C.; Chin, S.-W. Production of interspecific hybrids between commercial cultivars of the eggplant (Solanum melongena L.) and its wild relative S. torvum. Genet. Mol. Res.: GMR 2013, 12, 755–764. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | Code | Species | Gene Bank Accession Number | Origin |
|---|---|---|---|---|
| NN1 | 166 | Solanum aethiopicum L. | MN218076 | South Africa |
| NN2 | 53 | Solanum aethiopicum L. | MN218077 | Brazil |
| NN3 | 137 | Solanum aethiopicum L. | MN218078 | Ethiopia |
| NN4 | Y11 | Solanum aethiopicum L. | MN218079 | Ethiopia |
| NN5 | 177 | Solanum melongena L. | MN218080 | South China |
| NN6 | Lwpq | Solanum macrocapon L. | MN218081 | Laos |
| NN7 | Shf | Solanum melongena L. | MN218082 | Viet Nam |
| NN8 | 131 | Solanum melongena L. | MN218083 | South China |
| NN9 | Dhq | Solanum wrightii L. | MN218084 | South China |
| NN12 | Sq | Solanum torvum | MN218087 | South China |
| NN13 | Ctq | Solanum anguivi L. | MN218088 | Thailand |
| NN14 | ctq-B | Solanum anguivi L. | MN218089 | South China |
| NN15 | Sjq | Solanum sisymbriifolium | MN218090 | South China |
| Sample ID | Insert Size (bp) | Raw Data (Mb) | Clean Data (Mb) | Reads Length (bp) | Clean Data GC(%) | Clean Data Q30(%) | Average Organelle Depth |
|---|---|---|---|---|---|---|---|
| NN1 | 430 | 5981 | 5210 | (150:150) | 36.65 | 93.81 | 1452 |
| NN2 | 430 | 9111 | 8248 | (150:150) | 36.36 | 94.4 | 1521 |
| NN3 | 430 | 6754 | 6034 | (150:150) | 36.47 | 94.21 | 1336 |
| NN4 | 430 | 6374 | 5638 | (150:150) | 36.91 | 93.95 | 1824 |
| NN5 | 430 | 6515 | 5792 | (150:150) | 36.48 | 94.13 | 1599 |
| NN6 | 430 | 8670 | 8302 | (150:150) | 36.92 | 95.39 | 1477 |
| NN7 | 430 | 8584 | 8307 | (150:150) | 37.64 | 95.74 | 1581 |
| NN8 | 430 | 9893 | 9557 | (150:150) | 37.13 | 95.64 | 1798 |
| NN9 | 430 | 8504 | 8193 | (150:150) | 37.12 | 95.86 | 700 |
| NN12 | 430 | 10,165 | 9777 | (150:150) | 38.1 | 95.54 | 3639 |
| NN13 | 430 | 8408 | 8139 | (150:150) | 36.42 | 95.78 | 2337 |
| NN14 | 430 | 5781 | 5193 | (150:150) | 36.24 | 95.31 | 769 |
| NN15 | 430 | 5281 | 4503 | (150:150) | 39.27 | 93.5 | 636 |
| Sample ID | Total Length (bp) | LSC Length (bp) | SSC Length (bp) | IRA/IRB Length (bp) | Total GC Content (%) | Protein- Coding Gene Number | tRNA Number | rRNA Number | Total Gene Number |
|---|---|---|---|---|---|---|---|---|---|
| NN1 | 155,606 | 86,164 | 18,562 | 25,440 | 37.7 | 80 | 42 | 8 | 130 |
| NN2 | 155,598 | 86,160 | 18,558 | 25,440 | 37.7 | 80 | 42 | 8 | 130 |
| NN3 | 155,607 | 86,165 | 18,562 | 25,440 | 37.7 | 80 | 42 | 8 | 130 |
| NN4 | 155,614 | 86,168 | 18,564 | 25,441 | 37.7 | 80 | 42 | 8 | 130 |
| NN5 | 155,581 | 86,194 | 18,501 | 25,443 | 37.71 | 80 | 42 | 8 | 130 |
| NN6 | 156,004 | 86,542 | 18,538 | 25,462 | 37.61 | 80 | 42 | 8 | 130 |
| NN7 | 155,585 | 86,197 | 18,502 | 25,443 | 37.71 | 80 | 42 | 8 | 130 |
| NN8 | 155,585 | 86,197 | 18,502 | 25,443 | 37.71 | 80 | 42 | 8 | 130 |
| NN9 | 155,506 | 86,231 | 18,429 | 25,423 | 37.68 | 80 | 42 | 8 | 130 |
| NN12 | 154,942 | 85,646 | 18,436 | 25,430 | 37.81 | 80 | 42 | 8 | 130 |
| NN13 | 155,753 | 86,261 | 18,608 | 25,442 | 37.66 | 80 | 42 | 8 | 130 |
| NN14 | 155,752 | 86,262 | 18,608 | 25,441 | 37.67 | 80 | 42 | 8 | 130 |
| NN15 | 155,771 | 86,404 | 18,525 | 25,421 | 37.76 | 79 | 42 | 8 | 129 |
| Function | Gene Group | Gene Name |
|---|---|---|
| Photosynthesis pathways | Photosystem I | psa (A, B, C *, I, J) |
| Photosystem I assembly | ycf (3, 4) | |
| Photosystem II | psb (A-F, H-L, N, T, Z) | |
| F-type ATP synthase | atp (A, B, E, F, H, I) | |
| NDH complex | ndh (A *, B #, C, D *, E *, F *, G *, H *, I *, J, K) | |
| Component of cytochrome b6/f complex | pet (A, B, D, L) | |
| Inner envelope membrane | cemA | |
| Cytochrome biogenesis protein | ccsA * | |
| Large subunit of Rubisco | rbcL | |
| Structural RNAs | Transfer RNAs | trnH-GUG; trnK-UUU; trnQ-UUG; trnS-GCU; trnG-UCC; trnR-UCU; trnF-GAA; trnD-GUC; trnY-GUA; trnE-UUC; trnT-GGU; trnM-CAU; trnS-UGA; trnC-GCA; trnG-GCC; trnfM-CAU; trnS-GGA; trnT-UGU; trnL-UAA; trnI-CAU #; trnV-UAC; trnW-CCA; trnP-UGG; trnP-GGG; trnI-GAU #; trnA-UGC #; trnN-GUU#; trnL-UAG; trnR-ACG #; trnV-GAC #; trnL-CAA # |
| Ribosomal RNAs | rrn (4.5 #, 5 #, 16 #, 23 #) | |
| Genetic apparatus | Large subunit of ribosomal protein | rpl (2 #, 14, 16, 20, 22, 23 #, 32 *, 33, 36) |
| Small subunit of ribosomal protein | rps (2, 3, 4, 7 #, 8, 11, 12 #&, 14, 15 *, 16 0, 18, 19) | |
| Subunits of RNA polymerase | rpo (A, B, C1, C2) | |
| Post-transcriptional modification | Maturase | MatK |
| Protein- modifying | ATP- dependent Clp protease proteolytic subunit | clpP |
| Unknown Proteins | ycf (1 #, 2 #, 15) |
| Sample ID | Species | CDS InDel | Intergenic InDel | Insertion | Deletion | Total InDel | CDS SNPs | Intergenic_SNPs | ts/tv | Total_SNPs |
|---|---|---|---|---|---|---|---|---|---|---|
| NN1 | S. aethiopicum L. | 7 | 83 | 57 | 33 | 90 | 98 | 242 | 1.11 | 340 |
| NN2 | S. aethiopicum L. | 6 | 86 | 58 | 34 | 92 | 100 | 239 | 1.08 | 339 |
| NN3 | S. aethiopicum L. | 7 | 83 | 57 | 33 | 90 | 99 | 243 | 1.12 | 342 |
| NN4 | S. aethiopicum L. | 7 | 81 | 56 | 32 | 88 | 100 | 244 | 1.12 | 344 |
| NN5 | S. melongena L. | 8 | 46 | 46 | 8 | 54 | 9 | 30 | 0.44 | 39 |
| NN7 | S. melongena L. | 8 | 47 | 46 | 9 | 55 | 9 | 30 | 0.44 | 39 |
| NN8 | S. melongena L. | 8 | 45 | 45 | 8 | 53 | 10 | 31 | 0.44 | 41 |
| NN6 | S. melongena L. | 5 | 90 | 64 | 31 | 95 | 111 | 344 | 0.95 | 455 |
| NN9 | S. wrightii L. | 4 | 103 | 59 | 48 | 107 | 267 | 606 | 0.95 | 873 |
| NN12 | S. torvum | 5 | 114 | 72 | 47 | 119 | 243 | 599 | 0.91 | 842 |
| NN13 | S. indicum L. | 6 | 87 | 57 | 36 | 93 | 103 | 268 | 1.01 | 371 |
| NN14 | S. indicum L. | 6 | 87 | 57 | 36 | 93 | 103 | 267 | 1.01 | 370 |
| NN15 | S. sisymbriifolium | 5 | 121 | 76 | 50 | 126 | 259 | 637 | 0.94 | 896 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Li, Y.; Cai, L.; Gan, G.; Wang, P.; Li, W.; Li, W.; Jiang, Y.; Li, D.; Wang, M.; et al. Characteristics, Comparative Analysis, and Phylogenetic Relationships of Chloroplast Genomes of Cultivars and Wild Relatives of Eggplant (Solanum melongena). Curr. Issues Mol. Biol. 2023, 45, 2832-2846. https://doi.org/10.3390/cimb45040185
Yang Q, Li Y, Cai L, Gan G, Wang P, Li W, Li W, Jiang Y, Li D, Wang M, et al. Characteristics, Comparative Analysis, and Phylogenetic Relationships of Chloroplast Genomes of Cultivars and Wild Relatives of Eggplant (Solanum melongena). Current Issues in Molecular Biology. 2023; 45(4):2832-2846. https://doi.org/10.3390/cimb45040185
Chicago/Turabian StyleYang, Qihong, Ye Li, Liangyu Cai, Guiyun Gan, Peng Wang, Weiliu Li, Wenjia Li, Yaqin Jiang, Dandan Li, Mila Wang, and et al. 2023. "Characteristics, Comparative Analysis, and Phylogenetic Relationships of Chloroplast Genomes of Cultivars and Wild Relatives of Eggplant (Solanum melongena)" Current Issues in Molecular Biology 45, no. 4: 2832-2846. https://doi.org/10.3390/cimb45040185
APA StyleYang, Q., Li, Y., Cai, L., Gan, G., Wang, P., Li, W., Li, W., Jiang, Y., Li, D., Wang, M., Xiong, C., Chen, R., & Wang, Y. (2023). Characteristics, Comparative Analysis, and Phylogenetic Relationships of Chloroplast Genomes of Cultivars and Wild Relatives of Eggplant (Solanum melongena). Current Issues in Molecular Biology, 45(4), 2832-2846. https://doi.org/10.3390/cimb45040185