
Do Elevated YKL-40 Levels Drive the Immunosuppressive Tumor Microenvironment in Colorectal Cancer? Assessment of the Association of the Expression of YKL-40, MMP-8, IL17A, and PD-L1 with Coexisting Type 2 Diabetes, Obesity, and Active Smoking
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of Immune Cell Infiltration Scores and the Immune Landscape and Gene Set Enrichment Analysis (GSEA) in Colorectal Cancer
2.2. Gene Expression Omnibus (GEO) Dataset Analysis
2.3. Tissue Homogenates of CRC
2.4. Preparation of Samples
2.5. Evaluation of YKL-40, MMP8, IL17A, and PD-L1 Levels
2.6. Statistical Analysis
3. Results
3.1. GEO Profile Analysis
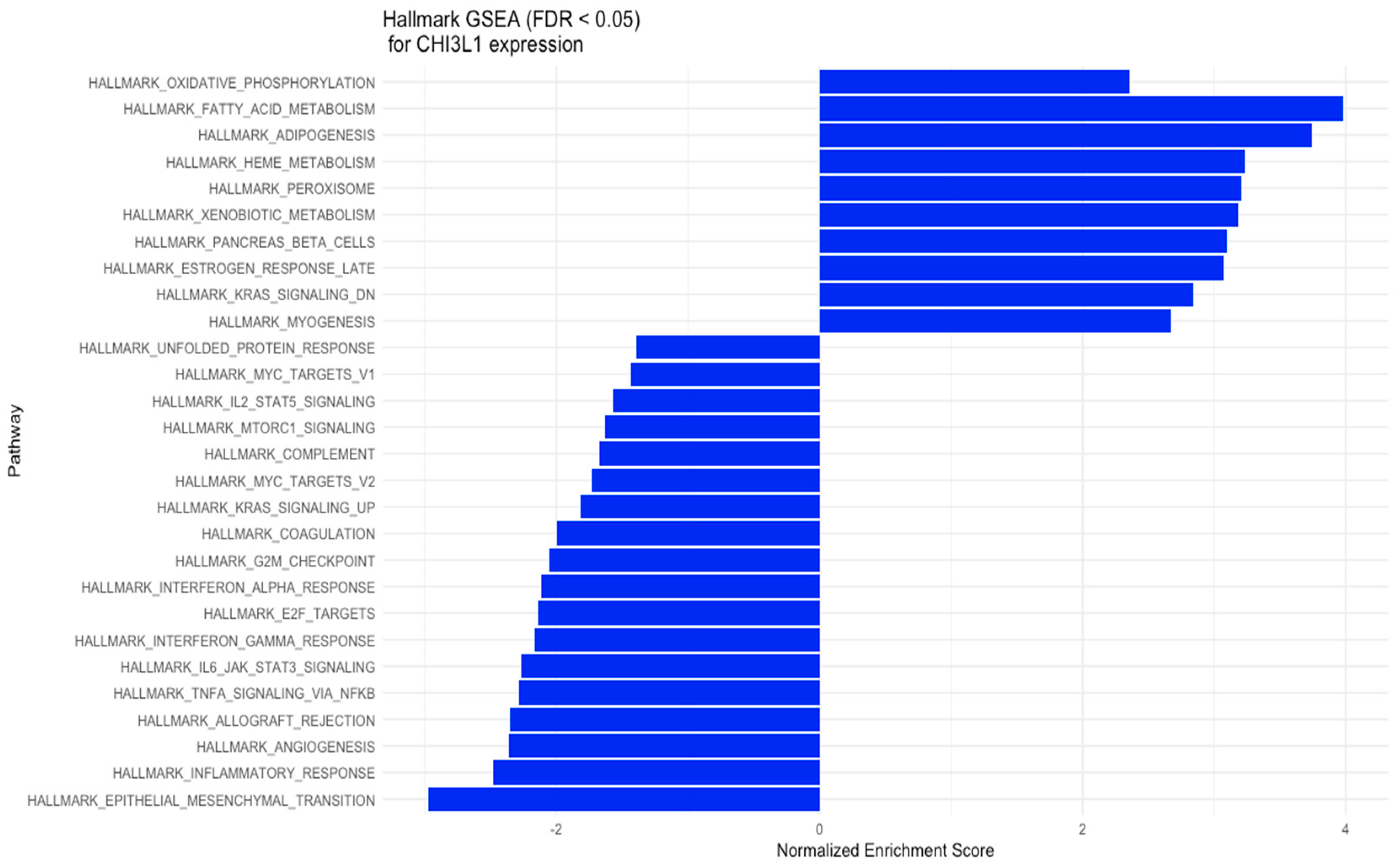
3.2. GSEA Results: Predicted Pathways and Biological Processes for High–Low YKL-40 Expression
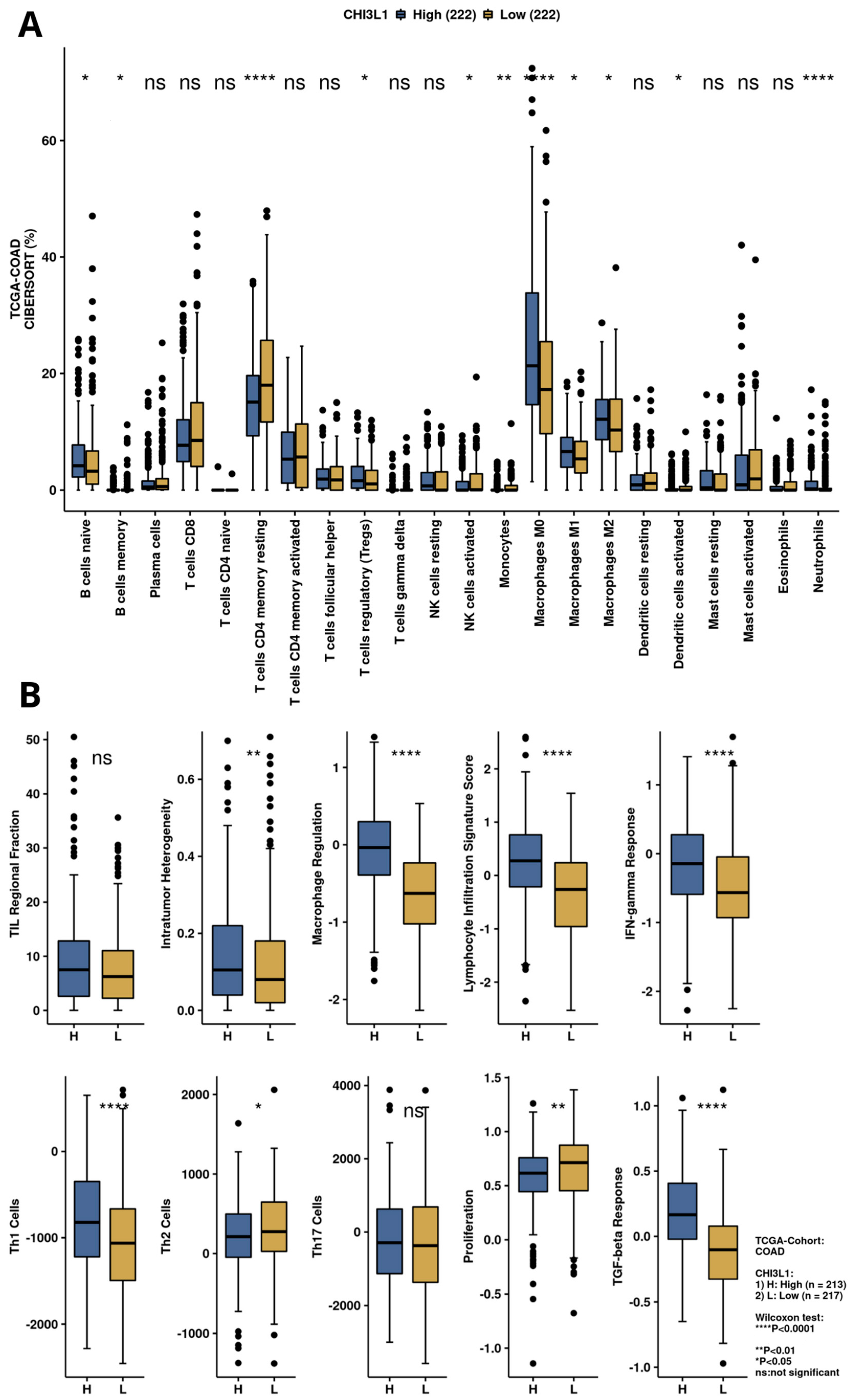
3.3. Exploration of Immunogenicty and the Immune Infiltration Landscape of YKL-40 Based on TCGA-COAD Data
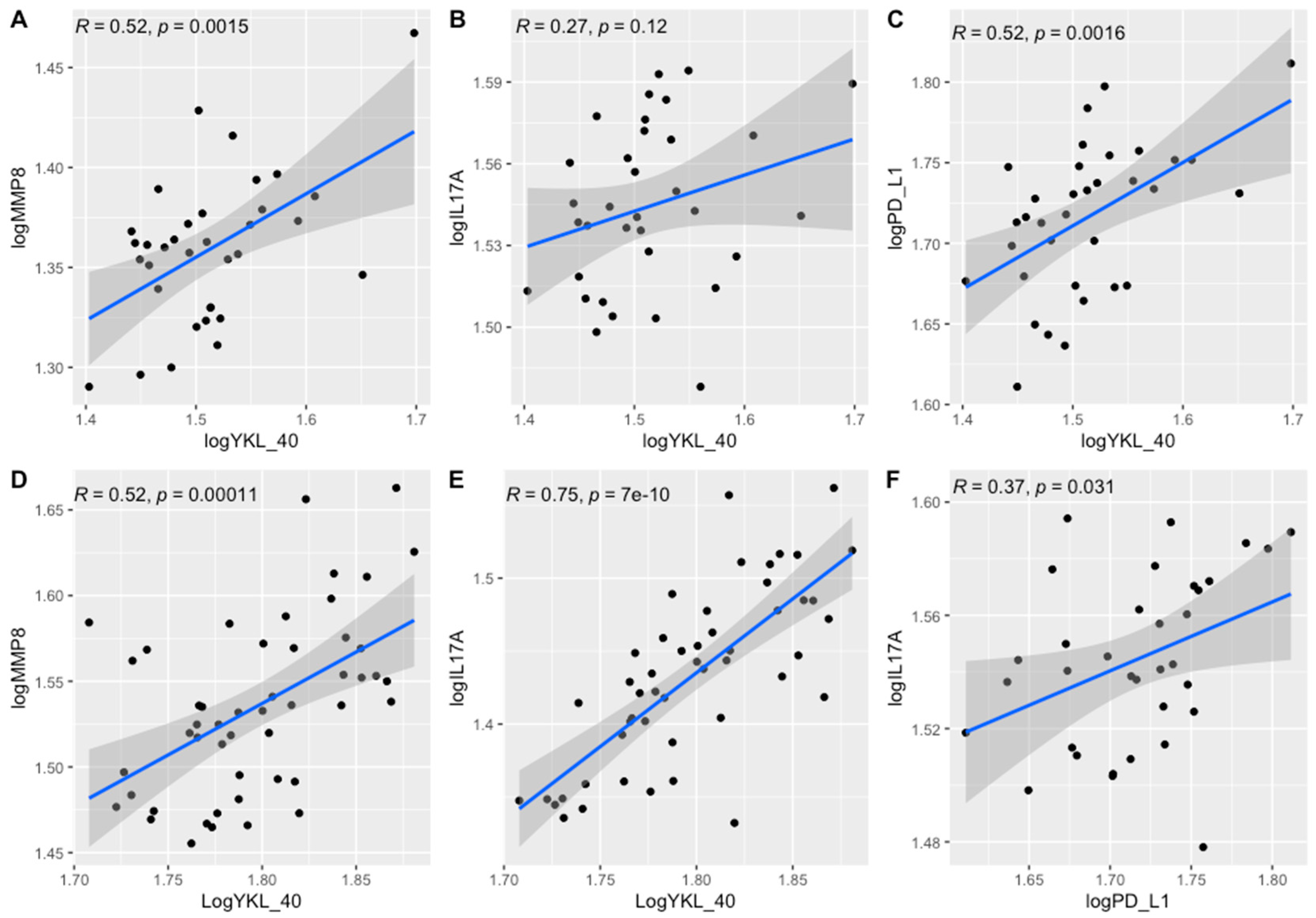
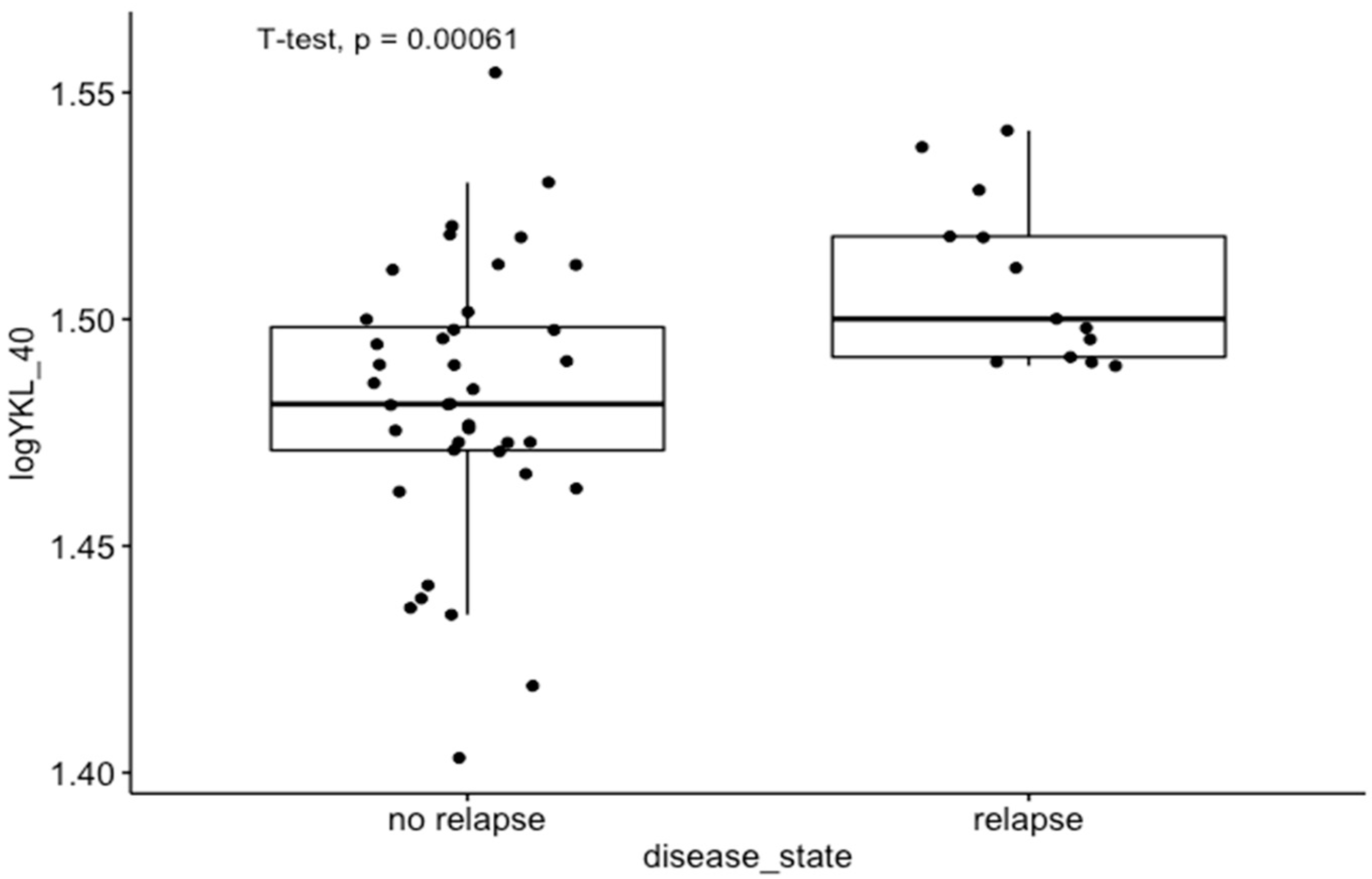
3.4. Results Obtained from the Tissue Homogenates
4. Discussion
4.1. Correlation with Clinical Data
4.2. Study Limitation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Oh, I.H.; Pyo, J.-S.; Son, B.K. Prognostic Impact of YKL-40 Immunohistochemical Expression in Patients with Colorectal Cancer. Curr. Oncol. 2021, 28, 3139–3149. [Google Scholar] [CrossRef]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-Protein-1 Function and Its Role in Diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Frühbeck, G. Increased Circulating and Visceral Adipose Tissue Expression Levels of YKL-40 in Obesity-Associated Type 2 Diabetes Are Related to Inflammation: Impact of Conventional Weight Loss and Gastric Bypass. J. Clin. Endocrinol. Metab. 2011, 96, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Hempen, M.; Kopp, H.-P.; Elhenicky, M.; Höbaus, C.; Brix, J.-M.; Koppensteiner, R.; Schernthaner, G.; Schernthaner, G.-H. YKL-40 Is Elevated in Morbidly Obese Patients and Declines after Weight Loss. Obes. Surg. 2009, 19, 1557–1563. [Google Scholar] [CrossRef]
- Fuksiewicz, M.; Kotowicz, B.; Rutkowski, A.; Achinger-Kawecka, J.; Wagrodzki, M.; Kowalska, M.M. The Assessment of Clinical Usage and Prognostic Value of YKL-40 Serum Levels in Patients With Rectal Cancer Without Distant Metastasis. Technol. Cancer Res. Treat. 2018, 17, 1533033818765209. [Google Scholar] [CrossRef] [Green Version]
- Razi, S.; Baradaran Noveiry, B.; Keshavarz-Fathi, M.; Rezaei, N. IL-17 and Colorectal Cancer: From Carcinogenesis to Treatment. Cytokine 2019, 116, 7–12. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, X.; Herjan, T.; Li, X. The Role of Interleukin-17 in Tumor Development and Progression. J. Exp. Med. 2019, 217, e20190297. [Google Scholar] [CrossRef]
- Awasthi, A.; Murugaiyan, G.; Kuchroo, V.K. Interplay between Effector Th17 and Regulatory T Cells. J. Clin. Immunol. 2008, 28, 660–670. [Google Scholar] [CrossRef]
- Malek-Hosseini, Z.; Taherinejad, M.; Malekzadeh, M.; Ghaderi, A.; Doroudchi, M. IL-17A Levels in the Sera of Patients with Gastric Cancer Show Limited Elevation. Asian Pac. J. Cancer Prev. 2015, 16, 7149–7153. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Dong, X. Colorectal Cancer-Derived Foxp3+IL-17+ T Cells Suppress Tumour-Specific CD8+ T Cells. Scand. J. Immunol. 2011, 74, 47–51. [Google Scholar] [CrossRef]
- Kortylewski, M.; Xin, H.; Kujawski, M.; Lee, H.; Liu, Y.; Harris, T.; Drake, C.; Pardoll, D.; Yu, H. Regulation of the IL-23 and IL-12 Balance by Stat3 Signaling in the Tumor Microenvironment. Cancer Cell 2009, 15, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Qu, Y.; Leng, Y.; Sun, W.; Ma, S.; Wei, J.; Hu, J.; Zhang, X. Human Colon Carcinogenesis Is Associated with Increased Interleukin-17-Driven Inflammatory Responses. Drug Des. Dev. Ther. 2015, 9, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-C.; Chen, C.-N.; Kuo, H.-C.; Shi, C.-S.; Hsieh, M.C.; Kuo, Y.-H.; Tung, S.-Y.; Lee, K.-F.; Huang, W.-S. Interleukin-17 Induces CC Chemokine Receptor 6 Expression and Cell Migration in Colorectal Cancer Cells. J. Cell. Physiol. 2015, 230, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Shen, J.; Cao, J.; Zhou, Y.; Shang, L.; Jin, S.; Cao, S.; Che, D.; Liu, F.; Yu, Y. Interleukin-17 Promotes Angiogenesis by Stimulating VEGF Production of Cancer Cells via the STAT3/GIV Signaling Pathway in Non-Small-Cell Lung Cancer. Sci. Rep. 2015, 5, 16053. [Google Scholar] [CrossRef] [Green Version]
- Parry, R.V.; Chemnitz, J.M.; Frauwirth, K.A.; Lanfranco, A.R.; Braunstein, I.; Kobayashi, S.V.; Linsley, P.S.; Thompson, C.B.; Riley, J.L. CTLA-4 and PD-1 Receptors Inhibit T-Cell Activation by Distinct Mechanisms. Mol. Cell. Biol. 2005, 25, 9543–9553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodig, N.; Ryan, T.; Allen, J.A.; Pang, H.; Grabie, N.; Chernova, T.; Greenfield, E.A.; Liang, S.C.; Sharpe, A.H.; Lichtman, A.H.; et al. Endothelial Expression of PD-L1 and PD-L2 down-Regulates CD8+ T Cell Activation and Cytolysis. Eur. J. Immunol. 2003, 33, 3117–3126. [Google Scholar] [CrossRef]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-Associated B7-H1 Promotes T-Cell Apoptosis: A Potential Mechanism of Immune Evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef]
- Schütz, F.; Stefanovic, S.; Mayer, L.; von Au, A.; Domschke, C.; Sohn, C. PD-1/PD-L1 Pathway in Breast Cancer. ORT 2017, 40, 294–297. [Google Scholar] [CrossRef]
- Tu, X.; Qin, B.; Zhang, Y.; Zhang, C.; Kahila, M.; Nowsheen, S.; Yin, P.; Yuan, J.; Pei, H.; Li, H.; et al. PD-L1 (B7-H1) Competes with the RNA Exosome to Regulate the DNA Damage Response and Can Be Targeted to Sensitize to Radiation or Chemotherapy. Mol. Cell 2019, 74, 1215–1226.e4. [Google Scholar] [CrossRef]
- Wu, X.; Gu, Z.; Chen, Y.; Chen, B.; Chen, W.; Weng, L.; Liu, X. Application of PD-1 Blockade in Cancer Immunotherapy. Comput. Struct. Biotechnol. J. 2019, 17, 661–674. [Google Scholar] [CrossRef]
- Wrobel, P.; Ahmed, S. Current Status of Immunotherapy in Metastatic Colorectal Cancer. Int. J. Color. Dis. 2019, 34, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Kiyotani, K.; Sakata, S.; Nagano, S.; Kumehara, S.; Baba, S.; Besse, B.; Yanagitani, N.; Friboulet, L.; Nishio, M.; et al. Secreted PD-L1 Variants Mediate Resistance to PD-L1 Blockade Therapy in Non–Small Cell Lung Cancer. J. Exp. Med. 2019, 216, 982–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 Contributes to Immunosuppression and Is Associated with Anti-PD-1 Response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- PD-L1 Degradation Is Regulated by Electrostatic Membrane Association of Its Cytoplasmic Domain|Nature Communications. Available online: https://www.nature.com/articles/s41467-021-25416-7 (accessed on 4 March 2023).
- Cha, J.-H.; Yang, W.-H.; Xia, W.; Wei, Y.; Chan, L.-C.; Lim, S.-O.; Li, C.-W.; Kim, T.; Chang, S.-S.; Lee, H.-H.; et al. Metformin Promotes Antitumor Immunity via Endoplasmic Reticulum-Associated Degradation of PD-L1. Mol. Cell 2018, 71, 606–620.e7. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, Z.; Hou, J.; Xiong, W.; Kim, H.; Chen, J.; Zheng, C.; Jiang, X.; Yoon, J.; Shen, J. Tumor Selective Metabolic Reprogramming as a Prospective PD-L1 Depression Strategy to Reactivate Immunotherapy. Adv. Mater. 2022, 34, 2206121. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, J.; Liu, Y.; Zheng, C.; Luo, W.; Chen, L.; Zhou, S.; Li, Z.; Shen, J. Cascade Two-Stage Tumor Re-Oxygenation and Immune Re-Sensitization Mediated by Self-Assembled Albumin-Sorafenib Nanoparticles for Enhanced Photodynamic Immunotherapy. Acta Pharm. Sin. B 2022, 12, 4204–4223. [Google Scholar] [CrossRef]
- Bao, R.; Wang, Y.; Lai, J.; Zhu, H.; Zhao, Y.; Li, S.; Li, N.; Huang, J.; Yang, Z.; Wang, F.; et al. Enhancing Anti-PD-1/PD-L1 Immune Checkpoint Inhibitory Cancer Therapy by CD276-Targeted Photodynamic Ablation of Tumor Cells and Tumor Vasculature. Mol. Pharm. 2019, 16, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Yang, C.; Luo, Y.; Liu, F.; Zheng, J.; Liu, W.; Ran, H.; Sun, Y.; Ren, J.; Wang, Z. Near-Infrared II Nanoadjuvant-Mediated Chemodynamic, Photodynamic, and Photothermal Therapy Combines Immunogenic Cell Death with PD-L1 Blockade to Enhance Antitumor Immunity. Small 2022, 18, e2107809. [Google Scholar] [CrossRef]
- CAMOIP: A Web Server for Comprehensive Analysis on Multi-Omics of Immunotherapy in Pan-Cancer|Briefings in Bioinformatics|Oxford Academic. Available online: https://academic.oup.com/bib/article-abstract/23/3/bbac129/6565619?redirectedFrom=fulltext (accessed on 2 February 2023).
- Dampier, C.H.; Devall, M.; Jennelle, L.T.; Díez-Obrero, V.; Plummer, S.J.; Moreno, V.; Casey, G. Oncogenic Features in Histologically Normal Mucosa: Novel Insights Into Field Effect From a Mega-Analysis of Colorectal Transcriptomes. Clin. Transl. Gastroenterol. 2020, 11, e00210. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gene Ontology Consortium. The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef] [PubMed]
- Khamas, A.; Ishikawa, T.; Shimokawa, K.; Mogushi, K.; Iida, S.; Ishiguro, M.; Mizushima, H.; Tanaka, H.; Uetake, H.; Sugihara, K. Screening for Epigenetically Masked Genes in Colorectal Cancer Using 5-Aza-2’-Deoxycytidine, Microarray and Gene Expression Profile. Cancer Genom. Proteom. 2012, 9, 67–75. [Google Scholar]
- Gröne, J.; Lenze, D.; Jurinovic, V.; Hummel, M.; Seidel, H.; Leder, G.; Beckmann, G.; Sommer, A.; Grützmann, R.; Pilarsky, C.; et al. Molecular Profiles and Clinical Outcome of Stage UICC II Colon Cancer Patients. Int. J. Color. Dis. 2011, 26, 847–858. [Google Scholar] [CrossRef]
- Alhopuro, P.; Sammalkorpi, H.; Niittymäki, I.; Biström, M.; Raitila, A.; Saharinen, J.; Nousiainen, K.; Lehtonen, H.J.; Heliövaara, E.; Puhakka, J.; et al. Candidate Driver Genes in Microsatellite-Unstable Colorectal Cancer. Int. J. Cancer 2012, 130, 1558–1566. [Google Scholar] [CrossRef]
- Ngernyuang, N.; Yan, W.; Schwartz, L.M.; Oh, D.; Liu, Y.; Chen, H.; Shao, R. A Heparin Binding Motif Rich in Arginine and Lysine Is the Functional Domain of YKL-40. Neoplasia 2017, 20, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Coffman, F.D. Chitinase 3-Like-1 (CHI3L1): A Putative Disease Marker at the Interface of Proteomics and Glycomics. Crit. Rev. Clin. Lab. Sci. 2008, 45, 531–562. [Google Scholar] [CrossRef]
- Fusetti, F.; Pijning, T.; Kalk, K.H.; Bos, E.; Dijkstra, B.W. Crystal Structure and Carbohydrate-Binding Properties of the Human Cartilage Glycoprotein-39. J. Biol. Chem. 2003, 278, 37753–37760. [Google Scholar] [CrossRef] [Green Version]
- Bigg, H.F.; Wait, R.; Rowan, A.D.; Cawston, T.E. The Mammalian Chitinase-like Lectin, YKL-40, Binds Specifically to Type I Collagen and Modulates the Rate of Type I Collagen Fibril Formation. J. Biol. Chem. 2006, 281, 21082–21095. [Google Scholar] [CrossRef] [Green Version]
- McQuade, K.J.; Beauvais, D.M.; Burbach, B.J.; Rapraeger, A.C. Syndecan-1 Regulates Alphavbeta5 Integrin Activity in B82L Fibroblasts. J. Cell Sci. 2006, 119, 2445–2456. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; He, C.H.; Yang, D.S.; Nguyen, T.; Cao, Y.; Kamle, S.; Lee, C.-M.; Gochuico, B.R.; Gahl, W.A.; Shea, B.S.; et al. Galectin-3 Interacts with the CHI3L1 Axis and Contributes to Hermansky-Pudlak Syndrome Lung Disease. J. Immunol. 2018, 200, 2140–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-M.; He, C.H.; Nour, A.M.; Zhou, Y.; Ma, B.; Park, J.W.; Kim, K.H.; Dela Cruz, C.; Sharma, L.; Nasr, M.L.; et al. IL-13Rα2 Uses TMEM219 in Chitinase 3-like-1-Induced Signalling and Effector Responses. Nat. Commun. 2016, 7, 12752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francescone, R.A.; Scully, S.; Faibish, M.; Taylor, S.L.; Oh, D.; Moral, L.; Yan, W.; Bentley, B.; Shao, R. Role of YKL-40 in the Angiogenesis, Radioresistance, and Progression of Glioblastoma. J. Biol. Chem. 2011, 286, 15332–15343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in Cancer: Mechanistic Findings and Clinical Applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, S.Y.; Law, H.K.-W. JNK in Tumor Microenvironment: Present Findings and Challenges in Clinical Translation. Cancers 2021, 13, 2196. [Google Scholar] [CrossRef]
- Kzhyshkowska, J.; Yin, S.; Liu, T.; Riabov, V.; Mitrofanova, I. Role of Chitinase-like Proteins in Cancer. Biol. Chem. 2016, 397, 231–247. [Google Scholar] [CrossRef]
- Freisinger, C.M.; Huttenlocher, A. Live Imaging and Gene Expression Analysis in Zebrafish Identifies a Link between Neutrophils and Epithelial to Mesenchymal Transition. PLoS ONE 2014, 9, e112183. [Google Scholar] [CrossRef]
- Hu, P.; Shen, M.; Zhang, P.; Zheng, C.; Pang, Z.; Zhu, L.; Du, J. Intratumoral Neutrophil Granulocytes Contribute to Epithelial-Mesenchymal Transition in Lung Adenocarcinoma Cells. Tumor Biol. 2015, 36, 7789–7796. [Google Scholar] [CrossRef]
- Cai, J.; Xia, L.; Li, J.; Ni, S.; Song, H.; Wu, X. Tumor-Associated Macrophages Derived TGF-Β-Induced Epithelial to Mesenchymal Transition in Colorectal Cancer Cells through Smad2,3-4/Snail Signaling Pathway. Cancer Res. Treat. 2019, 51, 252–266. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.-S.; Zhou, B.-H.; Li, C.-L.; Zhang, F.; Wang, X.-F.; Zhang, G.; Bu, X.-Z.; Cai, S.-H.; Du, J. Epithelial-Mesenchymal Transition (EMT) Induced by TNF-α Requires AKT/GSK-3β-Mediated Stabilization of Snail in Colorectal Cancer. PLoS ONE 2013, 8, e56664. [Google Scholar] [CrossRef]
- Yin, Y.; Yao, S.; Hu, Y.; Feng, Y.; Li, M.; Bian, Z.; Zhang, J.; Qin, Y.; Qi, X.; Zhou, L.; et al. The Immune-Microenvironment Confers Chemoresistance of Colorectal Cancer through Macrophage-Derived IL6. Clin. Cancer Res. 2017, 23, 7375–7387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, L.; Pappan, L.; Galliher-Beckley, A.; Shi, J. IL-1β Promotes Stemness and Invasiveness of Colon Cancer Cells through Zeb1 Activation. Mol. Cancer 2012, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshkian, Z.; Nobili, S.; Peyravian, N.; Shojaee, B.; Nazari, H.; Soleimani, H.; Asadzadeh-Aghdaei, H.; Ashrafian Bonab, M.; Nazemalhosseini-Mojarad, E.; Mini, E. Insights into the Role of Matrix Metalloproteinases in Precancerous Conditions and in Colorectal Cancer. Cancers 2021, 13, 6226. [Google Scholar] [CrossRef]
- Korpi, J.T.; Kervinen, V.; Mäklin, H.; Väänänen, A.; Lahtinen, M.; Läärä, E.; Ristimäki, A.; Thomas, G.; Ylipalosaari, M.; Aström, P.; et al. Collagenase-2 (Matrix Metalloproteinase-8) Plays a Protective Role in Tongue Cancer. Br. J. Cancer 2008, 98, 766–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böckelman, C.; Beilmann-Lehtonen, I.; Kaprio, T.; Koskensalo, S.; Tervahartiala, T.; Mustonen, H.; Stenman, U.-H.; Sorsa, T.; Haglund, C. Serum MMP-8 and TIMP-1 Predict Prognosis in Colorectal Cancer. BMC Cancer 2018, 18, 679. [Google Scholar] [CrossRef] [PubMed]
- Khashaba, S.A.; Attwa, E.; Said, N.; Ahmed, S.; Khattab, F. Serum YKL-40 and IL 17 in Psoriasis: Reliability as Prognostic Markers for Disease Severity and Responsiveness to Treatment. Dermatol. Ther. 2021, 34, e14606. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, R.; Wang, B.; Lian, J.; Yao, Y.; Sun, H.; Zhang, C.; Fang, L.; Guan, X.; Shi, J.; et al. Blocking IL-17A Enhances Tumor Response to Anti-PD-1 Immunotherapy in Microsatellite Stable Colorectal Cancer. J. Immunother. Cancer 2021, 9, e001895. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Akosman, B.; Kamle, S.; Lee, C.-M.; He, C.H.; Koo, J.S.; Lee, C.G.; Elias, J.A. CHI3L1 Regulates PD-L1 and Anti–CHI3L1–PD-1 Antibody Elicits Synergistic Antitumor Responses. J. Clin. Investig. 2021, 131, e137750. [Google Scholar] [CrossRef]
- Yang, X.; Fang, D.; Li, M.; Chen, J.; Cheng, Y.; Luo, J. Knockdown of Chitinase 3-Like-1 Inhibits Cell Proliferation, Promotes Apoptosis, and Enhances Effect of Anti-Programmed Death Ligand 1 (PD-L1) in Diffuse Large B Cell Lymphoma Cells. Med. Sci. Monit. 2021, 27, e929431-1–e929431-9. [Google Scholar] [CrossRef]
- Edin, S.; Wikberg, M.L.; Dahlin, A.M.; Rutegård, J.; Öberg, Å.; Oldenborg, P.-A.; Palmqvist, R. The Distribution of Macrophages with a M1 or M2 Phenotype in Relation to Prognosis and the Molecular Characteristics of Colorectal Cancer. PLoS ONE 2012, 7, e47045. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Barreiro, L.; Zhang, L.; Abdel-Rahman, S.A.; Naik, S.P.; Gabr, M. Gut Microbial-Derived Metabolites as Immune Modulators of T Helper 17 and Regulatory T Cells. Int. J. Mol. Sci. 2023, 24, 1806. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, Y.; Ma, X.; Wang, Y.; Liu, J.; Yang, C.; Wang, Y.; Bao, C.; Song, X.; Feng, Y.; et al. A Study on the Correlation between M2 Macrophages and Regulatory T Cells in the Progression of Colorectal Cancer. Int. J. Biol. Markers 2022, 37, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qi, S.; Zhu, Y.-B.; Ding, L. Prognostic Value of YKL-40 in Colorectal Carcinoma Patients: A Meta-Analysis. World J. Clin. Cases 2022, 10, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Peltonen, R.; Hagström, J.; Tervahartiala, T.; Sorsa, T.; Haglund, C.; Isoniemi, H. High Expression of MMP-9 in Primary Tumors and High Preoperative MPO in Serum Predict Improved Prognosis in Colorectal Cancer with Operable Liver Metastases. OCL 2021, 99, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Reijonen, P.; Peltonen, R.; Tervahartiala, T.; Sorsa, T.; Isoniemi, H. Serum Matrix Metalloproteinase-8 and Myeloperoxidase Predict Survival after Resection of Colorectal Liver Metastases. OCL 2021, 99, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Sirniö, P.; Tuomisto, A.; Tervahartiala, T.; Sorsa, T.; Klintrup, K.; Karhu, T.; Herzig, K.-H.; Mäkelä, J.; Karttunen, T.J.; Salo, T.; et al. High-Serum MMP-8 Levels Are Associated with Decreased Survival and Systemic Inflammation in Colorectal Cancer. Br. J. Cancer 2018, 119, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Sharp, S.P.; Avram, D.; Stain, S.C.; Lee, E.C. Local and Systemic Th17 Immune Response Associated with Advanced Stage Colon Cancer. J. Surg. Res. 2017, 208, 180–186. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Su, H.; Zhong, W.; Yuan, Y.; Yu, Z.; Fang, Y.; Zhou, H.; Li, C.; Huang, K. Interleukin-17 Is a Favorable Prognostic Marker for Colorectal Cancer. Clin. Transl. Oncol. 2015, 17, 50–56. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, Y.; Dong, W.; Lin, M.; He, J.; Zhang, X.; Tian, T.; Yang, Y.; Chen, K.; Lei, Q.-Y.; et al. D-Mannose Facilitates Immunotherapy and Radiotherapy of Triple-Negative Breast Cancer via Degradation of PD-L1. Proc. Natl. Acad. Sci. USA 2022, 119, e2114851119. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Song, W.; Jiang, X.; Deng, Z.; Xiong, W.; Shen, J. Metabolic Reprogramming Mediated PD-L1 Depression and Hypoxia Reversion to Reactivate Tumor Therapy. J. Control. Release 2022, 352, 793–812. [Google Scholar] [CrossRef]
- Mostafa, M.M.; Amin, M.M.; Zakaria, M.Y.; Hussein, M.A.; Shamaa, M.M.; Abd El-Halim, S.M. Chitosan Surface-Modified PLGA Nanoparticles Loaded with Cranberry Powder Extract as a Potential Oral Delivery Platform for Targeting Colon Cancer Cells. Pharmaceutics 2023, 15, 606. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Yu, J.E.; Kim, K.C.; Lee, D.H.; Son, D.J.; Lee, H.P.; Jung, J.-K.; Kim, N.D.; Ham, Y.W.; Yun, J.; et al. A Small Molecule Targeting CHI3L1 Inhibits Lung Metastasis by Blocking IL-13Rα2-Mediated JNK-AP-1 Signals. Mol. Oncol. 2022, 16, 508–526. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female | Male | ||
|---|---|---|---|
| 20 | 28 | 48 (100%) | |
| Age | 63.06 ± 11.27 | 63.81 ± 8.61 | 63,47 ± 9.75 |
| BMI | 24.99 ± 5.06 | 26.75 ± 4.50 | 25.96 ± 4.78 |
| Obesity | |||
| Yes | 3 (15%) | 5 (17.86%) | 8 (16.67%) |
| No | 17 (85%) | 23 (82.14%) | 40 (83.33%) |
| Diabetes | |||
| Yes | 4 (20%) | 10 (35.71%) | 14 (29.17%) |
| No | 16 (8%) | 18 (64.29%) | 34 (70.83%) |
| Nicotinism | |||
| Yes | 8 (40%) | 9 (32.14%) | 17 (35.42%) |
| No | 12 (60%) | 19 (67.86%) | 31 (64.58%) |
| T parameter | |||
| T1 | 0 (0%) | 0 (0%) | 0 (0%) |
| T2 | 7 (35%) | 5 (17.86%) | 12 (25%) |
| T3 | 10 (50%) | 14 (50%) | 24 (50%) |
| T4 | 3 (15%) | 9 (32.14%) | 12 (25%) |
| N parameter | |||
| N0 | 9 (45%) | 12 (42.86%) | 21 (43.75%) |
| N1 | 8 (40%) | 9 (32.14%) | 17 (35.42%) |
| N2 | 3 (15%) | 7 (25%) | 10 (20.83%) |
| M parameter | |||
| M0 | 18 (90%) | 19 (67.86%) | 37 (77.08%) |
| M1 | 2 (10%) | 9 (32.14%) | 11 (22.92%) |
| TNM stage | |||
| I | 6 (30%) | 4 (14.29%) | 10 (20.83%) |
| II | 3 (15%) | 7 (25.00%) | 10 (20.83%) |
| III | 9 (45%) | 8 (28.57%) | 17 (35.42%) |
| IV | 2 (10%) | 9 (32.14%) | 11 (22.92%) |
| Grading | |||
| G1 | 1 (5%) | 0 (0%) | 1 (2.08%) |
| G2 | 19 (95%) | 28 (100%) | 47 (97.92%) |
| G3 | 0(0%) | 0 (0%) | 0 (0%) |
| Tumor | Margin | p | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
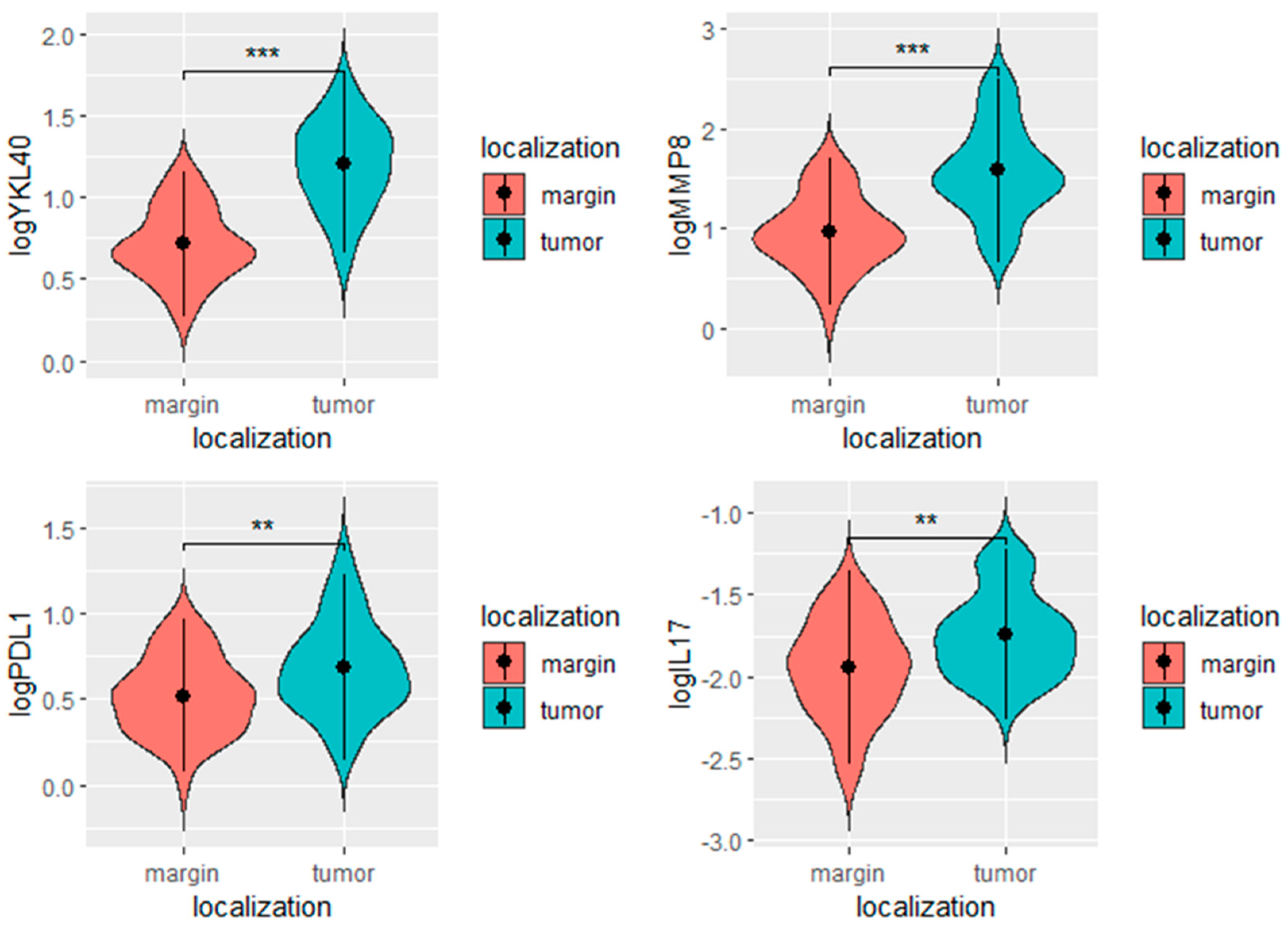
| log YKL-40 | 1.20 | 0.27 | 0.71 | 0.22 | <0.0001 |
| log PD-L1 | 0.69 | 0.27 | 0.52 | 0.22 | 0.001 |
| log MMP-8 | 1.58 | 0.46 | 0.96 | 0.37 | <0.0001 |
| log IL-17 | −1.74 | 0.26 | −1.95 | 0.29 | <0.0001 |
| Pair of Variables | R | p |
|---|---|---|
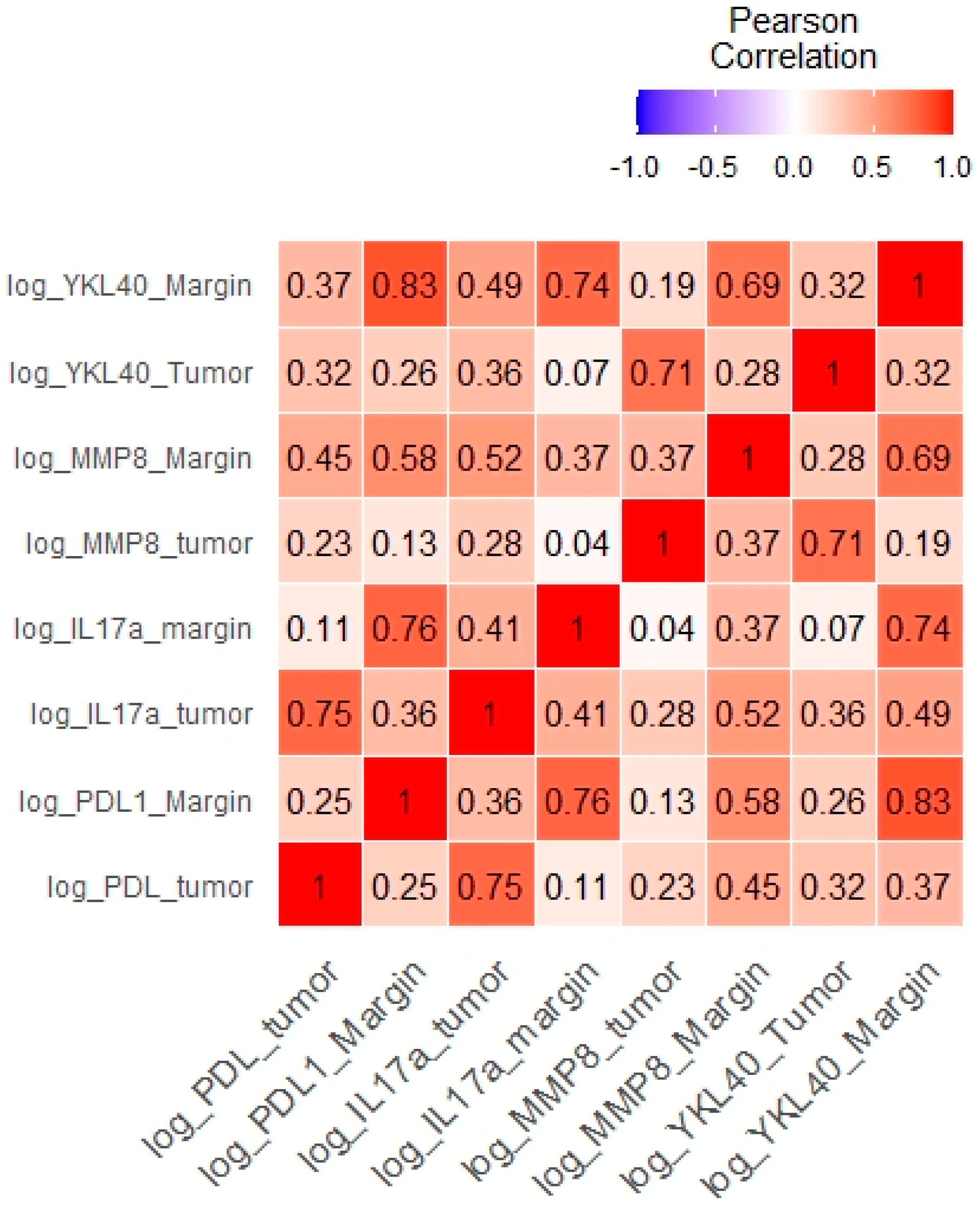
| Tumor log YKL-40 and tumor log PD-L1 | 0.32 | 0.044 |
| Tumor log YKL-40 and tumor log MMP-8 | 0.71 | <0.0001 |
| Tumor log YKL-40 and tumor log IL-17 | 0.36 | 0.026 |
| Tumor log YKL-40 and margin log YKL-40 | 0.32 | 0.032 |
| Margin log YKL-40 and margin log PD-L1 | 0.37 | 0.022 |
| Margin log YKL-40 and margin log IL-17 | 0.49 | <0.0001 |
| Margin log YKL-40 and margin log MMP-8 | 0.70 | <0.0001 |
| Margin log YKL-40 and tumor log IL-17 | 0.49 | 0.022 |
| Margin log YKL-40 and tumor PD-L1 | 0.37 | 0.022 |
| Tumor log PD-L1 and tumor IL-17 | 0.75 | <0.001 |
| Mean | SD | Mean | SD | p | |
|---|---|---|---|---|---|
| log YKL-40_tumor | Diabetes | no Diabetes | |||
| 1.22 | 0.31 | 1.2 | 0.24 | 0.79 | |
| obesity | no obesity | ||||
| 1.22 | 0.43 | 1.2 | 0.25 | 0.86 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochman, B.; Mielcarska, S.; Kula, A.; Dawidowicz, M.; Robotycka, J.; Piecuch, J.; Szrot, M.; Dzięgielewska-Gęsiak, S.; Muc-Wierzgoń, M.; Waniczek, D.; et al. Do Elevated YKL-40 Levels Drive the Immunosuppressive Tumor Microenvironment in Colorectal Cancer? Assessment of the Association of the Expression of YKL-40, MMP-8, IL17A, and PD-L1 with Coexisting Type 2 Diabetes, Obesity, and Active Smoking. Curr. Issues Mol. Biol. 2023, 45, 2781-2797. https://doi.org/10.3390/cimb45040182
Ochman B, Mielcarska S, Kula A, Dawidowicz M, Robotycka J, Piecuch J, Szrot M, Dzięgielewska-Gęsiak S, Muc-Wierzgoń M, Waniczek D, et al. Do Elevated YKL-40 Levels Drive the Immunosuppressive Tumor Microenvironment in Colorectal Cancer? Assessment of the Association of the Expression of YKL-40, MMP-8, IL17A, and PD-L1 with Coexisting Type 2 Diabetes, Obesity, and Active Smoking. Current Issues in Molecular Biology. 2023; 45(4):2781-2797. https://doi.org/10.3390/cimb45040182
Chicago/Turabian StyleOchman, Błażej, Sylwia Mielcarska, Agnieszka Kula, Miriam Dawidowicz, Julia Robotycka, Jerzy Piecuch, Monika Szrot, Sylwia Dzięgielewska-Gęsiak, Małgorzata Muc-Wierzgoń, Dariusz Waniczek, and et al. 2023. "Do Elevated YKL-40 Levels Drive the Immunosuppressive Tumor Microenvironment in Colorectal Cancer? Assessment of the Association of the Expression of YKL-40, MMP-8, IL17A, and PD-L1 with Coexisting Type 2 Diabetes, Obesity, and Active Smoking" Current Issues in Molecular Biology 45, no. 4: 2781-2797. https://doi.org/10.3390/cimb45040182
APA StyleOchman, B., Mielcarska, S., Kula, A., Dawidowicz, M., Robotycka, J., Piecuch, J., Szrot, M., Dzięgielewska-Gęsiak, S., Muc-Wierzgoń, M., Waniczek, D., & Świętochowska, E. (2023). Do Elevated YKL-40 Levels Drive the Immunosuppressive Tumor Microenvironment in Colorectal Cancer? Assessment of the Association of the Expression of YKL-40, MMP-8, IL17A, and PD-L1 with Coexisting Type 2 Diabetes, Obesity, and Active Smoking. Current Issues in Molecular Biology, 45(4), 2781-2797. https://doi.org/10.3390/cimb45040182