
Overexpression of OLIG2 and MYT1L Transcription Factors Enhance the Differentiation Potential of Human Mesenchymal Stem Cells into Oligodendrocytes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
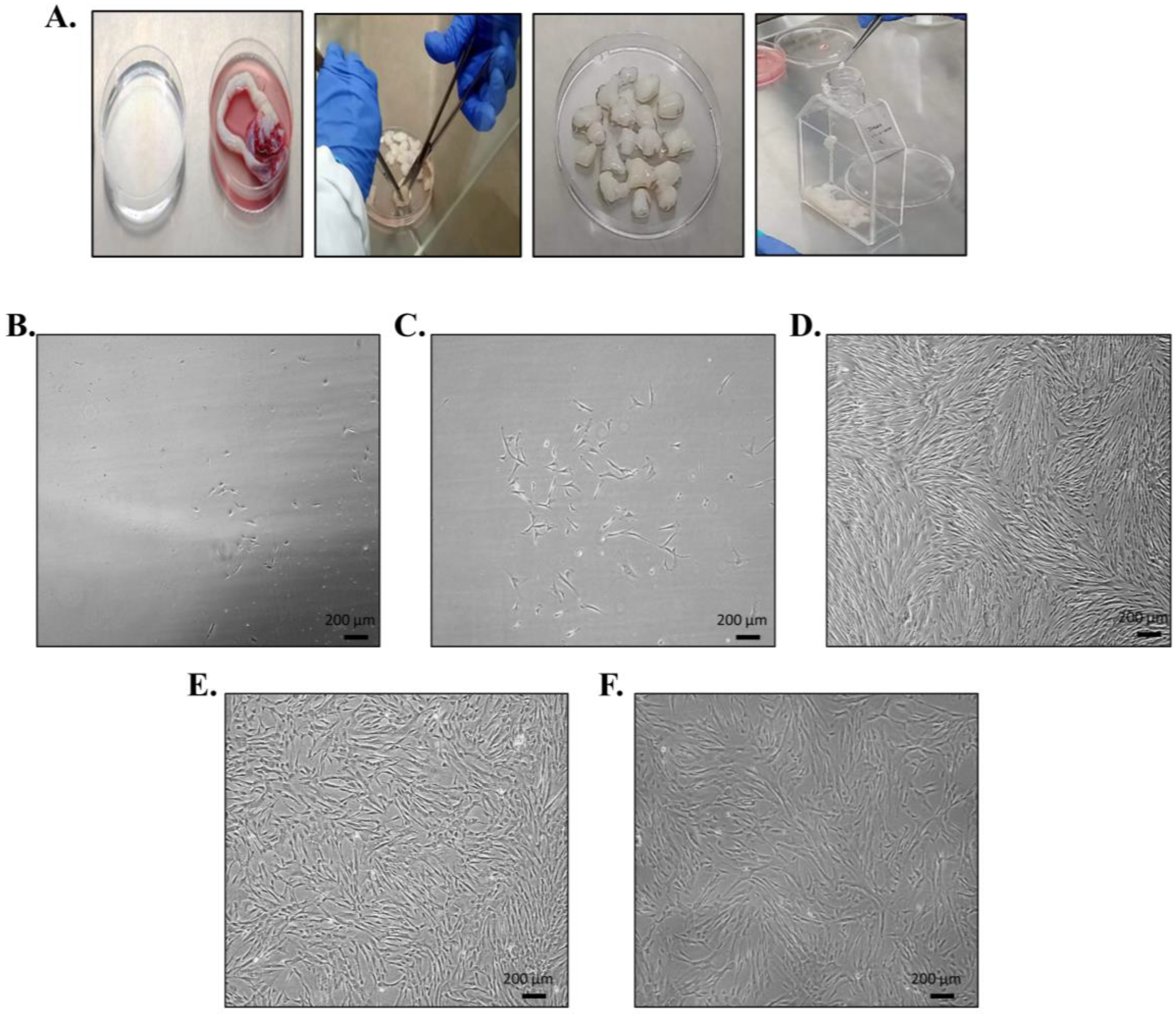
2.1. Ethical Consent and Umbilical Cord Sample Collection
2.2. Isolation and Propagation of hUC-MSCs
2.3. Characterization of hUC-MSCs
2.3.1. Morphological Features of hUC-MSCs
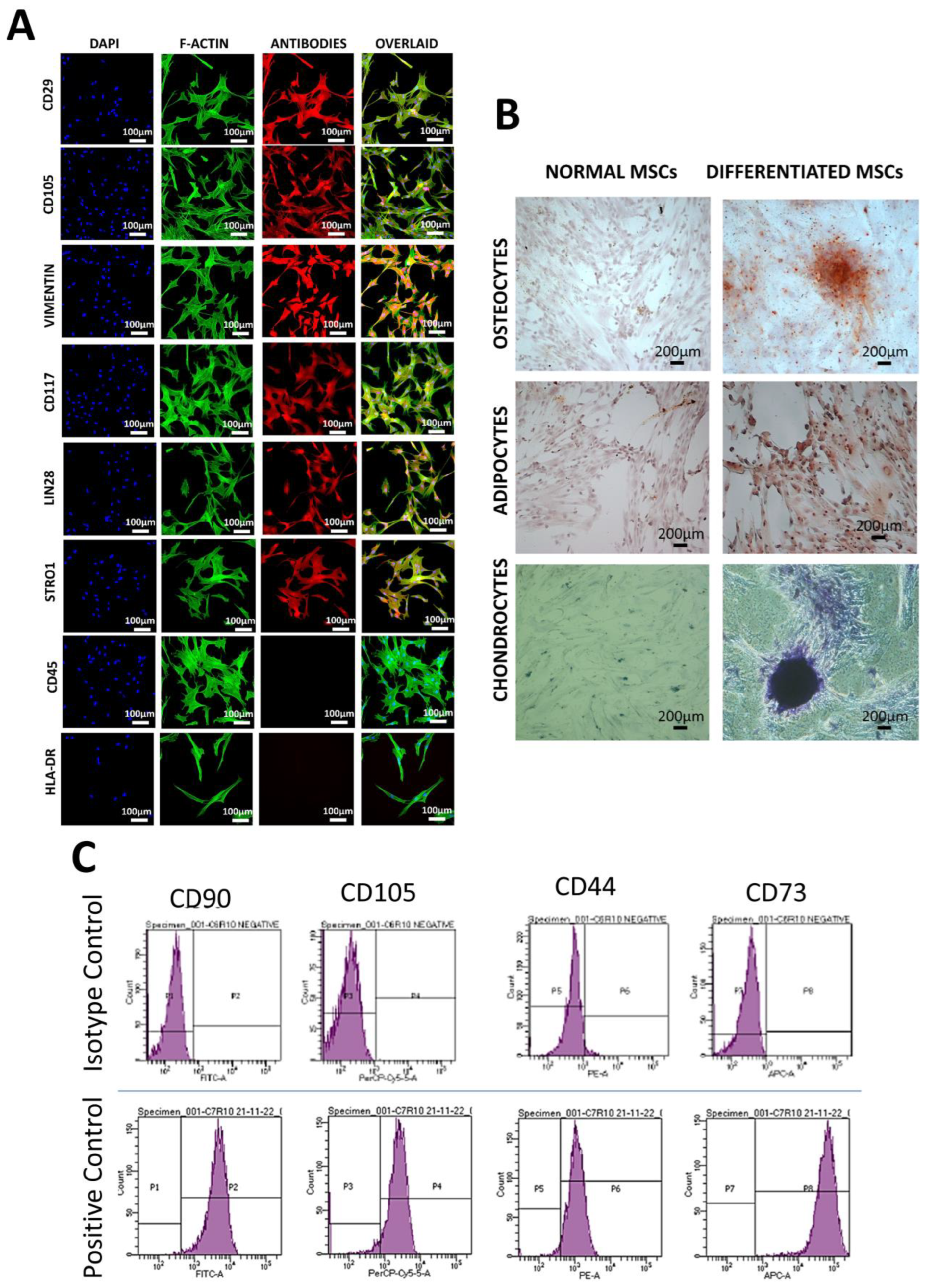
2.3.2. Immunocytochemistry
2.3.3. Immunophenotyping
2.3.4. Trilineage Differentiation
2.4. Isolation of Plasmid Vectors
2.5. Experimental Groups
2.6. Transfection of hUC-MSCs
2.7. Morphological Examination of Transfected hUC-MSCs
2.8. Gene Expression Analysis
2.9. Protein Expression Analysis of Transfected MSCs
2.10. Statistical Analysis
3. Results
3.1. Umbilical Cord Processing; Isolation, Propagation, and Morphological Features of hUC-MSCs
3.2. Characterization of hUC-MSCs
3.3. Transfection, Differentiation, and Morphological Assessment of hUC-MSCs
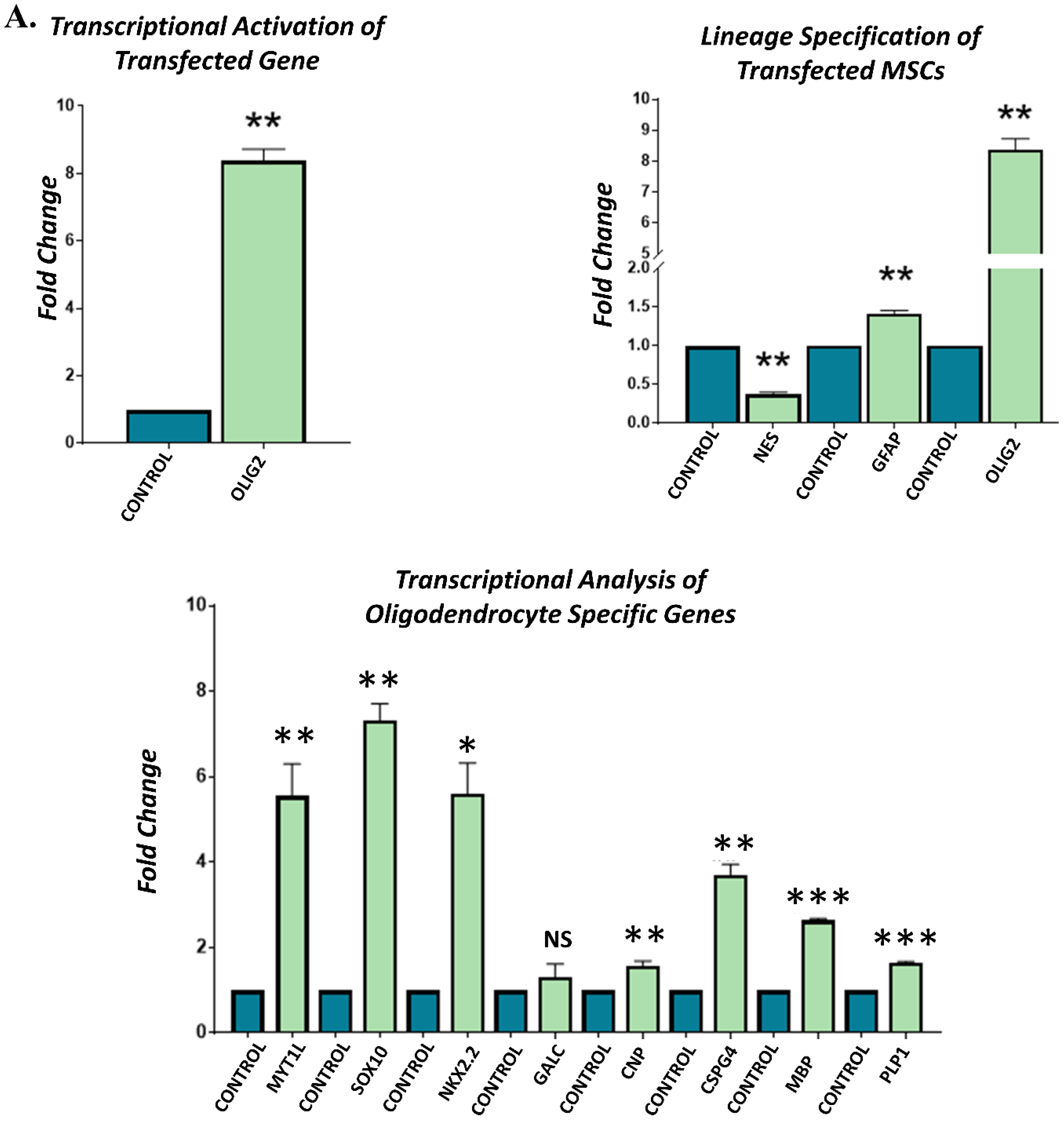
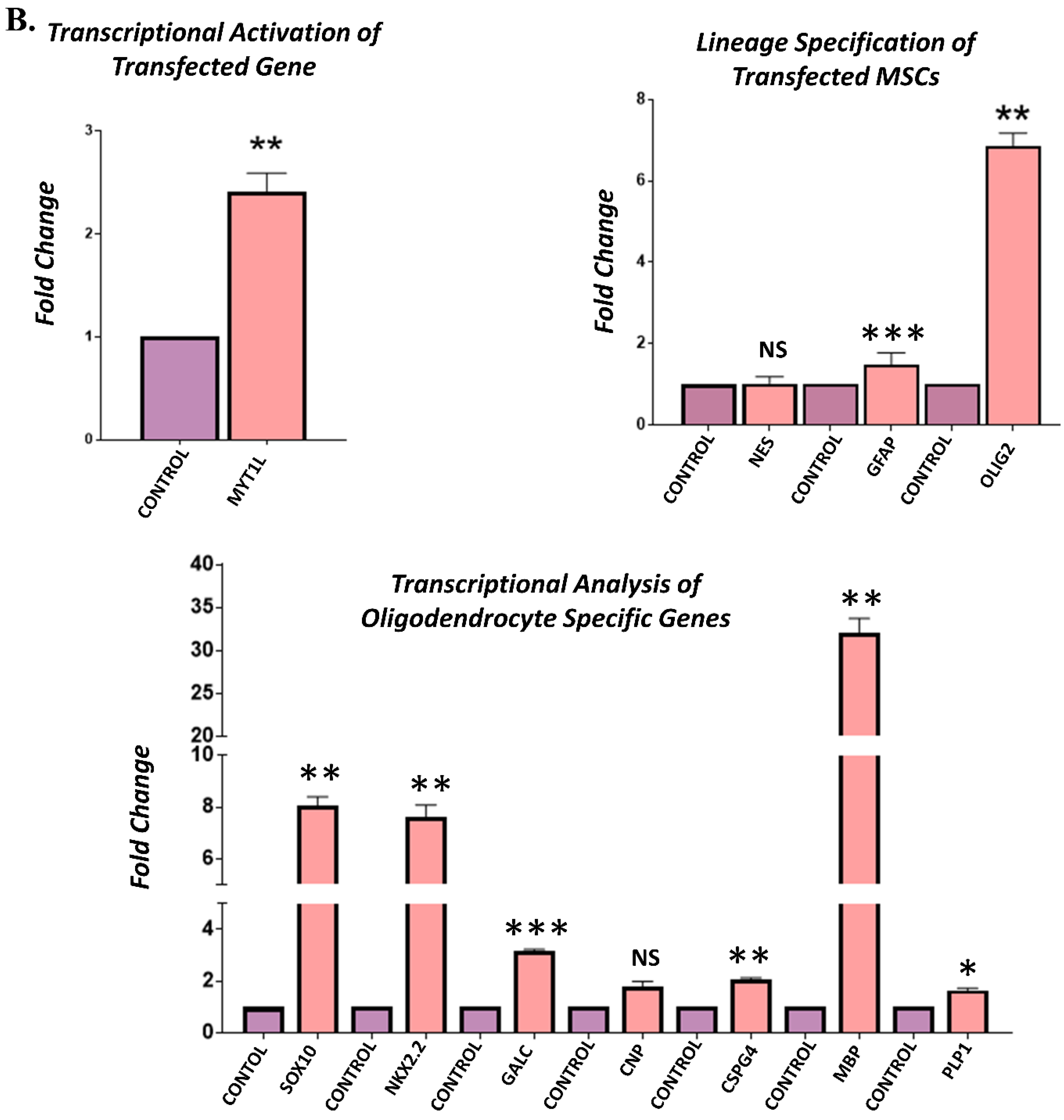
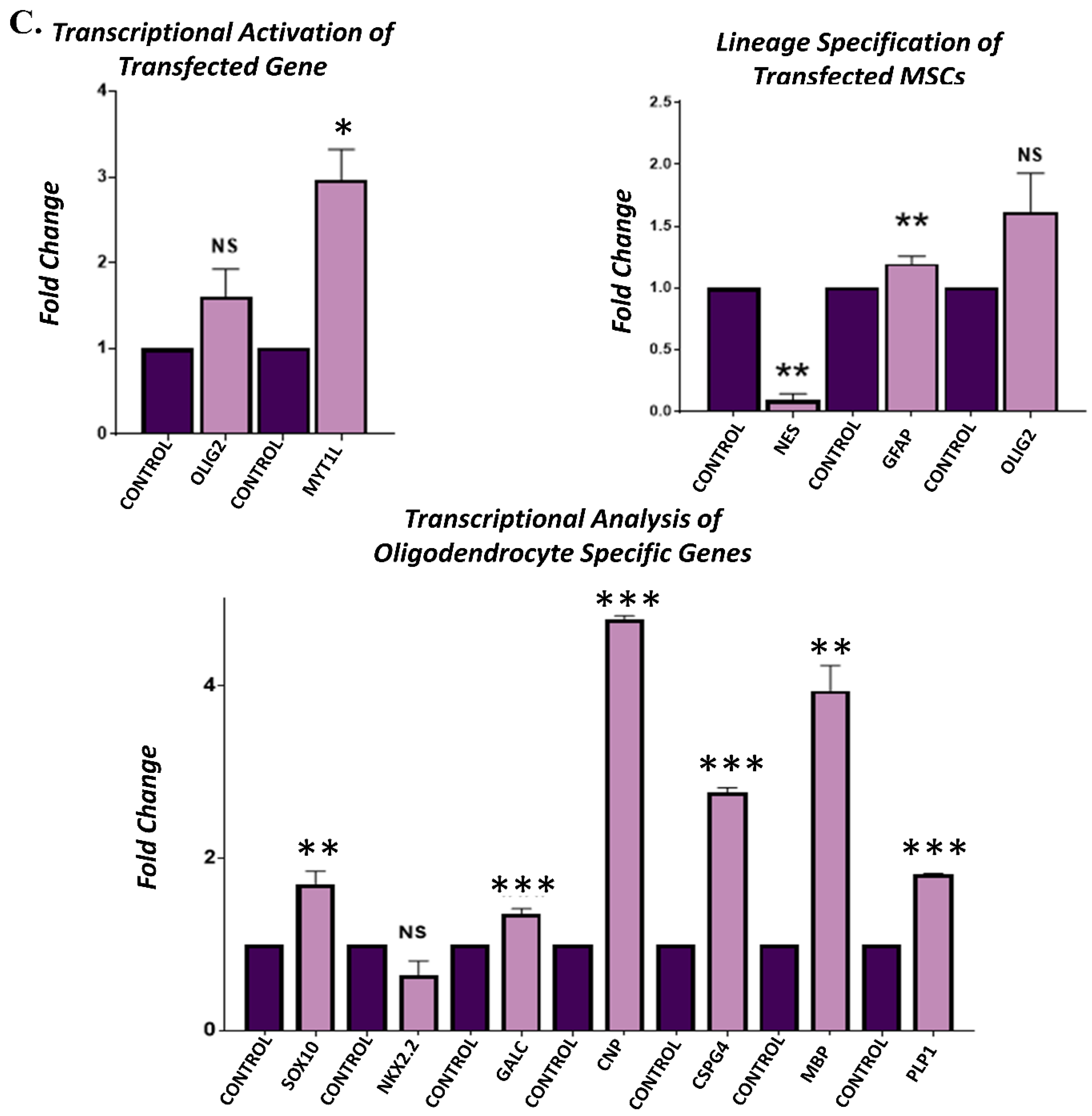
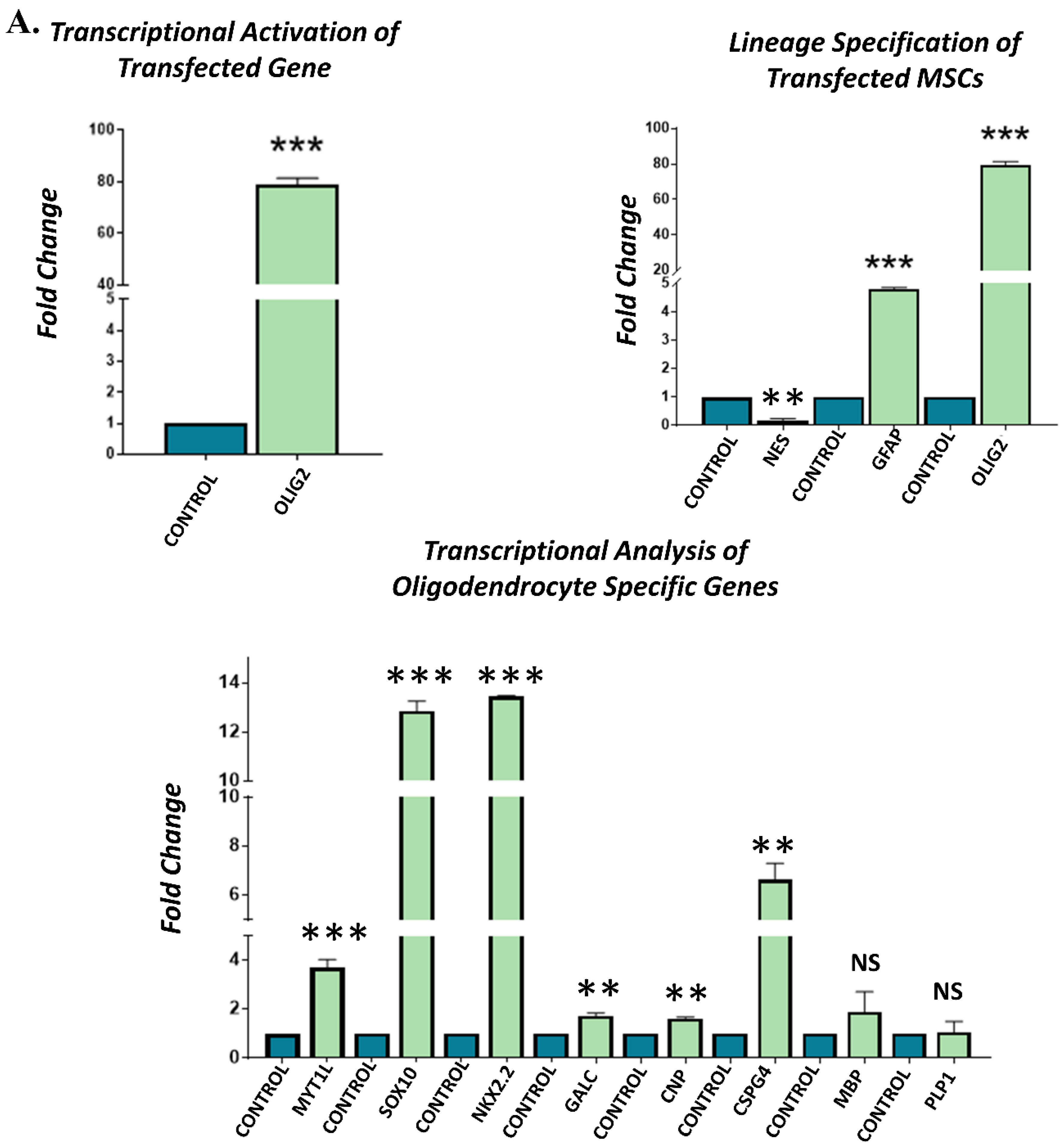
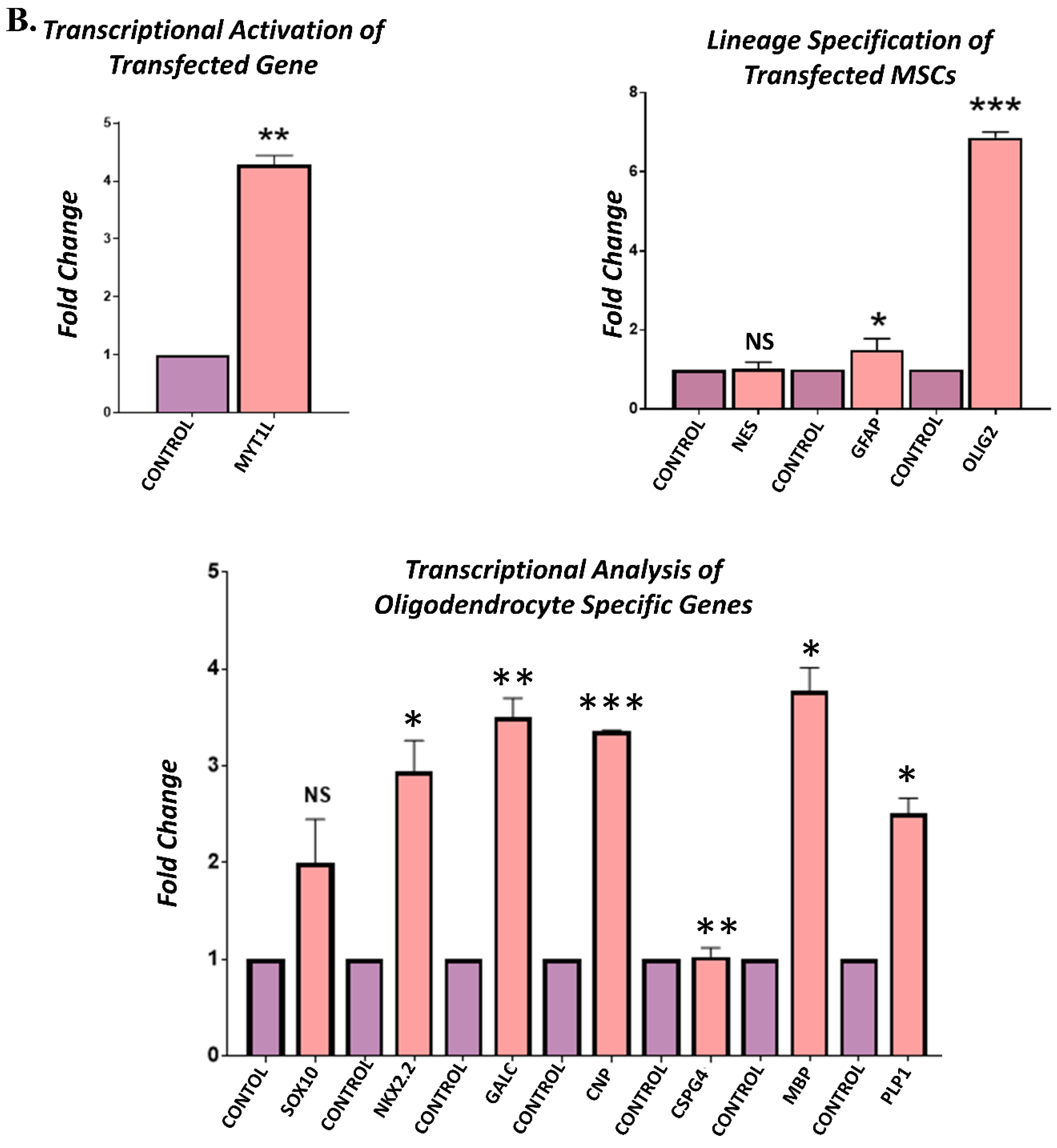
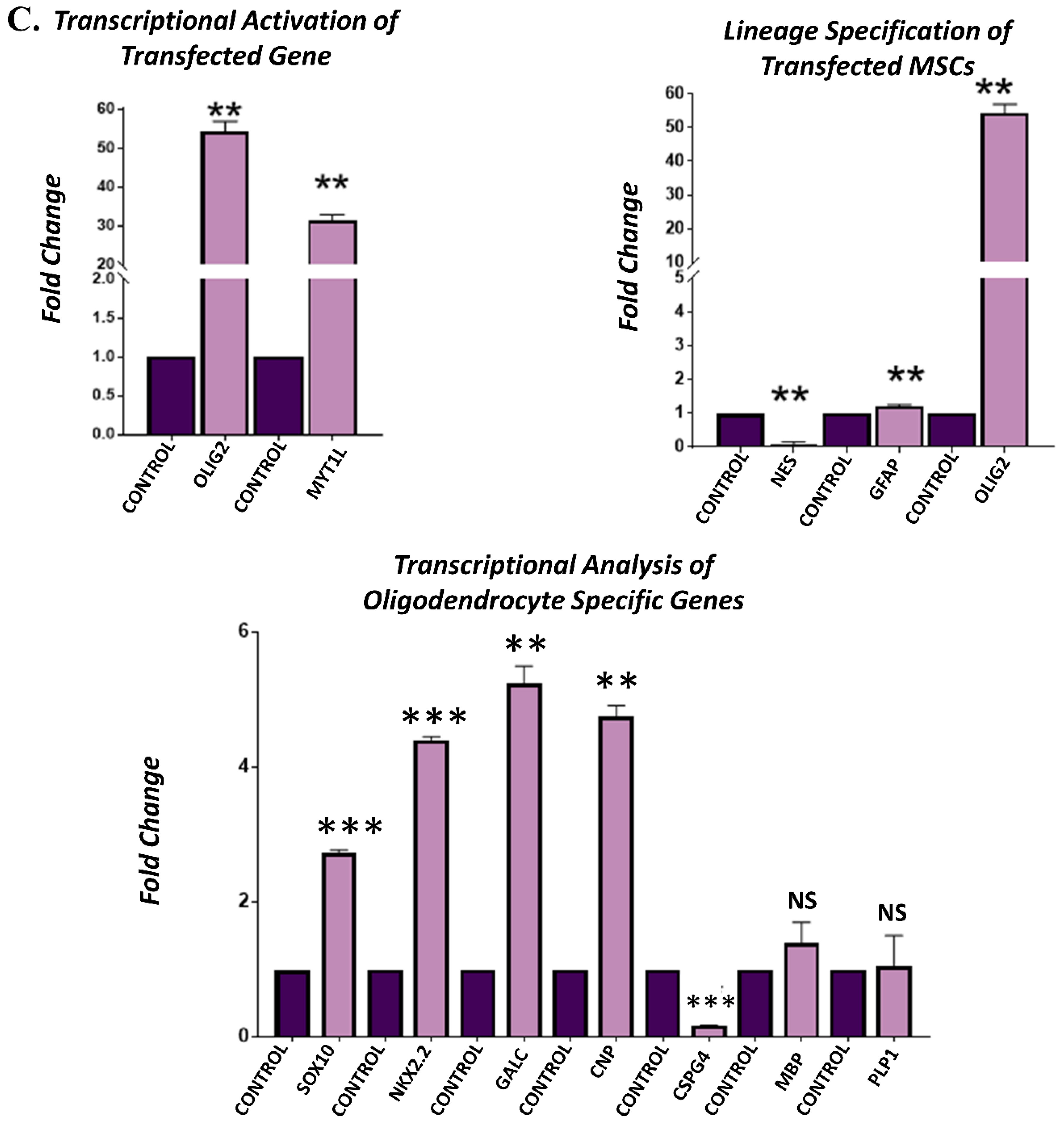
3.4. Gene Expression Analysis of Transfected hUC-MSCs Incubated in Normal Medium
3.5. Gene Expression Analysis of Transfected hUC-MSCs Incubated in Oligo Induction Medium
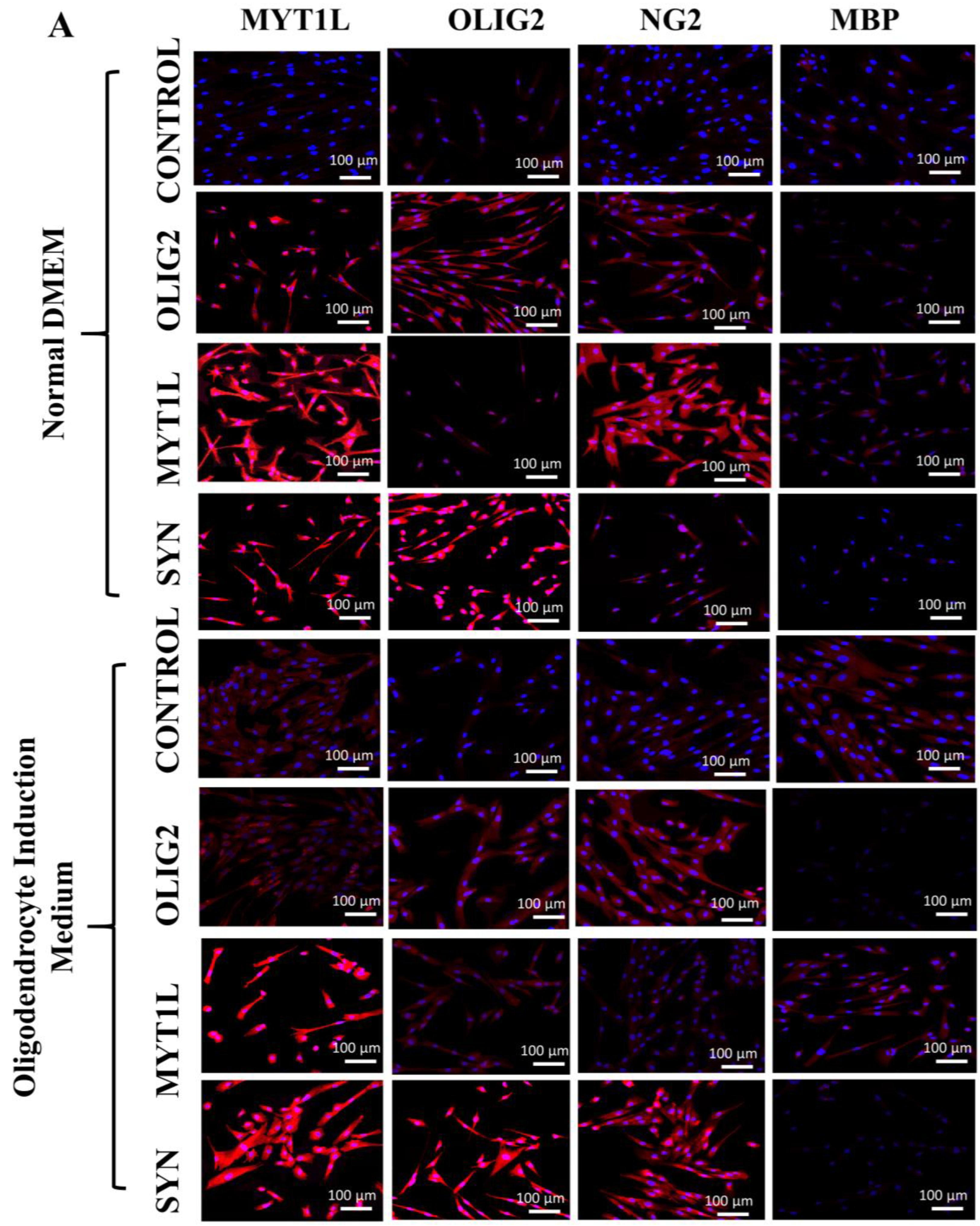
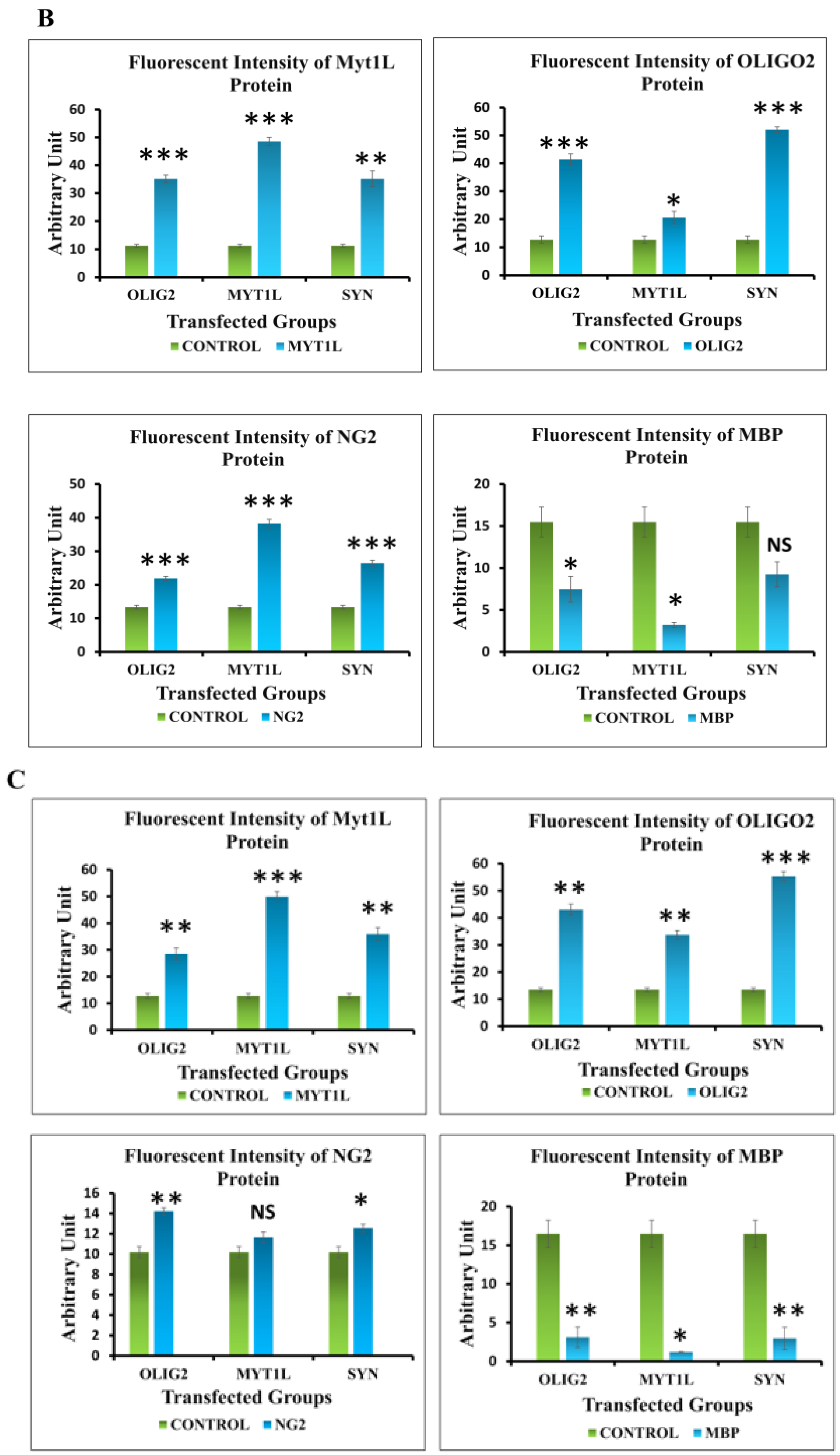
3.6. Protein Expression Analysis of Transfected hUC-MSCs in Normal and Oligo Induction Media after 3 Days of Incubation
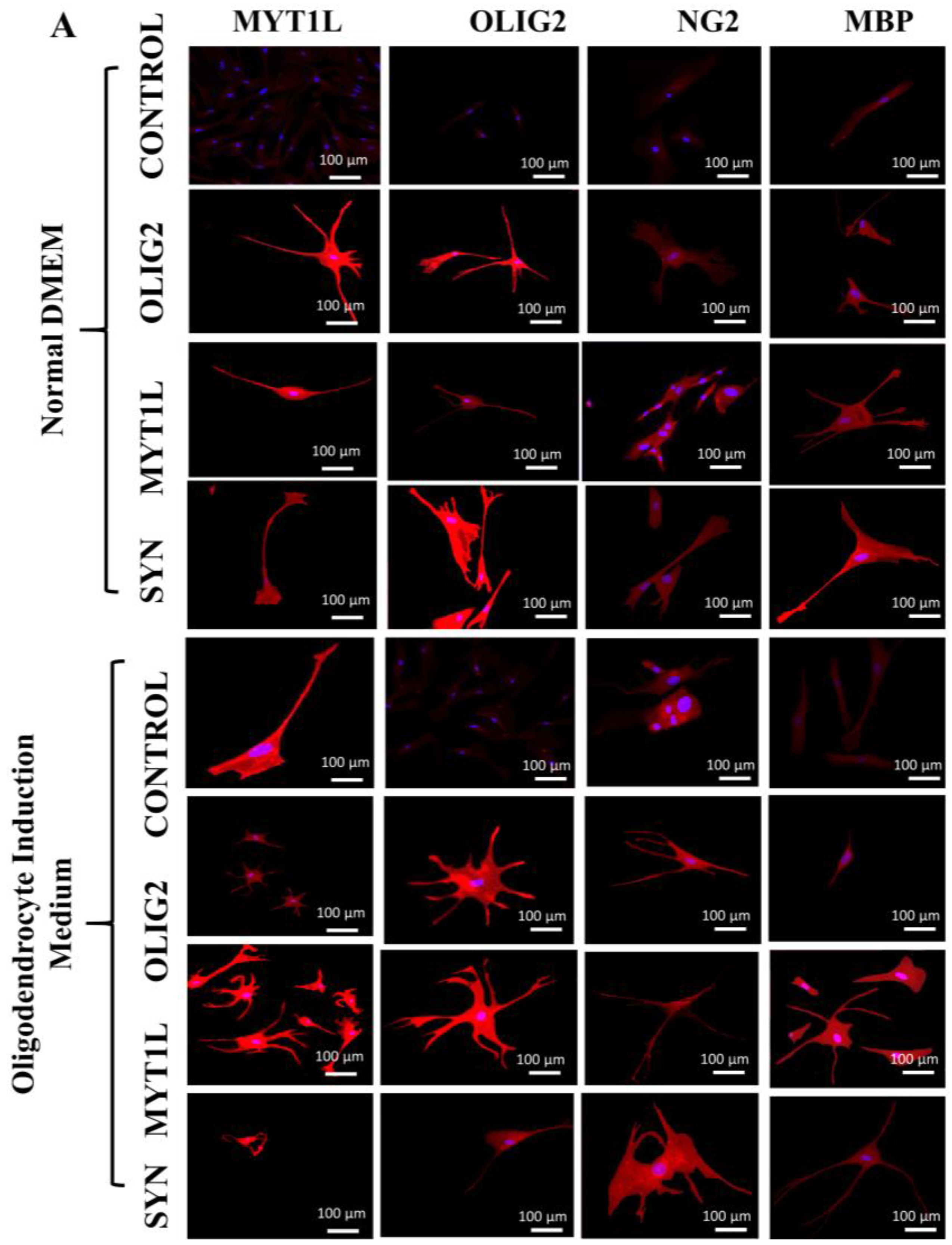
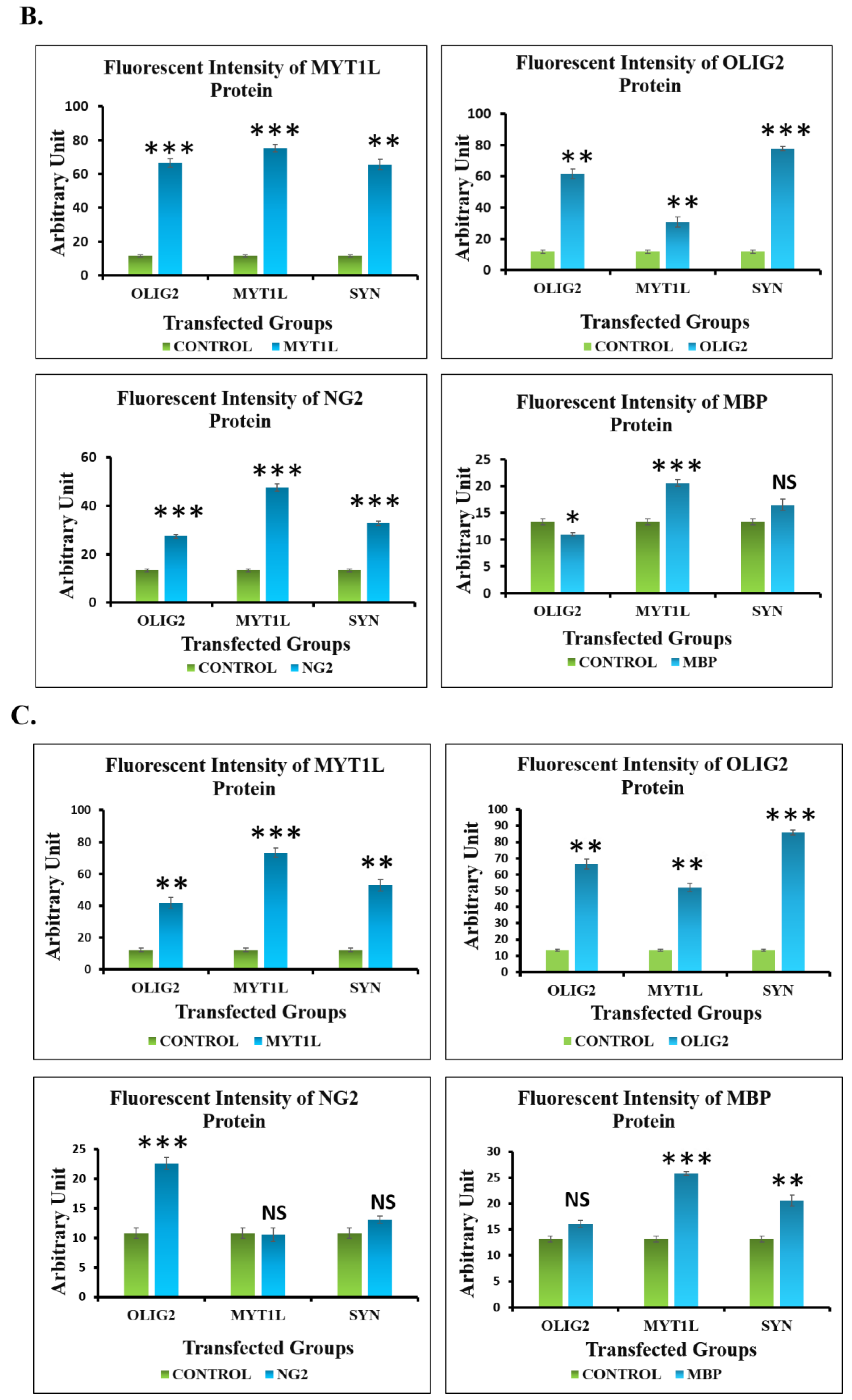
3.7. Protein Expression Analysis of Transfected hUC-MSCs in Normal and Oligo Induction Media after 7 Days of Incubation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cruz-Martinez, P.; González-Granero, S.; Molina-Navarro, M.M.; Pacheco-Torres, J.; García-Verdugo, J.M.; Geijo-Barrientos, E.; Jones, J.; Martinez, S. Intraventricular Injections of Mesenchymal Stem Cells Activate Endogenous Functional Remyelination in a Chronic Demyelinating Murine Model. Cell Death Dis. 2016, 7, e2223. [Google Scholar] [CrossRef] [PubMed]
- Dulamea, A.O. The Contribution of Oligodendrocytes and Oligodendrocyte Progenitor Cells to Central Nervous System Repair in Multiple Sclerosis: Perspectives for Remyelination Therapeutic Strategies. Neural Regen. Res. 2017, 12, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.A.; Denkova, M.; Ramanathan, S.; Dale, R.C.; Brilot, F. Pathogenesis of Autoimmune Demyelination: From Multiple Sclerosis to Neuromyelitis Optica Spectrum Disorders and Myelin Oligodendrocyte Glycoprotein Antibody-Associated Disease. Clin. Transl. Immunol. 2021, 10, e1316. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tang, H.; Zhu, J.; Zhang, J.H. Transplanting Mesenchymal Stem Cells for Treatment of Ischemic Stroke. Cell Transplant. 2018, 27, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, A.; Salim, A.; Simjee, S.U.; Khan, I.; Adli, D.S.H. Alpha Terpineol Directs Bone Marrow Mesenchymal Stem Cells toward Neuronal Lineage through Regulation of Wnt Signaling Pathway. Cell Biochem. Funct. 2023, 41, 223–233. [Google Scholar] [CrossRef]
- Islam, M.A.; Kundu, S.; Hassan, R. Gene Therapy Approaches in an Autoimmune Demyelinating Disease: Multiple Sclerosis. Curr. Gene Ther. 2020, 19, 376–385. [Google Scholar] [CrossRef]
- Stangel, M.; Penner, I.K.; Kallmann, B.A.; Lukas, C.; Kieseier, B.C. Towards the Implementation of ‘no Evidence of Disease Activity’ in Multiple Sclerosis Treatment: The Multiple Sclerosis Decision Model. Ther. Adv. Neurol. Disord. 2015, 8, 3–13. [Google Scholar] [CrossRef]
- Harris, V.K.; Sadiq, S.A. Biomarkers of Therapeutic Response in Multiple Sclerosis: Current Status. Mol. Diagn. Ther. 2014, 18, 605–617. [Google Scholar] [CrossRef]
- Aslam, S.; Khan, I.; Jameel, F.; Zaidi, M.B.; Salim, A. Umbilical Cord-Derived Mesenchymal Stem Cells Preconditioned with Isorhamnetin: Potential Therapy for Burn Wounds. World J. Stem Cells 2020, 12, 1652–1666. [Google Scholar] [CrossRef]
- Scopetti, M.; Santurro, A.; Gatto, V.; Russa, R.L.; Manetti, F.; Errico, S.D.; Frati, P.; Fineschi, V. Mesenchymal Stem Cells in Neurodegenerative Diseases: Opinion Review on Ethical Dilemmas. World J. Stem Cells 2020, 12, 168–177. [Google Scholar] [CrossRef]
- Khanabdali, R.; Saadat, A.; Fazilah, M.; Bazli, K.F.K.; Qazi, R.e.M.; Khalid, R.S.; Hasan Adli, D.S.; Moghadamtousi, S.Z.; Naeem, N.; Khan, I.; et al. Promoting Effect of Small Molecules in Cardiomyogenic and Neurogenic Differentiation of Rat Bone Marrow-Derived Mesenchymal Stem Cells. Drug Des. Devel. Ther. 2016, 10, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, A.; Alassiri, M.; Jawdat, D.; Almalik, Y. Mesenchymal Stem Cell Therapy: A Review of Clinical Trials for Multiple Sclerosis. Regen. Ther. 2022, 21, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Turano, E.; Scambi, I.; Virla, F.; Bonetti, B.; Mariotti, R. Extracellular Vesicles from Mesenchymal Stem Cells: Towards Novel Therapeutic Strategies for Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 2917. [Google Scholar] [CrossRef] [PubMed]
- Oppliger, B.; Joerger-Messerli, M.S.; Simillion, C.; Mueller, M.; Surbek, D.V.; Schoeberlein, A. Mesenchymal Stromal Cells from Umbilical Cord Wharton’s Jelly Trigger Oligodendroglial Differentiation in Neural Progenitor Cells through Cell-to-Cell Contact. Cytotherapy 2017, 19, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, A.; Khan, I.; Salim, A.; Qazi, R.E.M.; Malick, T.S.; Adli, D.S.H. Effect of α-Pinene and Thymoquinone on the Differentiation of Bone Marrow Mesenchymal Stem Cells into Neuroprogenitor Cells. BioImpacts 2022, 12, 147–154. [Google Scholar] [CrossRef]
- Sun, L.; Wang, F.; Chen, H.; Liu, D.; Qu, T.; Li, X.; Xu, D.; Liu, F.; Yin, Z.; Chen, Y. Co-Transplantation of Human Umbilical Cord Mesenchymal Stem Cells and Human Neural Stem Cells Improves the Outcome in Rats with Spinal Cord Injury. Cell Transplant. 2019, 28, 147–154. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chen, J.; Cui, Y.; Lu, M.; Elias, S.B.; Mitchell, J.B.; Hammill, L.; Vanguri, P.; Chopp, M. Human Bone Marrow Stromal Cell Treatment Improves Neurological Functional Recovery in EAE Mice. Exp. Neurol. 2005, 195, 16–26. [Google Scholar] [CrossRef]
- Shiri, E.; Pasbakhsh, P.; Borhani-Haghighi, M.; Alizadeh, Z.; Nekoonam, S.; Mojaverrostami, S.; Pirhajati Mahabadi, V.; Mehdi, A.; Zibara, K.; Kashani, I.R. Mesenchymal Stem Cells Ameliorate Cuprizone-Induced Demyelination by Targeting Oxidative Stress and Mitochondrial Dysfunction. Cell. Mol. Neurobiol. 2021, 41, 1467–1481. [Google Scholar] [CrossRef]
- Coursimault, J.; Guerrot, A.M.; Morrow, M.M.; Schramm, C.; Zamora, F.M.; Shanmugham, A.; Liu, S.; Zou, F.; Bilan, F.; Le Guyader, G.; et al. MYT1L-Associated Neurodevelopmental Disorder: Description of 40 New Cases and Literature Review of Clinical and Molecular Aspects. Hum. Genet. 2022, 141, 65–80. [Google Scholar] [CrossRef]
- Tahmasebi, F.; Pasbakhsh, P.; Barati, S.; Madadi, S.; Kashani, I.R. The Effect of Microglial Ablation and Mesenchymal Stem Cell Transplantation on a Cuprizone-Induced Demyelination Model. J. Cell. Physiol. 2021, 236, 3552–3564. [Google Scholar] [CrossRef]
- Popescu, B.F.G.; Lucchinetti, C.F. Pathology of Demyelinating Diseases. Annu. Rev. Pathol. Mech. Dis. 2012, 7, 185–217. [Google Scholar] [CrossRef] [PubMed]
- Matjusaitis, M.; Wagstaff, L.J.; Martella, A.; Baranowski, B.; Blin, C.; Gogolok, S.; Williams, A.; Pollard, S.M. Reprogramming of Fibroblasts to Oligodendrocyte Progenitor-like Cells Using CRISPR/Cas9-Based Synthetic Transcription Factors. Stem Cell Rep. 2019, 13, 1053–1067. [Google Scholar] [CrossRef]
- Khalid, S.; Ekram, S.; Salim, A.; Chaudhry, G.R.; Khan, I. Transcription Regulators Differentiate Mesenchymal Stem Cells into Chondroprogenitors, and Their in Vivo Implantation Regenerated the Intervertebral Disc Degeneration. World J. Stem Cells 2022, 14, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Ekram, S.; Khalid, S.; Bashir, I.; Salim, A.; Khan, I. Human Umbilical Cord-Derived Mesenchymal Stem Cells and Their Chondroprogenitor Derivatives Reduced Pain and Inflammation Signaling and Promote Regeneration in a Rat Intervertebral Disc Degeneration Model. Mol. Cell. Biochem. 2021, 476, 3191–3205. [Google Scholar] [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef] [PubMed]
- Mojaverrostami, S.; Khadivi, F.; Zarini, D.; Mohammadi, A. Combination Effects of Mesenchymal Stem Cells Transplantation and Anodal Transcranial Direct Current Stimulation on a Cuprizone-Induced Mouse Model of Multiple Sclerosis. J. Mol. Histol. 2022, 53, 817–831. [Google Scholar] [CrossRef]
- Bojnordi, M.N.; Ghasemi, H.H.; Akbari, E. Remyelination after Lysophosphatidyl Choline-Induced Demyelination Is Stimulated by Bone Marrow Stromal Cell-Derived Oligoprogenitor Cell Transplantation. Cells Tissues Organs 2015, 200, 300–306. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Z.; Lu, Z.; Borlongan, C.; Pan, J.; Chen, J.; Qian, L.; Liu, Z.; Zhu, L.; Zhang, J.; et al. Human Umbilical Cord Stem Cells Ameliorate Experimental Autoimmune Encephalomyelitis by Regulating Immunoinflammation and Remyelination. Stem Cells Dev. 2013, 22, 1053–1062. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, S.; Yang, Q.; Guo, S.; Chen, Q.; Liu, Z.; Li, L.; Jiang, M.; Li, H.; Hu, J.; et al. The Oligodendrocyte Transcription Factor 2 OLIG2 Regulates Transcriptional Repression during Myelinogenesis in Rodents. Nat. Commun. 2022, 13, 1423. [Google Scholar] [CrossRef]
- Xing, L.; Chai, R.; Wang, J.; Lin, J.; Li, H.; Wang, Y.; Lai, B.; Sun, J.; Chen, G. Expression of Myelin Transcription Factor 1 and Lamin B Receptor Mediate Neural Progenitor Fate Transition in the Zebrafish Spinal Cord PMN Domain. J. Biol. Chem. 2022, 298, 102452. [Google Scholar] [CrossRef]
- Shi, Y.; Shao, Q.; Li, Z.; Gonzalez, G.A.; Lu, F.; Wang, D.; Pu, Y.; Huang, A.; Zhao, C.; He, C.; et al. Myt1L Promotes Differentiation of Oligodendrocyte Precursor Cells and Is Necessary for Remyelination After Lysolecithin-Induced Demyelination. Neurosci. Bull. 2018, 34, 247–260. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Primary Antibody | Function/ Binding to | Working Dilution | Catalog Number | Manufacturer |
|---|---|---|---|---|---|
| MSC Characterization Markers | |||||
| 1. | CD29 | Membrane glycoprotein | 1:100 | MAB-1981 | Chemicon International, Katy, TX, USA |
| 2. | CD105 | Endoglin | 1:100 | 560839 | BD Pharmingen, San Diego, CA, USA |
| 3. | Vimentin | Epithelial–mesenchymal transition | 1:100 | V6389 | Sigma-Aldrich, Inc., St. Louis, MI, USA |
| 4. | C-Kit (CD117) | Stem cell factor receptor | 1:100 | 32–9000 | Zymed Laboratories, Inc., South San Francisco, CA, USA |
| 5. | Lin28 | Cell surface MSC marker | 1:100 | PA1-096 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 6. | Stro-1 | Mesenchymal precursor cell marker | 1:100 | 14-6688-82 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 7. | HLA-DR | MHC class II immunogenic marker | 1:100 | 14-9956-82 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 8. | CD45 | Lymphocyte antigen | 1:100 | CBL415 | BD Pharmingen, Diego, CA, USA |
| Oligodendrocyte-specific Markers | |||||
| 9. | OLIG2 | Oligodendrocyte lineage-specific marker | 1:100 | PA5-85734 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 10. | Myt1L | Myelin transcription factor 1-like | 1:50 | PA5-34468 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 11. | NG2 | Neural/Glial antigen 2 | 1:100 | PA5-100235 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 12. | MBP | Myelin basic protein | 1:100 | MA1-24990 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| Secondary Antibodies | |||||
| 13. | Goat Anti-rabbit | Alexa Fluor 546 | 1:200 | A-11010 | Molecular Probes, Invitrogen, Eugene, OR, USA |
| 14. | Anti-Rat IgG Isotype | Alexa Fluor 488 | 1:200 | 012-090-003 | Jackson Immuno Research, Inc., West Grove, PA, USA |
| Experimental Groups | Transcription Factor(s) Inserted |
|---|---|
| Normal Medium | |
| Control | -- |
| OLIG2-transfected | OLIG2 |
| MYT1L-transfected | MYT1L |
| Synergistic | OLIG2 + MYT1L |
| Oligo Induction Medium | |
| Control | -- |
| OLIG2-transfected | OLIG2 |
| MYT1L-transfected | MYT1L |
| Synergistic | OLIG2 + MYT1L |
| Genes | Primer Sequences (5′-3′) | Annealing Temperature | |
|---|---|---|---|
| Lineage-specific Genes | NES (F) NES (R) | TTCCAGACTCCACTCCCCTG CTCAGTCCCCAGGTCCTCAA | 55 °C |
| GFAP (F) GFAP (R) | ATGCTGGCTTCAAGGAGACC GGTGGCTTCATCTGCTTCCT | 55 °C | |
| OLIG2 (F) OLIG2 (R) | TCAAGTCATCCTCGTCCAGC TCACCAGTCGCTTCATCTCC | 55 °C | |
| Oligodendrocyte-specific Gene | MYT1L (F) MYT1L (R) | GACTGCGGAACAGGATTTGG CGACCAGGGTTTGAAGATGC | 55 °C |
| NKX2.2 (F) NKX2.2 (R) | TTCCTCGCCACCAGCC TTCGGCCACAGAGCCC | 55 °C | |
| SOX10 (F) SOX10 (R) | ACGTCAAGCGGCCCAT TCCCACCTTGCTCGGC | 55 °C | |
| GALC (F) GALC (R) | GAATTTTCCAAAGAATGGCTGGG CAGTGATGATCAAGTTACTGCCA | 55 °C | |
| CNP (F) CNP (R) | CCTTCAAGAAGGAGCTGCGA AGCTTGTCCACATCACTCGG | 55 °C | |
| CSPG4 (F) CSPG4 (R) | GGATGCCACCCTACAAGTGA TTTTGCGCCTCTAGTGGGAT | 55 °C | |
| PLP1 (F) PLP1 (R) | ATTCTTTGGAGCGGGTGTGT GAAGGTGAGCAGGGAAACCA | 55 °C | |
| MBP (F) MBP (R) | GCGGCCCCTGTCTCC GCGGCTCCCTGGGTC | 55 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahim, I.; Ishaque, A.; Ramzan, F.; Shamsuddin, S.A.B.A.; Ali, A.; Salim, A.; Khan, I. Overexpression of OLIG2 and MYT1L Transcription Factors Enhance the Differentiation Potential of Human Mesenchymal Stem Cells into Oligodendrocytes. Curr. Issues Mol. Biol. 2023, 45, 4100-4123. https://doi.org/10.3390/cimb45050261
Fahim I, Ishaque A, Ramzan F, Shamsuddin SABA, Ali A, Salim A, Khan I. Overexpression of OLIG2 and MYT1L Transcription Factors Enhance the Differentiation Potential of Human Mesenchymal Stem Cells into Oligodendrocytes. Current Issues in Molecular Biology. 2023; 45(5):4100-4123. https://doi.org/10.3390/cimb45050261
Chicago/Turabian StyleFahim, Ifrah, Aisha Ishaque, Faiza Ramzan, Shamsul Azlin Bin Ahmad Shamsuddin, Anwar Ali, Asmat Salim, and Irfan Khan. 2023. "Overexpression of OLIG2 and MYT1L Transcription Factors Enhance the Differentiation Potential of Human Mesenchymal Stem Cells into Oligodendrocytes" Current Issues in Molecular Biology 45, no. 5: 4100-4123. https://doi.org/10.3390/cimb45050261
APA StyleFahim, I., Ishaque, A., Ramzan, F., Shamsuddin, S. A. B. A., Ali, A., Salim, A., & Khan, I. (2023). Overexpression of OLIG2 and MYT1L Transcription Factors Enhance the Differentiation Potential of Human Mesenchymal Stem Cells into Oligodendrocytes. Current Issues in Molecular Biology, 45(5), 4100-4123. https://doi.org/10.3390/cimb45050261