

The Epigenetic Reader Protein SP140 Regulates Dendritic Cell Activation, Maturation and Tolerogenic Potential

 , , , , and
, , , , and 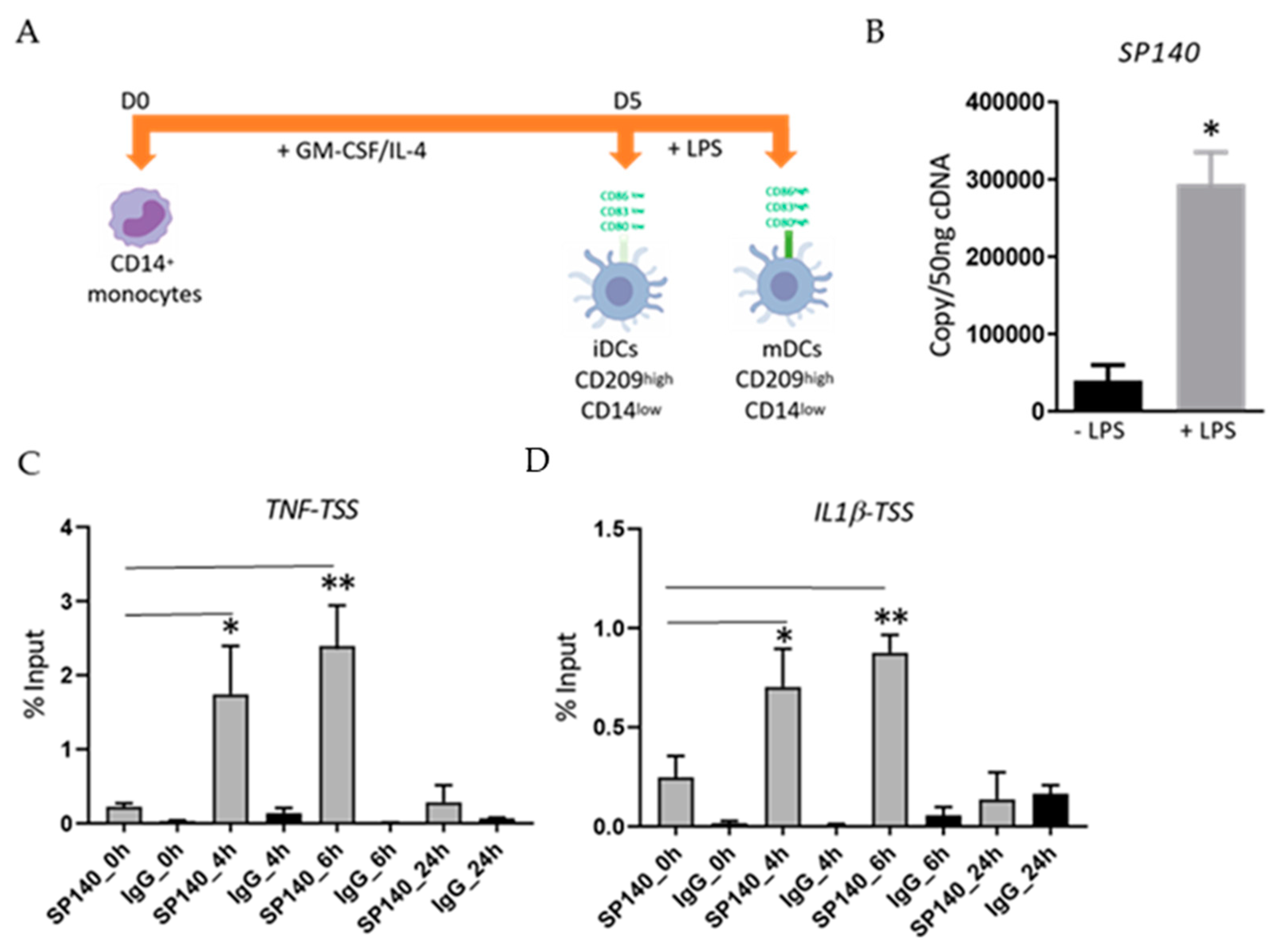{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LPS-Stimulation Induces an Increase in SP140 Expression in DCs as Well as Its Recruitment to TSS of Pro-Inflammatory Cytokine Genes
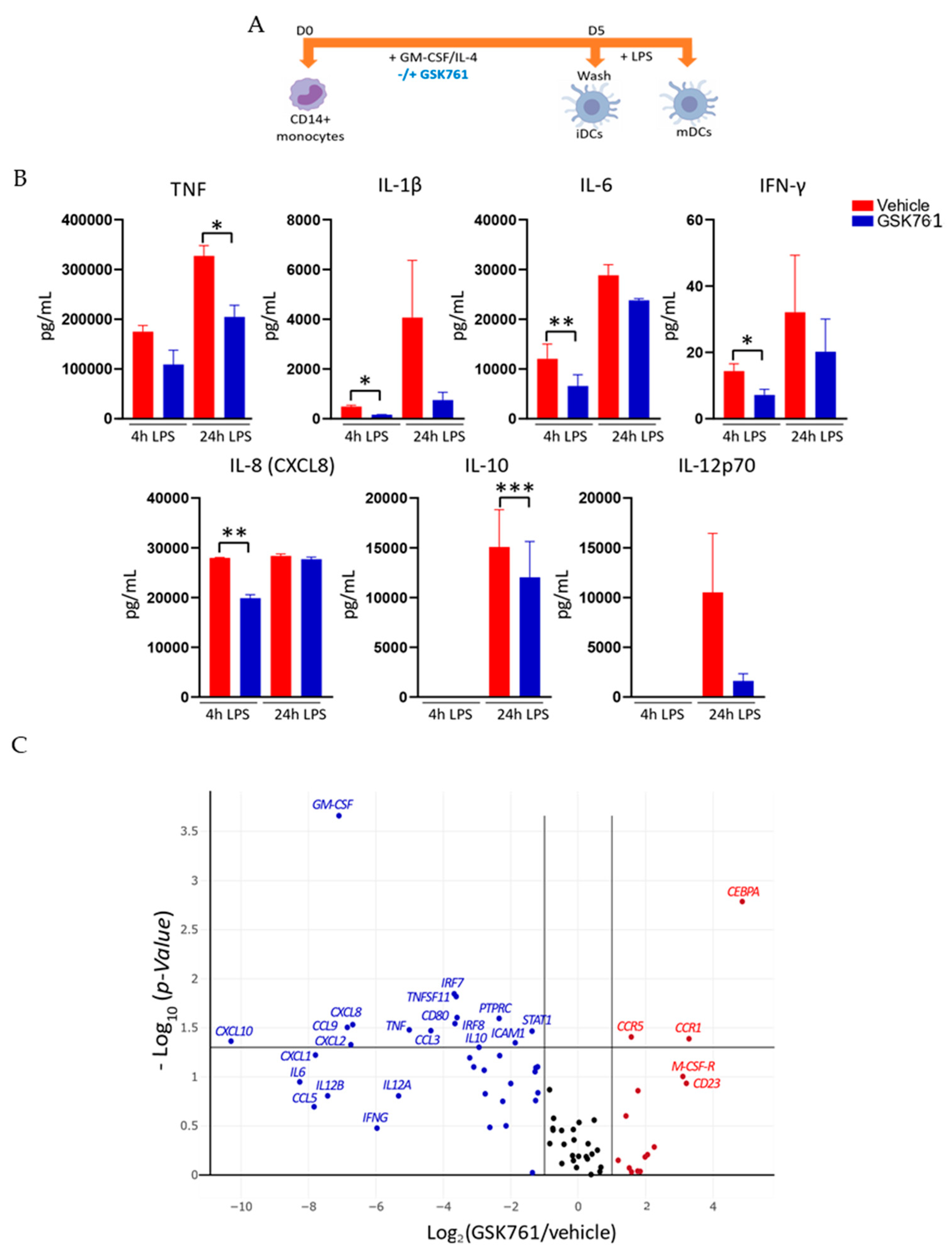
2.2. GSK761 Attenuates the Inflammatory Activation of DCs
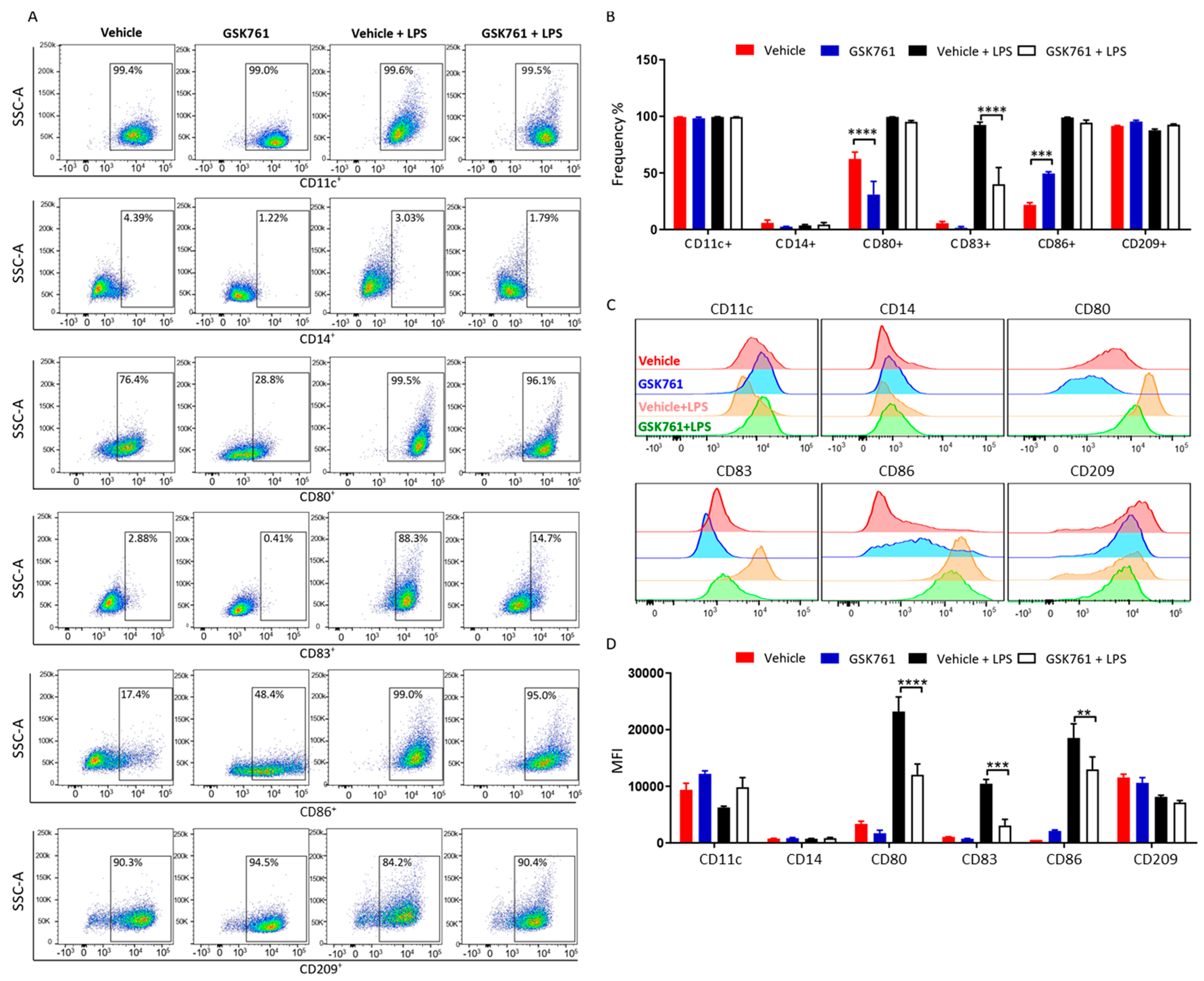
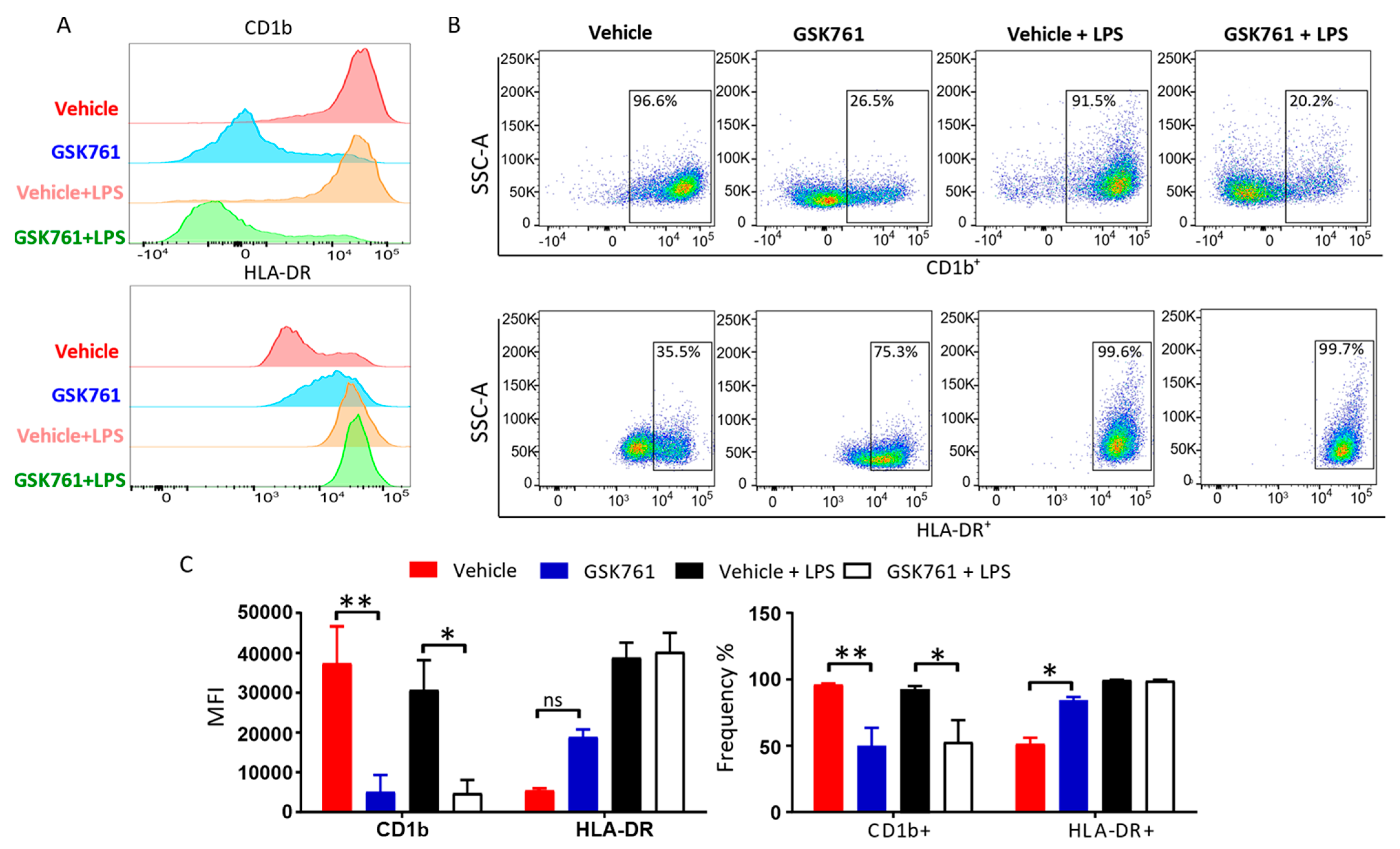
2.3. SP140 Is Critical for DCs Maturation
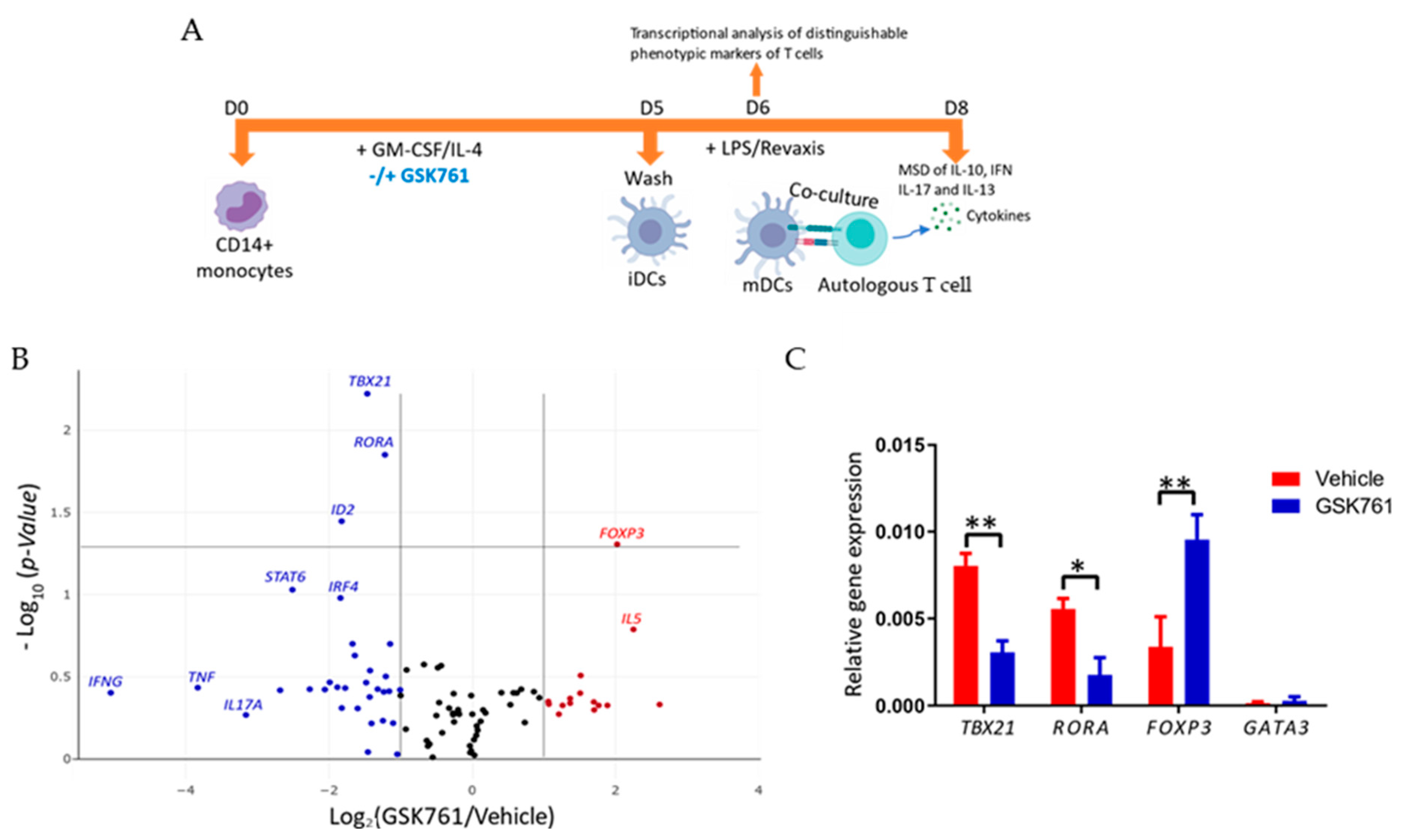
2.4. GSK761-Treatment of DCs Decreases Revaxis-Induced TBX21 and Increases FOXP3 Expression in Autologous T Cells
3. Discussion
4. Materials and Methods
4.1. CD14+ Monocyte Isolation, In Vitro Generation of DCs and SP140 Inhibition
4.2. siRNA-Mediated SP140 Knockdown
4.3. Isolation of Human Autologous CD4+ T Cells and DCs-T Cells Co-Culture
4.4. Chromatin Immunoprecipitation (ChIP)
4.5. Cytotoxicity Assay
4.6. T2 Profiler qPCR-Array and qPCR
4.7. Cytokine Analysis
4.8. Flow Cytometry (FACS)
4.9. Statistics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lewis, K.L.; Reizis, B. Dendritic Cells: Arbiters of Immunity and Immunological Tolerance. Cold Spring Harb. Perspect. Biol. 2012, 4, a007401. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Manz, M.G. Dendritic cell homeostasis. Blood 2009, 113, 3418–3427. [Google Scholar] [CrossRef] [PubMed]
- Wardowska, A.; Komorniczak, M.; Bullo-Piontecka, B.; Dȩbska-Ślizień, M.A.; Pikuła, M. Transcriptomic and Epigenetic Alterations in Dendritic Cells Correspond with Chronic Kidney Disease in Lupus Nephritis. Front. Immunol. 2019, 10, 2026. [Google Scholar] [CrossRef] [PubMed]
- Nencioni, A.; Beck, J.; Werth, D.; Grünebach, F.; Patrone, F.; Ballestrero, A.; Brossart, P. Histone deacetylase inhibitors affect dendritic cell differentiation and immunogenicity. Clin. Cancer Res. 2007, 13, 3933–3941. [Google Scholar] [CrossRef]
- Brogdon, J.L.; Xu, Y.; Szabo, S.J.; An, S.; Buxton, F.; Cohen, D.; Huang, Q. Histone deacetylase activities are required for innate immune cell control of Th1 but not Th2 effector cell function. Blood 2006, 109, 1123–1130. [Google Scholar] [CrossRef]
- Ge, Z.; Da, Y.; Xue, Z.; Zhang, K.; Zhuang, H.; Peng, M.; Li, Y.; Li, W.; Simard, A.; Hao, J.; et al. Vorinostat, a histone deacetylase inhibitor, suppresses dendritic cell function and ameliorates experimental autoimmune encephalomyelitis. Exp. Neurol. 2013, 241, 56–66. [Google Scholar] [CrossRef]
- Woo, S.J.; Lee, S.-M.; Lim, H.S.; Hah, Y.-S.; Jung, I.D.; Park, Y.-M.; Kim, H.-O.; Cheon, Y.-H.; Jeon, M.-G.; Jang, K.Y.; et al. Myeloid deletion of SIRT1 suppresses collagen-induced arthritis in mice by modulating dendritic cell maturation. Exp. Mol. Med. 2016, 48, e221. [Google Scholar] [CrossRef]
- Elmesmari, A.; Alivernini, S.; Tolusso, B.; Bui, L.; Vaughan, D.; Gigante, M.; Federico, F.; Ferraccioli, G.; Gremese, E.; McInnes, I.; et al. SAT0003 Synovial tissue cd1c+ dendritic cells in rheumatoid arthritis express high levels of the epigenetic regulator of inflammation, microrna-155 and inflammatory cytokines. Ann. Rheum. Dis. 2018, 77, 870–871. [Google Scholar] [CrossRef]
- Fraschilla, I.; Jeffrey, K.L. The Speckled Protein (SP) Family: Immunity’s Chromatin Readers. Trends Immunol. 2020, 41, 572–585. [Google Scholar] [CrossRef]
- Ghiboub, M.; Koster, J.; Craggs, P.D.; Yim, A.Y.F.L.; Shillings, A.; Hutchinson, S.; Bingham, R.P.; Gatfield, K.; Hageman, I.L.; Yao, G.; et al. Modulation of macrophage inflammatory function through selective inhibition of the epigenetic reader protein SP140. BMC Biol. 2022, 20, 182. [Google Scholar] [CrossRef]
- Matesanz, F.; Potenciano, V.; Fedetz, M.; Ramos-Mozo, P.; Abad-Grau, M.D.M.; Karaky, M.; Barrionuevo, C.; Izquierdo, G.; Ruiz-Peña, J.L.; García-Sánchez, M.I.; et al. A functional variant that affects exon-skipping and protein expression of SP140 as genetic mechanism predisposing to multiple sclerosis. Hum. Mol. Genet. 2015, 24, 5619–5627. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Cronkite, D.A.; Basavappa, M.; Saunders, T.L.; Adiliaghdam, F.; Amatullah, H.; Morrison, S.A.; Pagan, J.D.; Anthony, R.M.; Tonnerre, P.; et al. Maintenance of macrophage transcriptional programs and intestinal homeostasis by epigenetic reader SP140. Sci. Immunol. 2017, 2, aag3160. [Google Scholar] [CrossRef] [PubMed]
- Zucchelli, C.; Tamburri, S.; Filosa, G.; Ghitti, M.; Quilici, G.; Bachi, A.; Musco, G. Sp140 is a multi-SUMO-1 target and its PHD finger promotes SUMOylation of the adjacent Bromodomain. Biochim. Biophys. Acta Gen. Subj. 2018, 1863, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, G.; Gao, X. Development of neurodevelopmental disorders: A regulatory mechanism involving bromodomain-containing proteins. J. Neurodev. Disord. 2013, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Ghiboub, M.; Elfiky, A.M.I.; de Winther, M.P.J.; Harker, N.R.; Tough, D.F.; de Jonge, W.J. Selective Targeting of Epigenetic Readers and Histone Deacetylases in Autoimmune and Inflammatory Diseases: Recent Advances and Future Perspectives. J. Pers. Med. 2021, 11, 336. [Google Scholar] [CrossRef]
- Coutant, F.; Miossec, P. Altered dendritic cell functions in autoimmune diseases: Distinct and overlapping profiles. Nat. Rev. Rheumatol. 2016, 12, 703–715. [Google Scholar] [CrossRef]
- Schilderink, R.; Bell, M.; Reginato, E.; Patten, C.; Rioja, I.; Hilbers, F.W.; Kabala, P.A.; Reedquist, K.A.; Tough, D.F.; Tak, P.P.; et al. BET bromodomain inhibition reduces maturation and enhances tolerogenic properties of human and mouse dendritic cells. Mol. Immunol. 2016, 79, 66–76. [Google Scholar] [CrossRef]
- Toniolo, P.A.; Liu, S.; Yeh, J.E.; Moraes-Vieira, P.M.; Walker, S.R.; Vafaizadeh, V.; Barbuto, J.A.M.; Frank, D.A. Inhibiting STAT5 by the BET Bromodomain Inhibitor JQ1 Disrupts Human Dendritic Cell Maturation. J. Immunol. 2015, 194, 3180–3190. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Toubai, T.; Oravecz-Wilson, K.; Liu, C.; Mathewson, N.; Wu, J.; Rossi, C.; Cummings, E.; Wu, D.; et al. BET bromodomain inhibition suppresses graft-versus-host disease after allogeneic bone marrow transplantation in mice. Blood 2015, 125, 2724–2728. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef]
- Romani, N.; Gruner, S.; Brang, D.; Kämpgen, E.; Lenz, A.; Trockenbacher, B.; Konwalinka, G.; Fritsch, P.O.; Steinman, R.M.; Schuler, G. Proliferating dendritic cell progenitors in human blood. J. Exp. Med. 1994, 180, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Hiasa, M.; Abe, M.; Nakano, A.; Oda, A.; Amou, H.; Kido, S.; Takeuchi, K.; Kagawa, K.; Yata, K.; Hashimoto, T.; et al. GM-CSF and IL-4 induce dendritic cell differentiation and disrupt osteoclastogenesis through M-CSF receptor shedding by up-regulation of TNF-α converting enzyme (TACE). Blood 2009, 114, 4517–4526. [Google Scholar] [CrossRef] [PubMed]
- Chometon, T.Q.; Siqueira, M.D.S.; Sant’anna, J.C.; Almeida, M.R.; Gandini, M.; Nogueira, A.C.M.D.A.; Antas, P.R.Z. A protocol for rapid monocyte isolation and generation of singular human monocyte-derived dendritic cells. PLoS ONE 2020, 15, e0231132. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Lee, S.H.; Lu, J. CD83 is preformed inside monocytes, macrophages and dendritic cells, but it is only stably expressed on activated dendritic cells. Biochem. J. 2004, 385, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.H.; Yu, C.-R.; Mahdi, R.M.; Ebong, S.; Egwuagu, C.E. Dendritic Cell Maturation Requires STAT1 and Is under Feedback Regulation by Suppressors of Cytokine Signaling. J. Immunol. 2004, 172, 2307–2315. [Google Scholar] [CrossRef]
- Sallusto, F.; Schaerli, P.; Loetscher, P.; Schaniel, C.; Lenig, D.; Mackay, C.R.; Qin, S.; Lanzavecchia, A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur. J. Immunol. 1998, 28, 2760–2769. [Google Scholar] [CrossRef]
- Shahine, A. The intricacies of self-lipid antigen presentation by CD1b. Mol. Immunol. 2018, 104, 27–36. [Google Scholar] [CrossRef]
- ten Broeke, T.; Wubbolts, R.; Stoorvogel, W. MHC class II antigen presentation by dendritic cells regulated through endosomal sorting. Cold Spring Harb. Perspect. Biol. 2013, 5, a016873. [Google Scholar] [CrossRef]
- Gajdos, V.; Soubeyrand, B.; Vidor, E.; Richard, P.; Boyer, J.; Sadorge, C.; Fiquet, A. Immunogenicity and safety of combined adsorbed low-dose diphtheria, tetanus and inactivated poliovirus vaccine (REVAXIS (®®)) versus combined diphtheria, tetanus and inactivated poliovirus vaccine (DT Polio (®®)) given as a booster dose at 6 years of age. Hum. Vaccin. 2011, 7, 549–556. [Google Scholar] [CrossRef]
- Rybczynska, M.; Baudry, J.; Klaus, E. The impact of frost-damage on the quality and quantity of the secreted antigen-specific IgG repertoire. Vaccine 2020, 38, 5337–5342. [Google Scholar] [CrossRef]
- Roberts, C.A.; Durham, L.E.; Fleskens, V.; Evans, H.G.; Taams, L.S. TNF Blockade Maintains an IL-10+ Phenotype in Human Effector CD4+ and CD8+ T Cells. Front. Immunol. 2017, 8, 157. [Google Scholar] [CrossRef]
- Shaw, L.A.; Bélanger, S.; Omilusik, K.D.; Cho, S.; Scott-Browne, J.P.; Nance, J.P.; Goulding, J.; Lasorella, A.; Lu, L.-F.; Crotty, S.; et al. Id2 reinforces TH1 differentiation and inhibits E2A to repress TFH differentiation. Nat. Immunol. 2016, 17, 834–843. [Google Scholar] [CrossRef]
- Kanhere, A.; Hertweck, A.; Bhatia, U.; Gökmen, M.R.; Perucha, E.; Jackson, I.; Lord, G.M.; Jenner, R.G. T-bet and GATA3 orchestrate Th1 and Th2 differentiation through lineage-specific targeting of distal regulatory elements. Nat. Commun. 2012, 3, 1268. [Google Scholar] [CrossRef]
- Unutmaz, D. RORC2: The master of human Th17 cell programming. Eur. J. Immunol. 2009, 39, 1452–1455. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Tsun, A.; Li, B. FOXP3+ regulatory T cells and their functional regulation. Cell Mol. Immunol. 2015, 12, 558–565. [Google Scholar] [CrossRef]
- Fragale, A.; Gabriele, L.; Stellacci, E.; Borghi, P.; Perrotti, E.; Ilari, R.; Lanciotti, A.; Remoli, A.L.; Venditti, M.; Belardelli, F.; et al. IFN Regulatory Factor-1 Negatively Regulates CD4+CD25+ Regulatory T Cell Differentiation by Repressing Foxp3 Expression. J. Immunol. 2008, 181, 1673–1682. [Google Scholar] [CrossRef]
- Hartenstein, B.; Teurich, S.; Hess, J.; Schenkel, J.; Schorpp-Kistner, M.; Angel, P. Th2 cell-specific cytokine expression and allergen-induced airway inflammation depend on JunB. EMBO J. 2002, 21, 6321–6329. [Google Scholar] [CrossRef]
- O’garra, A.; Arai, N. The molecular basis of T helper 1 and T helper 2 cell differentiation. Trends Cell Biol. 2000, 10, 542–550. [Google Scholar] [CrossRef]
- Britt, R.D.; Thompson, M.A.; Sasse, S.K.; Pabelick, C.M.; Gerber, A.N.; Prakash, Y.S. Th1 cytokines TNF-α and IFN-γ promote corticosteroid resistance in developing human airway smooth muscle. Am. J. Physiol. Cell Mol. Physiol. 2019, 316, L71–L81. [Google Scholar] [CrossRef]
- Annunziato, F.; Cosmi, L.; Santarlasci, V.; Maggi, L.; Liotta, F.; Mazzinghi, B.; Parente, E.; Filì, L.; Ferri, S.; Frosali, F.; et al. Phenotypic and functional features of human Th17 cells. J. Exp. Med. 2007, 204, 1849–1861. [Google Scholar] [CrossRef]
- Alqahtani, A.; Choucair, K.; Ashraf, M.; Hammouda, D.M.; Alloghbi, A.; Khan, T.; Senzer, N.; Nemunaitis, J. Bromodomain and extra-terminal motif inhibitors: A review of preclinical and clinical advances in cancer therapy. Future Sci. OA 2019, 5, FSO372. [Google Scholar] [CrossRef]
- Andrieu, G.; Belkina, A.C.; Denis, G.V. Clinical trials for BET inhibitors run ahead of the science. Drug Discov. Today Technol. 2016, 19, 45–50. [Google Scholar] [CrossRef]
- Wehr, P.; Purvis, H.; Law, S.; Thomas, R. Dendritic cells, T cells and their interaction in rheumatoid arthritis. Clin. Exp. Immunol. 2018, 196, 12–27. [Google Scholar] [CrossRef]
- Knight, S.C. Dendritic Cell-T-Cell Circuitry in Health and Changes in Inflammatory Bowel Disease and Its Treatment. Dig. Dis. 2016, 34, 51–57. [Google Scholar] [CrossRef]
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef]
- Vendelova, E.; Ashour, D.; Blank, P.; Erhard, F.; Saliba, A.-E.; Kalinke, U.; Lutz, M.B. Tolerogenic Transcriptional Signatures of Steady-State and Pathogen-Induced Dendritic Cells. Front. Immunol. 2018, 9, 333. [Google Scholar] [CrossRef]
- Tze, L.E.; Horikawa, K.; Domaschenz, H.; Howard, D.R.; Roots, C.M.; Rigby, R.; Way, D.A.; Ohmura-Hoshino, M.; Ishido, S.; Andoniou, C.; et al. CD83 increases MHC II and CD86 on dendritic cells by opposing IL-10–driven MARCH1-mediated ubiquitination and degradation. J. Exp. Med. 2011, 208, 149–165. [Google Scholar] [CrossRef]
- Xu, J.-F.; Huang, B.-J.; Yin, H.; Xiong, P.; Feng, W.; Xu, Y.; Fang, M.; Zheng, F.; Wang, C.-Y.; Gong, F.-L. A limited course of soluble CD83 delays acute cellular rejection of MHC-mismatched mouse skin allografts. Transpl. Int. 2006, 20, 266–276. [Google Scholar] [CrossRef]
- Lan, Z.; Ge, W.; Arp, J.; Jiang, J.; Liu, W.; Gordon, D.; Healey, D.; Debenedette, M.; Nicolette, C.; Garcia, B.; et al. Induction of Kidney Allograft Tolerance by Soluble CD83 Associated with Prevalence of Tolerogenic Dendritic Cells and Indoleamine 2,3-Dioxygenase. Transplantation 2010, 90, 1286–1293. [Google Scholar] [CrossRef]
- Eckhardt, J.; Kreiser, S.; Döbbeler, M.; Nicolette, C.; DeBenedette, M.A.; Tcherepanova, I.Y.; Ostalecki, C.; Pommer, A.J.; Becker, C.; Günther, C.; et al. Soluble CD83 ameliorates experimental colitis in mice. Mucosal. Immunol. 2014, 7, 1006–1018. [Google Scholar] [CrossRef]
- Royzman, D.; Andreev, D.; Stich, L.; Rauh, M.; Baeuerle, T.; Ellmann, S.; Boon, L.; Kindermann, M.; Peckert, K.; Bozec, A.; et al. Soluble CD83 Triggers Resolution of Arthritis and Sustained Inflammation Control in IDO Dependent Manner. Front. Immunol. 2019, 10, 633. [Google Scholar] [CrossRef]
- Li, Z.; Ju, X.; Silveira, P.A.; Abadir, E.; Hsu, W.-H.; Hart, D.N.J.; Clark, G. CD83: Activation Marker for Antigen Presenting Cells and Its Therapeutic Potential. Front. Immunol. 2019, 10, 1312. [Google Scholar] [CrossRef]
- Skapenko, A.; Leipe, J.; Lipsky, P.E.; Schulze-Koops, H. The role of the T cell in autoimmune inflammation. Arthritis Res. Ther. 2005, 7, S4–S14. [Google Scholar] [CrossRef]
- Korhonen, R.; Moilanen, E. Abatacept, a novel CD80/86-CD28 T cell co-stimulation modulator, in the treatment of rheumatoid arthritis. Basic Clin. Pharmacol. Toxicol. 2009, 104, 276–284. [Google Scholar] [CrossRef]
- Noisette, A.; Hochberg, M.C. Abatacept for the treatment of adults with psoriatic arthritis: Patient selection and perspectives. Psoriasis Targets Ther. 2018, 8, 31–39. [Google Scholar] [CrossRef]
- De Vries, L.C.; Ghiboub, M.; van Hamersveld, P.H.; Welting, O.; Verseijden, C.; Bell, M.J.; Rioja, I.; Prinjha, R.K.; Koelink, P.J.; Strobl, B.; et al. Tyrosine kinase 2 signalling drives pathogenic T-cells in colitis. J. Crohn’s Colitis 2021, 15, 617–630. [Google Scholar] [CrossRef]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Bine, S.; Haziot, A.; Malikova, I.; Pelletier, J.; Charron, D.; Boucraut, J.; Mooney, N.; Gelin, C. Alteration of CD1 expression in multiple sclerosis. Clin. Exp. Immunol. 2012, 169, 10–16. [Google Scholar] [CrossRef]
- Olivier, M.; Foret, B.; Le Vern, Y.; Kerboeuf, D.; Guilloteau, L.A. Plasticity of Migrating CD1b+ and CD1b- Lymph Dendritic Cells in the Promotion of Th1, Th2 and Th17 in Response to Salmonella and Helminth Secretions. PLoS ONE 2013, 8, e79537. [Google Scholar] [CrossRef]
- Di Rosa, F.; Cossarizza, A.; Hayday, A.C. To Ki or Not to Ki: Re-Evaluating the Use and Potentials of Ki-67 for T Cell Analysis. Front. Immunol. 2021, 12, 653974. [Google Scholar] [CrossRef]
- Huang, Y.; Min, S.; Lui, Y.; Sun, J.; Su, X.; Liu, Y.; Zhang, Y.; Han, D.; Che, Y.; Zhao, C.; et al. Global mapping of H3K4me3 and H3K27me3 reveals chromatin state-based regulation of human monocyte-derived dendritic cells in different environments. Genes Immun. 2012, 13, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Węsierska-Gądek, J.; Gueorguieva, M.; Ranftler, C.; Zerza-Schnitzhofer, G. A new multiplex assay allowing simultaneous detection of the inhibition of cell proliferation and induction of cell death. J. Cell Biochem. 2005, 96, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhelev, Z.; Ohba, H.; Bakalova, R.; Hadjimitova, V.; Ishikawa, M.; Shinohara, Y.; Baba, Y. Phenothiazines suppress proliferation and induce apoptosis in cultured leukemic cells without any influence on the viability of normal lymphocytes. Cancer Chemother. Pharmacol. 2004, 53, 267–275. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghiboub, M.; Bell, M.; Sinkeviciute, D.; Prinjha, R.K.; de Winther, M.P.J.; Harker, N.R.; Tough, D.F.; de Jonge, W.J. The Epigenetic Reader Protein SP140 Regulates Dendritic Cell Activation, Maturation and Tolerogenic Potential. Curr. Issues Mol. Biol. 2023, 45, 4228-4245. https://doi.org/10.3390/cimb45050269
Ghiboub M, Bell M, Sinkeviciute D, Prinjha RK, de Winther MPJ, Harker NR, Tough DF, de Jonge WJ. The Epigenetic Reader Protein SP140 Regulates Dendritic Cell Activation, Maturation and Tolerogenic Potential. Current Issues in Molecular Biology. 2023; 45(5):4228-4245. https://doi.org/10.3390/cimb45050269
Chicago/Turabian StyleGhiboub, Mohammed, Matthew Bell, Dovile Sinkeviciute, Rab K. Prinjha, Menno P. J. de Winther, Nicola R. Harker, David F. Tough, and Wouter J. de Jonge. 2023. "The Epigenetic Reader Protein SP140 Regulates Dendritic Cell Activation, Maturation and Tolerogenic Potential" Current Issues in Molecular Biology 45, no. 5: 4228-4245. https://doi.org/10.3390/cimb45050269
APA StyleGhiboub, M., Bell, M., Sinkeviciute, D., Prinjha, R. K., de Winther, M. P. J., Harker, N. R., Tough, D. F., & de Jonge, W. J. (2023). The Epigenetic Reader Protein SP140 Regulates Dendritic Cell Activation, Maturation and Tolerogenic Potential. Current Issues in Molecular Biology, 45(5), 4228-4245. https://doi.org/10.3390/cimb45050269