

Mitogenomic Research of Silverleaf Sunflower (Helianthus argophyllus) and Its Interspecific Hybrids

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Mitochondrial DNA Extraction
2.2. Next-Generation Sequencing
2.3. Mitochondrial Genome Assembly and Annotation
2.4. SNV Localisation and RNA Editing Analysis
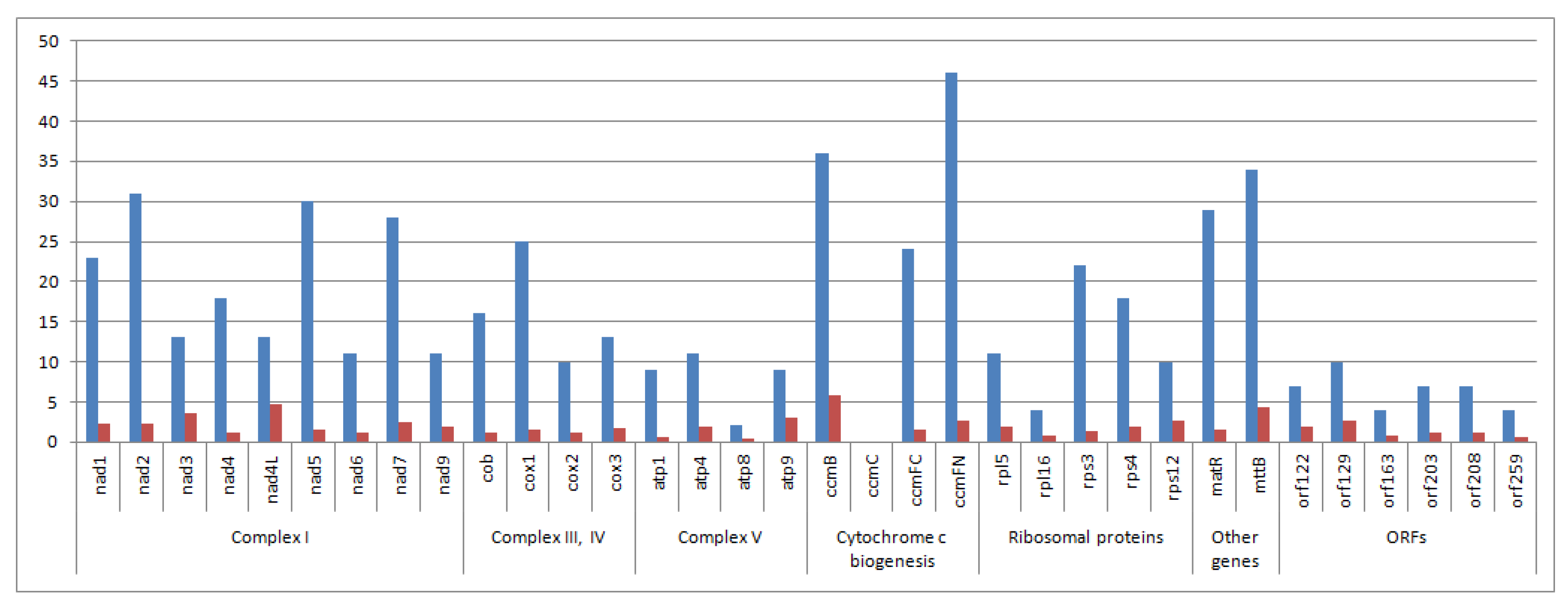
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timme, R.E.; Simpson, B.B.; Linder, C.R. High-Resolution Phylogeny for Helianthus (Asteraceae) Using the 18S-26S Ribosomal DNA External Transcribed Spacer. Am. J. Bot. 2007, 94, 1837–1852. [Google Scholar] [CrossRef]
- Moyers, B.T.; Rieseberg, L.H. Divergence in Gene Expression Is Uncoupled from Divergence in Coding Sequence in a Secondarily Woody Sunflower. Int. J. Plant Sci. 2015, 174, 1079–1089. [Google Scholar] [CrossRef]
- Seiler, G.J.; Jan, C.-C. Wild Sunflower Species as a Genetic Resource for Resistance to Sunflower Broomrape (Orobanche Cumana Wallr.). Helia 2014, 37, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Heiser, C.B., Jr. Hybridization in the Annual Sunflowers: Helianthus annuus × H. Argophyllus. Am. Nat. 1951, 85, 65–72. [Google Scholar] [CrossRef]
- Todesco, M.; Owens, G.L.; Bercovich, N.; Légaré, J.-S.; Soudi, S.; Burge, D.O.; Huang, K.; Ostevik, K.L.; Drummond, E.B.M.; Imerovski, I.; et al. Massive Haplotypes Underlie Ecotypic Differentiation in Sunflowers. Nature 2020, 584, 602–607. [Google Scholar] [CrossRef]
- Ostevik, K.L.; Samuk, K.; Rieseberg, L.H. Ancestral Reconstruction of Karyotypes Reveals an Exceptional Rate of Nonrandom Chromosomal Evolution in Sunflower. Genetics 2020, 214, 1031–1045. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Baack, E.J.; Whitney, K.D.; Bock, D.G.; Tetreault, H.M.; Rieseberg, L.H.; Ungerer, M.C. Phylogenetic Trends and Environmental Correlates of Nuclear Genome Size Variation in Helianthus Sunflowers. New Phytol. 2019, 221, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Strasburg, J.L.; Rieseberg, L.H. Molecular Demographic History of the Annual Sunflowers Helianthus annuus and H. petiolaris—Large Effective Population Sizes and Rates of Long-Term Gene Flow. Evolution 2008, 62, 1936–1950. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Andrew, R.L.; Owens, G.L.; Ostevik, K.L.; Rieseberg, L.H. Multiple Chromosomal Inversions Contribute to Adaptive Divergence of a Dune Sunflower Ecotype. Mol. Ecol. 2020, 29, 2535–2549. [Google Scholar] [CrossRef]
- Vekemans, X. What’s Good for You May Be Good for Me: Evidence for Adaptive Introgression of Multiple Traits in Wild Sunflower. New Phytol. 2010, 187, 6–9. [Google Scholar] [CrossRef]
- Gutierrez, A.; Carrera, A.; Basualdo, J.; Rodriguez, R.; Cantamutto, M.; Poverene, M. Gene Flow between Cultivated Sunflower and Helianthus petiolaris (Asteraceae). Euphytica 2010, 172, 67–76. [Google Scholar] [CrossRef]
- Barb, J.G.; Bowers, J.E.; Renaut, S.; Rey, J.I.; Knapp, S.J.; Rieseberg, L.H.; Burke, J.M. Chromosomal Evolution and Patterns of Introgression in Helianthus. Genetics 2014, 197, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzić, S.; Boniface, M.-C.; Marek, L.; Alvarez, D.; Baumann, K.; Gavrilova, V.; Joita-Pacureanu, M.; Sujatha, M.; Valkova, D.; Velasco, L.; et al. Gene Banks for Wild and Cultivated Sunflower Genetic Resources. OCL 2020, 27, 9. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.L.; Foley, M.E.; Cai, X.W.; Gulya, T.J. Genetics and Mapping of a Novel Downy Mildew Resistance Gene, Pl18, Introgressed from Wild Helianthus argophyllus into Cultivated Sunflower (Helianthus annuus L.). Appl. Genet. 2016, 129, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Kane, N.C.; Burke, J.M.; Marek, L.; Seiler, G.; Vear, F.; Baute, G.; Knapp, S.J.; Vincourt, P.; Rieseberg, L.H. Sunflower Genetic, Genomic and Ecological Resources. Mol. Ecol. Resour. 2013, 13, 10–20. [Google Scholar] [CrossRef]
- Dimitrijevic, A.; Horn, R. Sunflower Hybrid Breeding: From Markers to Genomic Selection. Front. Plant Sci. 2018, 8, 2238. [Google Scholar] [CrossRef] [Green Version]
- Christov, M. Helianthus Species in Breeding Research on Sunflower. In Proceedings of the 17th International Sunflower Conference, Cordoba, Spain, 8–12 June 2008; Volume 2, pp. 709–714. [Google Scholar]
- Gausman, H.W.; Menges, R.M.; Escobar, D.E.; Everitt, J.H.; Bowen, R.L. Pubescence Affects Spectra and Imagery of Silverleaf Sunflower (Helianthus argophyllus). Weed Sci. 1977, 25, 437–440. [Google Scholar] [CrossRef]
- Dußle, C.M.; Hahn, V.; Knapp, S.J.; Bauer, E. PlArgfrom Helianthus argophyllus Is Unlinked to Other Known Downy Mildew Resistance Genes in Sunflower. Appl. Genet. 2004, 109, 1083–1086. [Google Scholar] [CrossRef]
- Hussain, M.M.; Rauf, S.; Riaz, M.A.; Al-Khayri, J.M.; Monneveux, P. Determination of Drought Tolerance Related Traits in Helianthus argophyllus, Helianthus annuus, and Their Hybrids. Breed. Sci. 2017, 67, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sujatha, M.; Lakshminarayana, M. Resistance to Spodoptera litura (Fabr.) in Helianthus Species and Backcross Derived Inbred Lines from Crosses Involving Diploid Species. Euphytica 2007, 155, 205–213. [Google Scholar] [CrossRef]
- Tyagi, V.; Dhillon, S.K.; Kaur, G.; Kaushik, P. Heterotic Effect of Different Cytoplasmic Combinations in Sunflower Hybrids Cultivated Under Diverse Irrigation Regimes. Plants 2020, 9, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cytomorphological and Molecular Characterization of Inter-Specific Hybrid between Cultivated Sunflower and Helianthus argophyllus—ProQuest. Available online: https://www.proquest.com/openview/4378599eaf690c287546ca282f2b2d7e/1?pq-origsite=gscholar&cbl=636374 (accessed on 26 January 2023).
- Anisimova, I.N.; Tumanova, L.G.; Gavrilova, V.A.; Dyagileva, A.V.; Pasha, L.I.; Mitin, V.A.; Timofeyeva, G.I. Genomic Instability in Sunflower Interspecific Hybrids. Russ. J. Genet. 2009, 45, 934–943. [Google Scholar] [CrossRef]
- Liu, H.; Cui, P.; Zhan, K.; Lin, Q.; Zhuo, G.; Guo, X.; Ding, F.; Yang, W.; Liu, D.; Hu, S.; et al. Comparative Analysis of Mitochondrial Genomes between a Wheat K-Type Cytoplasmic Male Sterility (CMS) Line and Its Maintainer Line. BMC Genom. 2011, 12, 163. [Google Scholar] [CrossRef] [Green Version]
- Kozik, A.; Rowan, B.A.; Lavelle, D.; Berke, L.; Schranz, M.E.; Michelmore, R.W.; Christensen, A.C. The Alternative Reality of Plant Mitochondrial DNA: One Ring Does Not Rule Them All. PLoS Genet. 2019, 15, e1008373. [Google Scholar] [CrossRef] [Green Version]
- Gualberto, J.M.; Mileshina, D.; Wallet, C.; Niazi, A.K.; Weber-Lotfi, F.; Dietrich, A. The Plant Mitochondrial Genome: Dynamics and Maintenance. Biochimie 2014, 100, 107–120. [Google Scholar] [CrossRef]
- Flood, P.J.; Theeuwen, T.P.J.M.; Schneeberger, K.; Keizer, P.; Kruijer, W.; Severing, E.; Kouklas, E.; Hageman, J.A.; Wijfjes, R.; Calvo-Baltanas, V.; et al. Reciprocal Cybrids Reveal How Organellar Genomes Affect Plant Phenotypes. Nat. Plants 2020, 6, 13–21. [Google Scholar] [CrossRef]
- Maréchal, A.; Brisson, N. Recombination and the Maintenance of Plant Organelle Genome Stability. New Phytol. 2010, 186, 299–317. [Google Scholar] [CrossRef]
- Ellison, C.K.; Burton, R.S. Cytonuclear Conflict in Interpopulation Hybrids: The Role of RNA Polymerase in MtDNA Transcription and Replication. J. Evol. Biol. 2010, 23, 528–538. [Google Scholar] [CrossRef]
- McCauley, D.E. Paternal Leakage, Heteroplasmy, and the Evolution of Plant Mitochondrial Genomes. New Phytol. 2013, 200, 966–977. [Google Scholar] [CrossRef]
- Christov, M. Contribution of Interspecific Hybridization to Sunflower Breeding. Helia 2012, 35, 37–46. [Google Scholar] [CrossRef]
- Christov, M. Contribution of interspecific and intergeneric hybridization to sunflower breeding. Helia 2013, 36, 1–18. [Google Scholar] [CrossRef]
- Horn, R. Molecular Diversity of Male Sterility Inducing and Male-Fertile Cytoplasms in the Genus Helianthus. Appl. Genet. 2002, 104, 562–570. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-Metagenomes from Highly Chimeric Reads. In Research in Computational Molecular Biology; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 158–170. [Google Scholar]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive Visualization of de Novo Genome Assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using Tablet for Visual Exploration of Second-Generation Sequencing Data. Brief. Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and Accurate Annotation of Organelle Genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Lenz, H.; Hein, A.; Knoop, V. Plant Organelle RNA Editing and Its Specificity Factors: Enhancements of Analyses and New Database Features in PREPACT 3.0. BMC Bioinform. 2018, 19, 255. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Wu, S.; Liu, W.; Aljohi, H.A.; Alromaih, S.A.; Alanazi, I.O.; Lin, Q.; Yu, J.; Hu, S. REDO: RNA Editing Detection in Plant Organelles Based on Variant Calling Results. J. Comput. Biol. 2018, 25, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Makarenko, M.; Usatov, A.; Tatarinova, T.; Azarin, K.; Kovalevich, A.; Gavrilova, V.; Horn, R. The Investigation of Perennial Sunflower Species (Helianthus L.) Mitochondrial Genomes. Genes 2020, 11, 982. [Google Scholar] [CrossRef] [PubMed]
- Makarenko, M.S.; Omelchenko, D.O.; Usatov, A.V.; Gavrilova, V.A. The Insights into Mitochondrial Genomes of Sunflowers. Plants 2021, 10, 1774. [Google Scholar] [CrossRef]
- Kovar, L.; Nageswara-Rao, M.; Ortega-Rodriguez, S.; Dugas, D.V.; Straub, S.; Cronn, R.; Strickler, S.R.; Hughes, C.E.; Hanley, K.A.; Rodriguez, D.N.; et al. PacBio-Based Mitochondrial Genome Assembly of Leucaena Trichandra (Leguminosae) and an Intrageneric Assessment of Mitochondrial RNA Editing. Genome Biol. Evol. 2018, 10, 2501–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szandar, K.; Krawczyk, K.; Myszczyński, K.; Ślipiko, M.; Sawicki, J.; Szczecińska, M. Breaking the Limits—Multichromosomal Structure of an Early Eudicot Pulsatilla Patens Mitogenome Reveals Extensive RNA-Editing, Longest Repeats and Chloroplast Derived Regions among Sequenced Land Plant Mitogenomes. BMC Plant Biol. 2022, 22, 109. [Google Scholar] [CrossRef] [PubMed]
- Small, I.D.; Schallenberg-Rüdinger, M.; Takenaka, M.; Mireau, H.; Ostersetzer-Biran, O. Plant Organellar RNA Editing: What 30 Years of Research Has Revealed. Plant J. 2020, 101, 1040–1056. [Google Scholar] [CrossRef]
- Wynn, E.L.; Christensen, A.C. Repeats of Unusual Size in Plant Mitochondrial Genomes: Identification, Incidence and Evolution. G3 Genes Genomes Genet. 2019, 9, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Owens, G.L.; Huang, K.; Todesco, M.; Rieseberg, L.H. Re-Evaluating Homoploid Reticulate Evolution in Helianthus Sunflowers. Mol. Biol. Evol. 2023, 40, msad013. [Google Scholar] [CrossRef]
- Zhang, J.-Q.; Imerovski, I.; Borkowski, K.; Huang, K.; Burge, D.; Rieseberg, L.H. Intraspecific Genetic Divergence within Helianthus niveus and the Status of Two New Morphotypes from Mexico. Am. J. Bot. 2019, 106, 1229–1239. [Google Scholar] [CrossRef]
- Sharma, M.; Gavisiddaiah, S.Y.; Rao, A.M.; Ramesh, S. Utilization of Wild Species for Diversifying the Cytoplasmic Male Sterility Source of Sunflower (Helianthus annuus L.) Hybrids. Helia 2022, 45, 71–98. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makarenko, M.S.; Azarin, K.V.; Gavrilova, V.A. Mitogenomic Research of Silverleaf Sunflower (Helianthus argophyllus) and Its Interspecific Hybrids. Curr. Issues Mol. Biol. 2023, 45, 4841-4849. https://doi.org/10.3390/cimb45060308
Makarenko MS, Azarin KV, Gavrilova VA. Mitogenomic Research of Silverleaf Sunflower (Helianthus argophyllus) and Its Interspecific Hybrids. Current Issues in Molecular Biology. 2023; 45(6):4841-4849. https://doi.org/10.3390/cimb45060308
Chicago/Turabian StyleMakarenko, Maksim S., Kirill V. Azarin, and Vera A. Gavrilova. 2023. "Mitogenomic Research of Silverleaf Sunflower (Helianthus argophyllus) and Its Interspecific Hybrids" Current Issues in Molecular Biology 45, no. 6: 4841-4849. https://doi.org/10.3390/cimb45060308
APA StyleMakarenko, M. S., Azarin, K. V., & Gavrilova, V. A. (2023). Mitogenomic Research of Silverleaf Sunflower (Helianthus argophyllus) and Its Interspecific Hybrids. Current Issues in Molecular Biology, 45(6), 4841-4849. https://doi.org/10.3390/cimb45060308