
Adenovirus as a Vector and Oncolytic Virus


Abstract
:1. Introduction
2. Trends of Viral Vectors
2.1. Retroviral Vectors
2.2. Lentiviral Vectors
2.3. Adeno-Associated Virus (AAV) Vectors
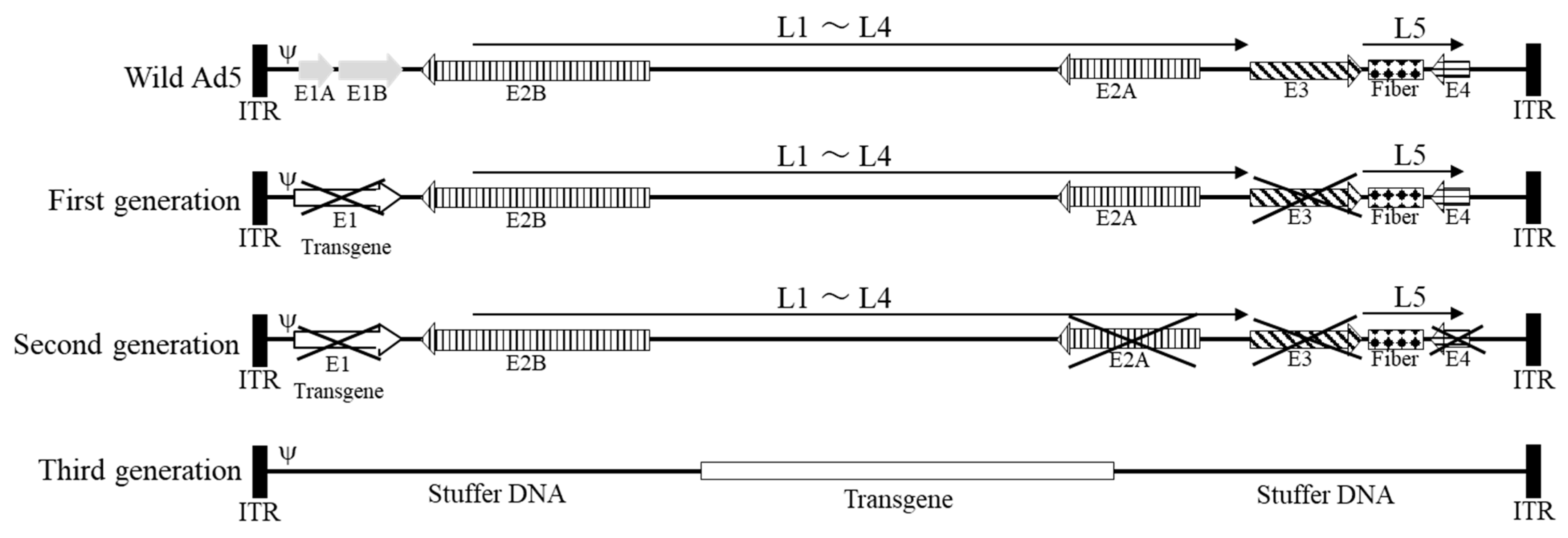
3. Adenovirus as a Vector
3.1. First-Generation Adenoviral Vectors
3.2. Second-Generation Adenoviral Vectors
3.3. Third-Generation Adenoviral Vectors
3.4. Current Status of Adenoviral Vectors
4. Adenovirus as an Oncolytic Virus
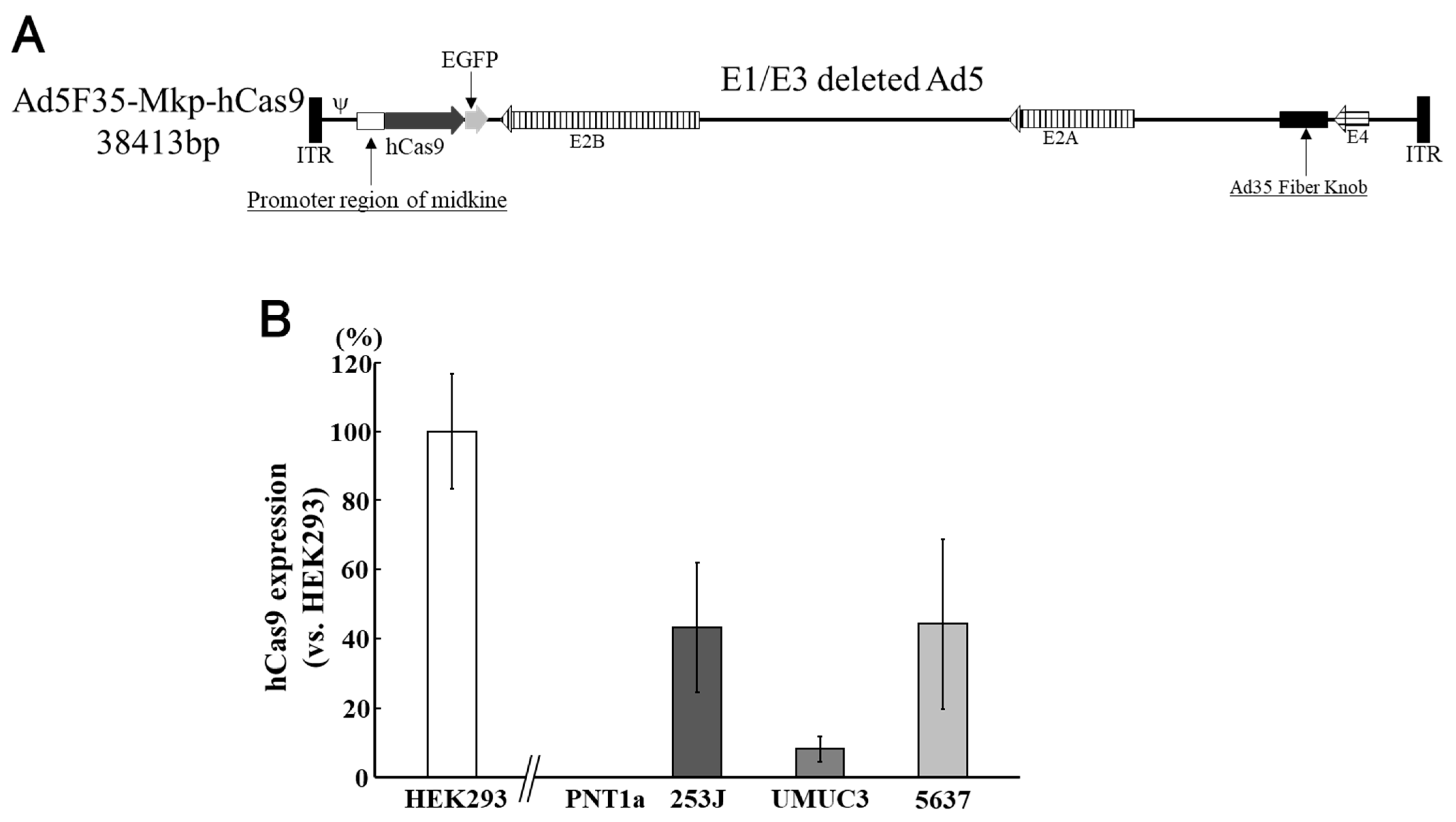
5. Use of Adenoviruses for Bladder Cancer

6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhingra, A.; Hage, E.; Ganzenmueller, T.; Böttcher, S.; Hofmann, J.; Hamprecht, K.; Obermeier, P.; Rath, B.; Hausmann, F.; Dobner, T.; et al. Molecular Evolution of Human Adenovirus (HAdV) Species C. Sci. Rep. 2019, 9, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akli, S.; Caillaud, C.; Vigne, E.; Stratford-Perricaudet, L.D.; Poenaru, L.; Perricaudet, M.; Kahn, A.; Peschanski, M.R. Transfer of a foreign gene into the brain using adenovirus vectors. Nat. Genet. 1993, 3, 224–228. [Google Scholar] [CrossRef]
- Gene Therapy Clinical Trials Worldwide. 2022. Available online: https://a873679.fmphost.com/fmi/webd/GTCT (accessed on 28 February 2023).
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Ginn, S.L.; Alexander, I.E.; Edelstein, M.L.; Abedi, M.R.; Wixon, J. Gene therapy clinical trials worldwide to 2012—An update. J. Gene Med. 2013, 15, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; Garrigue, A.; Wang, G.P.; Soulier, J.; Lim, A.; Morillon, E.; Clappier, E.; Caccavelli, L.; Delabesse, E.; Beldjord, K.; et al. Insertional oncogenesis in 4 patients after retrovirus-mediated gene therapy of SCID-X1. J. Clin. Investig. 2008, 118, 3132–3142. [Google Scholar] [CrossRef]
- Howe, S.J.; Mansour, M.R.; Schwarzwaelder, K.; Bartholomae, C.; Hubank, M.; Kempski, H.; Brugman, M.H.; Pike-Overzet, K.; Chatters, S.J.; de Ridder, D.; et al. Insertional mutagenesis combined with acquired somatic mutations causes leukemogenesis following gene therapy of SCID-X1 patients. J. Clin. Investig. 2008, 118, 3143–3150. [Google Scholar] [CrossRef]
- Labbé, R.P.; Vessillier, S.; Rafiq, Q.A. Lentiviral Vectors for T Cell Engineering: Clinical Applications, Bioprocessing and Future Perspectives. Viruses 2021, 13, 1528. [Google Scholar] [CrossRef]
- Mori, S.; Wang, L.; Takeuchi, T.; Kanda, T. Two novel adeno-associated viruses from cynomolgus monkey: Pseudotyping characterization of capsid protein. Virology 2004, 330, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, T.K.; Samulski, R.J. Addressing high dose AAV toxicity—‘One and done’ or ‘slower and lower’? Expert Opin. Biol. Ther. 2022, 22, 1067–1071. [Google Scholar] [CrossRef]
- High, K.A.; Anguela, X.M. Adeno-associated viral vectors for the treatment of hemophilia. Hum. Mol. Genet. 2016, 25, R36–R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo-Serrano, A.; Rico, A.J.; Roda, E.; Honrubia, A.; Arrieta, S.; Ariznabarreta, G.; Chocarro, J.; Lorenzo-Ramos, E.; Pejenaute, A.; Vázquez, A.; et al. Adeno-Associated Viral Vectors as Versatile Tools for Parkinson’s Research, Both for Disease Modeling Purposes and for Therapeutic Uses. Int. J. Mol. Sci. 2021, 22, 6389. [Google Scholar] [CrossRef] [PubMed]
- Reul, J.; Frisch, J.; Engeland, C.E.; Thalheimer, F.B.; Hartmann, J.; Ungerechts, G.; Buchholz, C.J. Tumor-Specific Delivery of Immune Checkpoint Inhibitors by Engineered AAV Vectors. Front. Oncol. 2019, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA Approves Innovative Gene Therapy to Treat Pediatric Patients with Spinal Muscular Atrophy, A Rare Disease and Leading Genetic Cause of Infant Mortality. 24 May 2019. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-innovative-gene-therapy-treat-pediatric-patients-spinal-muscular-atrophy-rare-disease (accessed on 19 January 2023).
- Day, J.W.; Finkel, R.S.; Chiriboga, C.A.; Connolly, A.M.; Crawford, T.O.; Darras, B.T.; Iannaccone, S.T.; Kuntz, N.L.; Peña, L.D.M.; Shieh, P.B.; et al. Onasemnogene abeparvovec gene therapy for symptomatic infantile-onset spinal muscular atrophy in patients with two copies of SMN2 (STR1VE): An open-label, single-arm, multicentre, phase 3 trial. Lancet Neurol. 2021, 20, 284–293. [Google Scholar] [CrossRef] [PubMed]
- King, C.R.; Zhang, A.; Tessier, T.M.; Gameiro, S.F.; Mymryk, J.S. Hacking the Cell: Network Intrusion and Exploitation by Adenovirus E1A. mBio 2018, 9, e00390-18. [Google Scholar] [CrossRef] [Green Version]
- Louis, N.; Evelegh, C.; Graham, F.L. Cloning and sequencing of the cellular-viral junctions from the human adenovirus type 5 transformed 293 cell line. Virology 1997, 233, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, E.R.A.; Bouvier, M. Immune evasion by adenoviruses: A window into host-virus adaptation. FEBS Lett. 2019, 593, 3496–3503. [Google Scholar] [CrossRef] [Green Version]
- Imperiale, M.J.; Kao, H.T.; Feldman, L.T.; Nevins, J.R.; Strickland, S. Common control of the heat shock gene and early adenovirus genes: Evidence for a cellular E1A-like activity. Mol. Cell Biol. 1984, 4, 867–874. [Google Scholar]
- Hartman, Z.C.; Appledorn, D.M.; Amalfitano, A. Adenovirus vector induced innate immune responses: Impact upon efficacy and toxicity in gene therapy and vaccine applications. Virus Res. 2008, 132, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Parks, R.J. Human adenovirus type 5 vectors deleted of early region 1 (E1) undergo limited expression of early replicative E2 proteins and DNA replication in non-permissive cells. PLoS ONE 2017, 12, e0181012. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Hearing, P. Distinct roles of the adenovirus E4 ORF3 protein in viral DNA replication and inhibition of genome concatenation. J. Virol. 2003, 77, 5295–5304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neal, W.K.; Zhou, H.; Morral, N.; Aguilar-Cordova, E.; Pestaner, J.; Langston, C.; Mull, B.; Wang, Y.; Beaudet, A.L.; Lee, B. Toxicological comparison of E2a-deleted and first-generation adenoviral vectors expressing alpha1-antitrypsin after systemic delivery. Hum. Gene Ther. 1998, 9, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Brunetti-Pierri, N.; Ng, P. Progress and prospects: Gene therapy for genetic diseases with helper-dependent adenoviral vectors. Gene Ther. 2008, 15, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.; Beauchamp, C.; Evelegh, C.; Parks, R.; Graham, F.L. Development of a FLP/frt system for generating helper-dependent adenoviral vectors. Mol. Ther. 2001, 3, 809–815. [Google Scholar] [CrossRef]
- Jozkowicz, A.; Dulak, J.; Nanobashvili, J.; Polterauer, P.; Prager, M.; Huk, I. Gutless adenoviral vectors—Promising tools for gene therapy. Eur. Surg. 2002, 34, 95–100. [Google Scholar] [CrossRef]
- Liu, J.; Seol, D.W. Helper virus-free gutless adenovirus (HF-GLAd): A new platform for gene therapy. BMB Rep. 2020, 53, 565–575. [Google Scholar] [CrossRef]
- Lee, D.; Liu, J.; Junn, H.J.; Lee, E.J.; Jeong, K.S.; Seol, D.W. No more helper adenovirus: Production of gutless adenovirus (GLAd) free of adenovirus and replication-competent adenovirus (RCA) contaminants. Exp. Mol. Med. 2019, 51, 1–18. [Google Scholar] [CrossRef]
- Parks, R.J.; Graham, F.L. A helper-dependent system for adenovirus vector production helps define a lower limit for efficient DNA packaging. J. Virol. 1997, 71, 3293–3329. [Google Scholar] [CrossRef] [Green Version]
- Parks, R.J.; Bramson, J.L.; Wan, Y.; Addison, C.L.; Graham, F.L. Effects of stuffer DNA on transgene expression from helper-dependent adenovirus vectors. J. Virol. 1999, 73, 8027–8034. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.J.; Kennedy, M.A.; Parks, R.J. Host cell detection of noncoding stuffer DNA contained in helper-dependent adenovirus vectors leads to epigenetic repression of transgene expression. J. Virol. 2009, 83, 8409–8417. [Google Scholar] [CrossRef] [Green Version]
- Schiedner, G.; Hertel, S.; Johnston, M.; Biermann, V.; Dries, V.; Kochanek, S. Variables affecting in vivo performance of high-capacity adenovirus vectors. J. Virol. 2002, 76, 1600–1609. [Google Scholar] [CrossRef] [Green Version]
- Boorjian, S.A.; Alemozaffar, M.; Konety, B.R.; Shore, N.D.; Gomella, L.G.; Kamat, A.M.; Bivalacqua, T.J.; Montgomery, J.S.; Lerner, S.P.; Busby, J.E.; et al. Intravesical nadofaragene firadenovec gene therapy for BCG-unresponsive non-muscle-invasive bladder cancer: A single-arm, open-label, repeat-dose clinical trial. Lancet Oncol. 2021, 1, 107–117. [Google Scholar] [CrossRef]
- Green, J.L.; Osterhout, R.E.; Klova, A.L.; Merkwirth, C.; McDonnell, S.R.P.; Zavareh, R.B.; Fuchs, B.C.; Kamal, A.; Jakobsen, J.S. Molecular characterization of type I IFN-induced cytotoxicity in bladder cancer cells reveals biomarkers of resistance. Mol. Ther. Oncolytics 2021, 23, 547–559. [Google Scholar] [CrossRef]
- FDA Approves First Gene Therapy for the Treatment of High-Risk, Non-Muscle-Invasive Bladder Cancer. (6 December 2022). Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-gene-therapy-treatment-high-risk-non-muscle-invasive-bladder-cancer (accessed on 22 May 2023).
- Zhu, R.; Weng, D.; Lu, S.; Lin, D.; Wang, M.; Chen, D.; Lv, J.; Li, H.; Lv, F.; Xi, L.; et al. Double-Dose Adenovirus-Mediated Adjuvant Gene Therapy Improves Liver Transplantation Outcomes in Patients with Advanced Hepatocellular Carcinoma. Hum. Gene Ther. 2018, 29, 251–258. [Google Scholar] [CrossRef]
- Kuruppu, D.; Tanabe, K.K. Viral oncolysis by herpes simplex virus and other viruses. Cancer Biol. Ther. 2005, 4, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Kelly, E.; Russell, S.J. History of oncolytic viruses: Genesis to genetic engineering. Mol. Ther. 2007, 15, 651–659. [Google Scholar] [CrossRef]
- Eto, Y.; Nagai, K. Oncolytic Viruses as Cancer Therapeutic agents Current Status of Oncolytic Viruses. J. Kyushu Univ. Health Welf. 2017, 18, 49–53. (In Japanese) [Google Scholar]
- Cao, G.D.; He, X.B.; Sun, Q.; Chen, S.; Wan, K.; Xu, X.; Feng, X.; Li, P.P.; Chen, B.; Xiong, M.M. The Oncolytic Virus in Cancer Diagnosis and Treatment. Front. Oncol. 2020, 10, 1786. [Google Scholar] [CrossRef]
- Heise, C.; Sampson-Johannes, A.; Williams, A.; McCormick, F.; Von Hoff, D.D.; Kirn, D.H. ONYX-015, an E1B gene-attenuated adenovirus, causes tumor-specific cytolysis and antitumoral efficacy that can be augmented by standard chemotherapeutic agents. Nat. Med. 1997, 3, 639–645. [Google Scholar] [CrossRef]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef]
- Nemunaitis, J.; Ganly, I.; Khuri, F.; Arseneau, J.; Kuhn, J.; McCarty, T.; Landers, S.; Maples, P.; Romel, L.; Randlev, B.; et al. Selective replication and oncolysis in p53 mutant tumors with ONYX-015, an E1B-55kD gene-deleted adenovirus, in patients with advanced head and neck cancer: A phase II trial. Cancer Res. 2000, 60, 6359–6366. [Google Scholar]
- Garber, K. China approves world’s first oncolytic virus therapy for cancer treatment. J. Nat. Cancer Inst. 2006, 98, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Babiker, H.M.; Riaz, I.B.; Husnain, M.; Borad, M.J. Oncolytic virotherapy including Rigvir and standard therapies in malignant melanoma. Oncolytic Virother. 2017, 6, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef]
- Press Release from Daiichi-Sankyo. 11 June 2021. Available online: https://www.daiichisankyo.co.jp/files/news/pressrelease/pdf/202106/20210611_J.pdf (accessed on 12 November 2022).
- Esaki, S.; Goshima, F.; Ozaki, H.; Takano, G.; Hatano, Y.; Kawakita, D.; Ijichi, K.; Watanabe, T.; Sato, Y.; Murata, T.; et al. Oncolytic activity of HF10 in head and neck squamous cell carcinomas. Cancer Gene Ther. 2020, 27, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, T.; Kagawa, S.; Kobayashi, N.; Shirakiya, Y.; Umeoka, T.; Teraishi, F.; Taki, M.; Kyo, S.; Tanaka, N.; Fujiwara, T. Telomerase-specific replication-selective virotherapy for human cancer. Clin. Cancer Res. 2004, 10, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Gohara, S.; Shinohara, K.; Yoshida, R.; Kariya, R.; Tazawa, H.; Hashimoto, M.; Inoue, J.; Kubo, R.; Nakashima, H.; Arita, H.; et al. An oncolytic virus as a promising candidate for the treatment of radioresistant oral squamous cell carcinoma. Mol. Ther. Oncolytics 2022, 27, 141–156. [Google Scholar] [CrossRef]
- Phase 2 Study of OBP-301 (Telomelysin™) in Combination with Pembrolizumab and SBRT in Patients with HNSCC with Inoperable, Recurrent or Progressive Disease. 28 December 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04685499 (accessed on 23 December 2022).
- Packiam, V.T.; Lamm, D.L.; Barocas, D.A.; Trainer, A.; Fand, B.; Davis, R.L., 3rd; Clark, W.; Kroeger, M.; Dumbadze, I.; Chamie, K.; et al. An open label, single-arm, phase II multicenter study of the safety and efficacy of CG0070 oncolytic vector regimen in patients with BCG-unresponsive non-muscle-invasive bladder cancer: Interim results. Urol. Oncol. 2018, 36, 440–447. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Li, L.; Wu, J.; Zhang, H.; Zhang, H.; Lei, T.; Xu, B. Oncolytic Adenovirus: Prospects for Cancer Immunotherapy. Front. Microbiol. 2021, 12, 707290. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
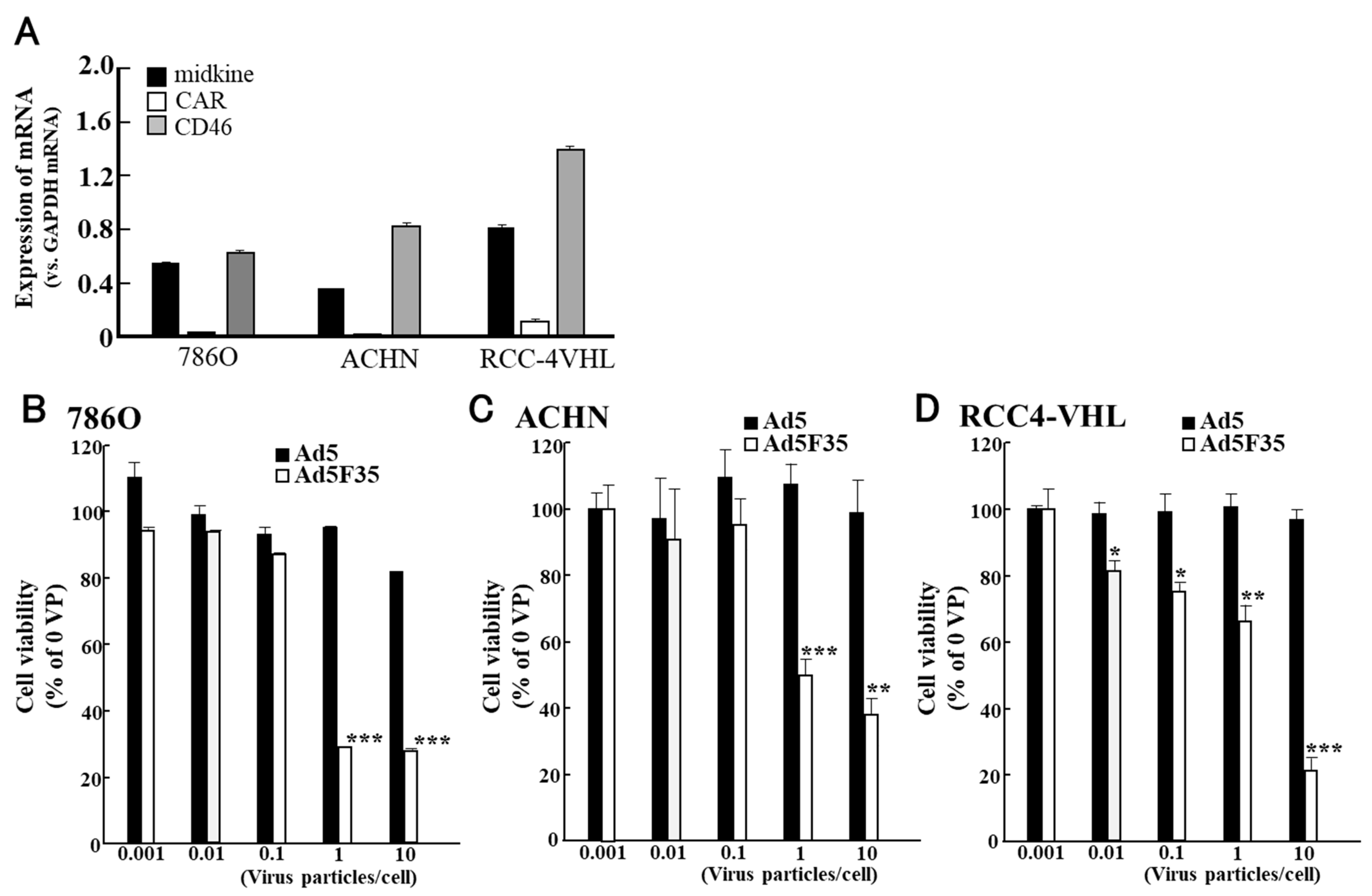
- Okegawa, T.; Sayne, J.R.; Nutahara, K.; Pong, R.C.; Saboorian, H.; Kabbani, W.; Higashihara, E.; Hsieh, J.T. A histone deacetylase inhibitor enhances adenoviral infection of renal cancer cells. J. Urol. 2007, 177, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
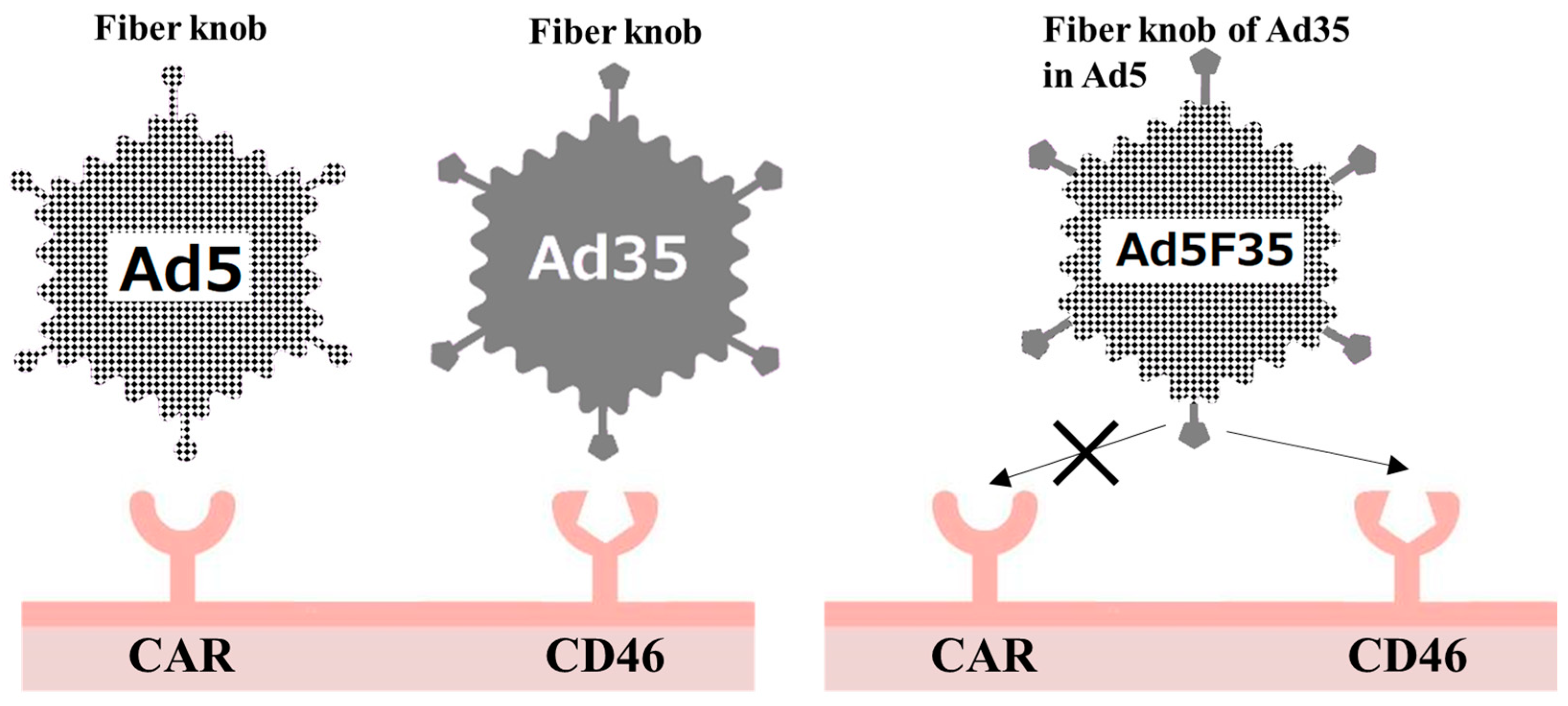
- Hensen, L.C.M.; Hoeben, R.C.; Bots, S.T.F. Adenovirus Receptor Expression in Cancer and Its Multifaceted Role in Oncolytic Adenovirus Therapy. Int. J. Mol. Sci. 2020, 21, 6828. [Google Scholar] [CrossRef]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Segerman, A.; Atkinson, J.P.; Marttila, M.; Dennerquist, V.; Wadell, G.; Arnberg, N. Adenovirus type 11 uses CD46 as a cellular receptor. J. Virol. 2003, 77, 9183–9191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuguchi, H.; Hayakawa, T. Adenovirus vectors containing chimeric type 5 and type 35 fiber proteins exhibit altered and expanded tropism and increase the size limit of foreign genes. Gene 2002, 285, 69–77. [Google Scholar] [CrossRef]
- Shayakhmetov, D.M.; Papayannopoulou, T.; Stamatoyannopoulos, G.; Lieber, A. Efficient gene transfer into human CD34(+) cells by a retargeted adenovirus vector. J. Virol. 2000, 74, 2567–2583. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, E.; Doi, R.; Kami, K.; Mori, T.; Ito, D.; Koizumi, M.; Kida, A.; Nagai, K.; Ito, T.; Masui, T.; et al. Adenovirus vectors with chimeric type 5 and 35 fiber proteins exhibit enhanced transfection of human pancreatic cancer cells. Int. J. Oncol. 2008, 33, 1141–1147. [Google Scholar]
- Yu, L.; Shimozato, O.; Li, Q.; Kawamura, K.; Ma, G.; Namba, M.; Ogawa, T.; Kaiho, I.; Tagawa, M. Adenovirus type 5 substituted with type 11 or 35 fiber structure increases its infectivity to human cells enabling dual gene transfer in CD46-dependent and independent manners. Anticancer Res. 2007, 27, 2311–2316. [Google Scholar]
- Ni, S.; Gaggar, A.; Di Paolo, N.; Li, Z.Y.; Liu, Y.; Strauss, R.; Sova, P.; Morihara, J.; Feng, Q.; Kiviat, N.; et al. Evaluation of adenovirus vectors containing serotype 35 fibers for tumor targeting. Cancer Gene Ther. 2006, 13, 1072–1081. [Google Scholar] [CrossRef]
- Ono, R.; Takayama, K.; Sakurai, F.; Mizuguchi, H. Efficient antitumor effects of a novel oncolytic adenovirus fully composed of species B adenovirus serotype 35. Mol. Ther. Oncolytics 2021, 20, 399–409. [Google Scholar] [CrossRef]
- Hemminki, O.; Diaconu, I.; Cerullo, V.; Pesonen, S.K.; Kanerva, A.; Joensuu, T.; Kairemo, K.; Laasonen, L.; Partanen, K.; Kangasniemi, L.; et al. Ad3-hTERT-E1A, a fully serotype 3 oncolytic adenovirus, in patients with chemotherapy refractory cancer. Mol. Ther. 2012, 20, 1821–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauerschmitz, G.J.; Guse, K.; Kanerva, A.; Menzel, A.; Herrmann, I.; Desmond, R.A.; Yamamoto, M.; Nettelbeck, D.M.; Hakkarainen, T.; Dall, P.; et al. Triple-targeted oncolytic adenoviruses featuring the cox2 promoter, E1A transcomplementation, and serotype chimerism for enhanced selectivity for ovarian cancer cells. Mol. Ther. 2006, 14, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhou, Y.; Wu, H.; Wang, Z.; Zhan, Y.; Feng, X.; Geng, R.; Wu, Y.; Kong, W.; Yu, X. Seroprevalence of neutralizing antibodies to human adenovirus type 5 in healthy adults in China. J. Med. Virol. 2012, 84, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Havunen, R.; Siurala, M.; Sorsa, S.; Grönberg-Vähä-Koskela, S.; Behr, M.; Tähtinen, S.; Santos, J.M.; Karell, P.; Rusanen, J.; Nettelbeck, D.M.; et al. Oncolytic Adenoviruses Armed with Tumor Necrosis Factor Alpha and Interleukin-2 Enable Successful Adoptive Cell Therapy. Mol. Ther. Oncolyt. 2017, 4, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Heiniö, C.; Cervera-Carrascon, V.; Quixabeira, D.C.A.; Siurala, M.; Havunen, R.; Butzow, R.; Zafar, S.; de Gruijl, T.; Lassus, H.; et al. Oncolytic adenovirus shapes the ovarian tumor microenvironment for potent tumor-infiltrating lymphocyte tumor reactivity. J. Immunother. Cancer 2020, 8, 000188. [Google Scholar] [CrossRef] [Green Version]
- Kuryk, L.; Haavisto, E.; Garofalo, M.; Capasso, C.; Hirvinen, M.; Pesonen, S.; Ranki, T.; Vassilev, L.; Cerullo, V. Synergistic anti-tumor efficacy of immunogenic adenovirus ONCOS-102 (Ad5/3-D24-GM-CSF) and standard of care chemotherapy in preclinical mesothelioma model. Int. J. Cancer 2016, 139, 1883–1893. [Google Scholar] [CrossRef] [Green Version]
- Barton, K.N.; Paielli, D.; Zhang, Y.; Koul, S.; Brown, S.L.; Lu, M.; Seely, J.; Kim, J.H.; Freytag, S.O. Second-generation replication-competent oncolytic adenovirus armed with improved suicide genes and ADP gene demonstrates greater efficacy without increased toxicity. Mol. Ther. 2006, 13, 347–356. [Google Scholar] [CrossRef]
- Nyati, M.K.; Symon, Z.; Kievit, E.; Dornfeld, K.J.; Rynkiewicz, S.D.; Ross, B.D.; Rehemtulla, A.; Lawrence, T.S. The potential of 5-fluorocytosine/cytosine deaminase enzyme prodrug gene therapy in an intrahepatic colon cancer model. Gene Ther. 2002, 9, 844–849. [Google Scholar] [CrossRef]
- Moolten, F.L. Tumor chemosensitivity conferred by inserted herpes thymidine kinase genes: Paradigm for a prospective cancer control strategy. Cancer Res. 1986, 46, 5276–5281. [Google Scholar]
- Leopardo, D.; Cecere, S.C.; Di Napoli, M.; Cavaliere, C.; Pisano, C.; Striano, S.; Marra, L.; Menna, L.; Claudio, L.; Perdonà, S.; et al. Intravesical chemo-immunotherapy in non-muscle invasive bladder cancer. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2145–2158. [Google Scholar]
- Herr, H.W.; Morales, A. History of bacillus Calmette-Guerin and bladder cancer: An immunotherapy success story. J. Urol. 2008, 179, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Lin, W.Y.; Chang, Y.C.; Huang, C.H.; Tzeng, H.E.; Abdul-Lattif, E.; Wang, T.H.; Tseng, T.H.; Kang, Y.N.; Chi, K.Y. The Association between Baseline Proton Pump Inhibitors, Immune Checkpoint Inhibitors, and Chemotherapy: A Systematic Review with Network Meta-Analysis. Cancers 2022, 15, 284. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Hughes, M.; Kammula, U.; Royal, R.; Sherry, R.M.; Topalian, S.L.; Suri, K.B.; Levy, C.; Allen, T.; Mavroukakis, S.; et al. Ipilimumab (anti-CTLA4 antibody) causes regression of metastatic renal cell cancer associated with enteritis and hypophysitis. J. Immunother. 2007, 30, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Guse, K.; Cerullo, V.; Hemminki, A. Oncolytic vaccinia virus for the treatment of cancer. Expert Opin. Biol. Ther. 2011, 11, 595–608. [Google Scholar] [CrossRef]
- Mantwill, K.; Klein, F.G.; Wang, D.; Hindupur, S.V.; Ehrenfeld, M.; Holm, P.S.; Nawroth, R. Concepts in Oncolytic Adenovirus Therapy. Int. J. Mol. Sci. 2021, 22, 10522. [Google Scholar] [CrossRef]
- Kadomatsu, K.; Huang, R.P.; Suganuma, T.; Murata, F.; Muramatsu, T. A retinoic acid responsive gene MK found in the teratocarcinoma system is expressed in spatially and temporally controlled manner during mouse embryogenesis. J. Cell Biol. 1990, 110, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Choudhuri, R.; Zhang, H.T.; Donnini, S.; Ziche, M.; Bicknell, R. An angiogenic role for the neurokines midkine and pleiotrophin in tumorigenesis. Cancer Res. 1997, 57, 1814–1819. [Google Scholar]
- Kadomatsu, K.; Hagihara, M.; Akhter, S.; Fan, Q.W.; Muramatsu, H.; Muramatsu, T. Midkine induces the transformation of NIH3T3 cells. Br. J. Cancer 1997, 75, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, T. Structure and function of midkine as the basis of its pharmacological effects. Br. J. Pharmacol. 2014, 171, 814–826. [Google Scholar] [CrossRef] [Green Version]
- Mashour, G.A.; Ratner, N.; Khan, G.A.; Wang, H.L.; Martuza, R.L.; Kurtz, A. The angiogenic factor midkine is aberrantly expressed in NF1-deficient Schwann cells and is a mitogen for neurofibroma-derived cells. Oncogene 2001, 20, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Kwiatkowski, D.J.; Henske, E.P. Midkine expression by stem-like tumor cells drives persistence to mTOR inhibition and an immune-suppressive microenvironment. Nat. Commun. 2022, 13, 5018. [Google Scholar] [CrossRef]
- Ikematsu, S.; Yano, A.; Aridome, K.; Kikuchi, M.; Kumai, H.; Nagano, H.; Okamoto, K.; Oda, M.; Sakuma, S.; Aikou, T.; et al. Serum midkine levels are increased in patients with various types of carcinomas. Br. J. Cancer 2000, 83, 701–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Maeta, H.; Kato, S.; Shinozawa, T.; Terada, T. Immunohistochemical and in situ hybridization analyses of midkine expression in thyroid papillary carcinoma. Mod. Pathol. 2000, 13, 1060–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowhari Shabgah, A.; Ezzatifar, F.; Aravindhan, S.; Olegovna Zekiy, A.; Ahmadi, M.; Gheibihayat, S.M.; Gholizadeh Navashenaq, J. Shedding more light on the role of Midkine in hepatocellular carcinoma: New perspectives on diagnosis and therapy. IUBMB Life 2021, 73, 659–669. [Google Scholar] [CrossRef]
- Uehara, K.; Matsubara, S.; Kadomatsu, K.; Tsutsui, J.; Muramatsu, T. Genomic structure of human midkine (MK), a retinoic acid-responsive growth/differentiation factor. J. Biochem. 1992, 111, 563–567. [Google Scholar] [CrossRef]
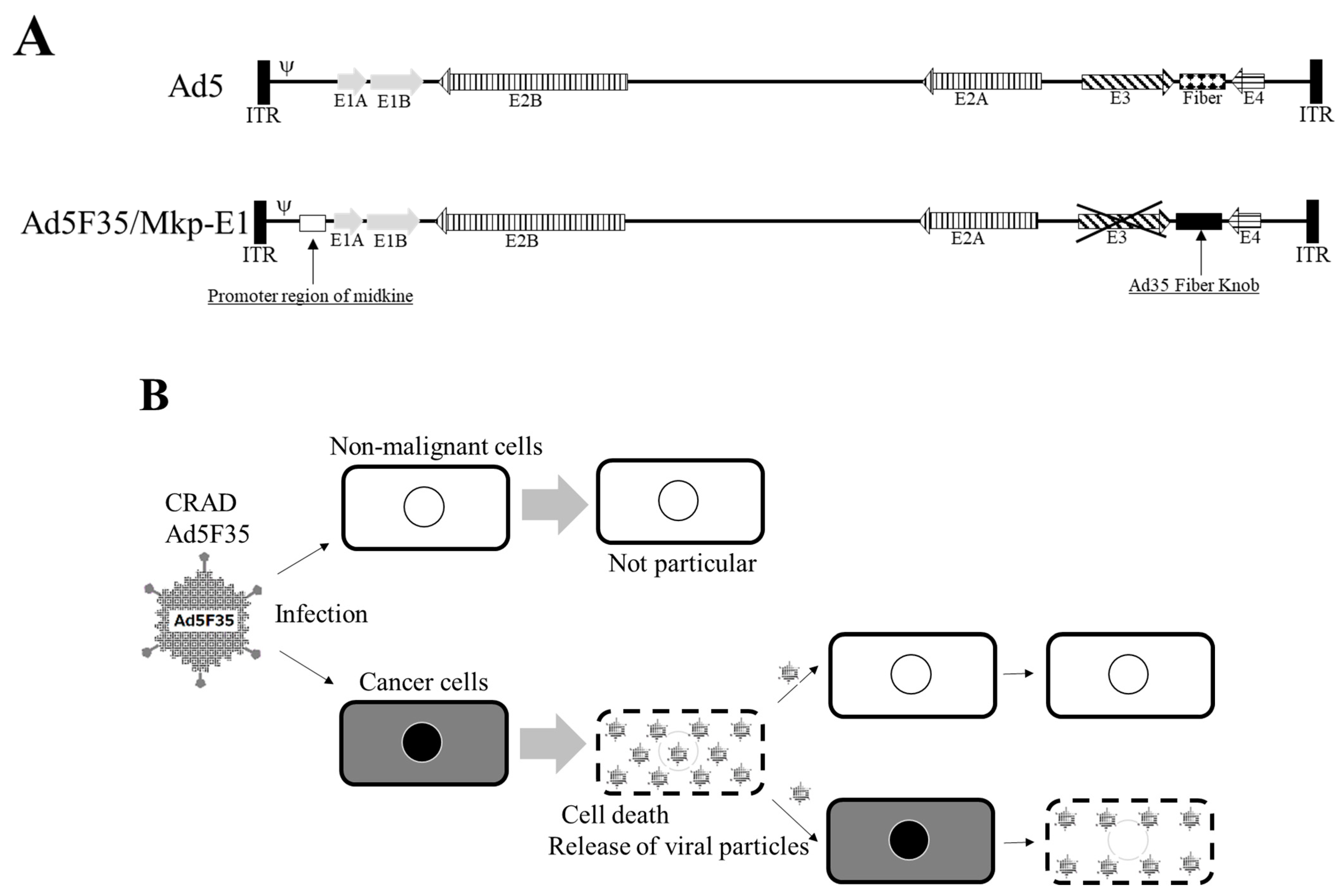
- Gotoh, A.; Nagaya, H.; Kanno, T.; Tagawa, M.; Nishizaki, T. Fiber-substituted conditionally replicating adenovirus Ad5F35 induces oncolysis of human bladder cancer cells in in vitro analysis. Urology 2013, 81, 920.e7–920.e11. [Google Scholar] [CrossRef]
- Kanno, T.; Gotoh, A.; Nakano, T.; Tagawa, M.; Nishizaki, T. Beneficial oncolytic effect of fiber-substituted conditionally replicating adenovirus on human lung cancer. Anticancer Res. 2012, 32, 4891–4895. [Google Scholar]
- Gotoh, A.; Kanno, T.; Nagaya, H.; Nakano, T.; Tabata, C.; Fukuoka, K.; Tagawa, M.; Nishizaki, T. Gene therapy using adenovirus against malignant mesothelioma. Anticancer Res. 2012, 32, 3743–3747. [Google Scholar]
- Nagaya, H.; Tagawa, M.; Hiwasa, K.; Terao, S.; Kanno, T.; Nishizaki, T.; Gotoh, A. Fiber-substituted conditionally replicating adenovirus for oncolysis of human renal carcinoma cells. Anticancer Res. 2012, 32, 2985–2989. [Google Scholar]
- Yao, S.; He, Z.; Chen, C. CRISPR/Cas9-mediated genome editing of epigenetic factors for cancer therapy. Hum. Gene Ther. 2015, 26, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Akram, F.; Ul Haq, I.; Ahmed, Z.; Khan, H.; Ali, M.S. CRISPR-Cas9, a promising therapeutic tool for cancer therapy: A review. Protein Pept. Lett. 2020, 27, 931–944. [Google Scholar] [PubMed]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunaga, W.; Hamada, K.; Tagawa, M.; Morinaga, T.; Gotoh, A. Cancer Cell-specific Transfection of hCas9 Gene Using Ad5F35 Vector. Anticancer Res. 2021, 41, 3731–3740. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Share in Clinical Trials until 2012 [5] | Share in Clinical Trials until 2017 [6] | Share in Clinical Trials until 2022 [3] | Increase 2012 to 17 | Increase 2017 to 22 | ||||
|---|---|---|---|---|---|---|---|---|
| Adenovirus | 23.3% | 438 | 20.5% | 547 | 15.5% | 573 | 109 | 26 |
| Retrovirus | 19.7% | 370 | 17.9% | 478 | 14.6% | 538 | 108 | 60 |
| Plasmid DNA | 18.3% | 345 | 16.6% | 442 | 13.1% | 483 | 97 | 41 |
| Lentivirus | 2.9% | 55 | 7.3% | 196 | 9.9% | 364 | 141 | 168 |
| AAV | 4.9% | 92 | 7.6% | 204 | 9.5% | 350 | 112 | 146 |
| Nondividing Cell | Cargo Limit | Duration of Gene Expression | Physical Containment | Immunogenicity of Vectors | Drawbacks | Safety | |
|---|---|---|---|---|---|---|---|
| Adenovirus | Yes | 7–36 kb | Transient * | P2 | High | Strong cytotoxicity | Moderate |
| Retrovirus | No | 8–9 kb | Stable | P2 | Moderate | Risk of the carcinogenesis | Moderate |
| Plasmid DNA | Yes | limitless | Transient | - | - | Low transduction efficiency | High |
| Lentivirus | Yes | 8–9 kb | Stable | P2 | Moderate | Concerns about genes derived from HIV-1 | High |
| AAV | Yes | 4.7 kb | Potentially Stable | P1 | Low | Difficult to purification, Small cargo limit | High |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsunaga, W.; Gotoh, A. Adenovirus as a Vector and Oncolytic Virus. Curr. Issues Mol. Biol. 2023, 45, 4826-4840. https://doi.org/10.3390/cimb45060307
Matsunaga W, Gotoh A. Adenovirus as a Vector and Oncolytic Virus. Current Issues in Molecular Biology. 2023; 45(6):4826-4840. https://doi.org/10.3390/cimb45060307
Chicago/Turabian StyleMatsunaga, Wataru, and Akinobu Gotoh. 2023. "Adenovirus as a Vector and Oncolytic Virus" Current Issues in Molecular Biology 45, no. 6: 4826-4840. https://doi.org/10.3390/cimb45060307
APA StyleMatsunaga, W., & Gotoh, A. (2023). Adenovirus as a Vector and Oncolytic Virus. Current Issues in Molecular Biology, 45(6), 4826-4840. https://doi.org/10.3390/cimb45060307