
Etiology of Idiopathic Macular Holes in the Light of Estrogen Hormone



Abstract
:1. Introduction
2. Biochemistry
2.1. Retina and Estrogen
2.2. Protective Effect of Estrogen in the Retina
2.2.1. Energy Homeostasis
2.2.2. Extracellular Matrix
2.2.3. Antioxidant Influence
2.2.4. Estrogen Treatment in Eye Disease
2.3. Influence of Estrogen on Mitochondria and Cone Cells of the Retina
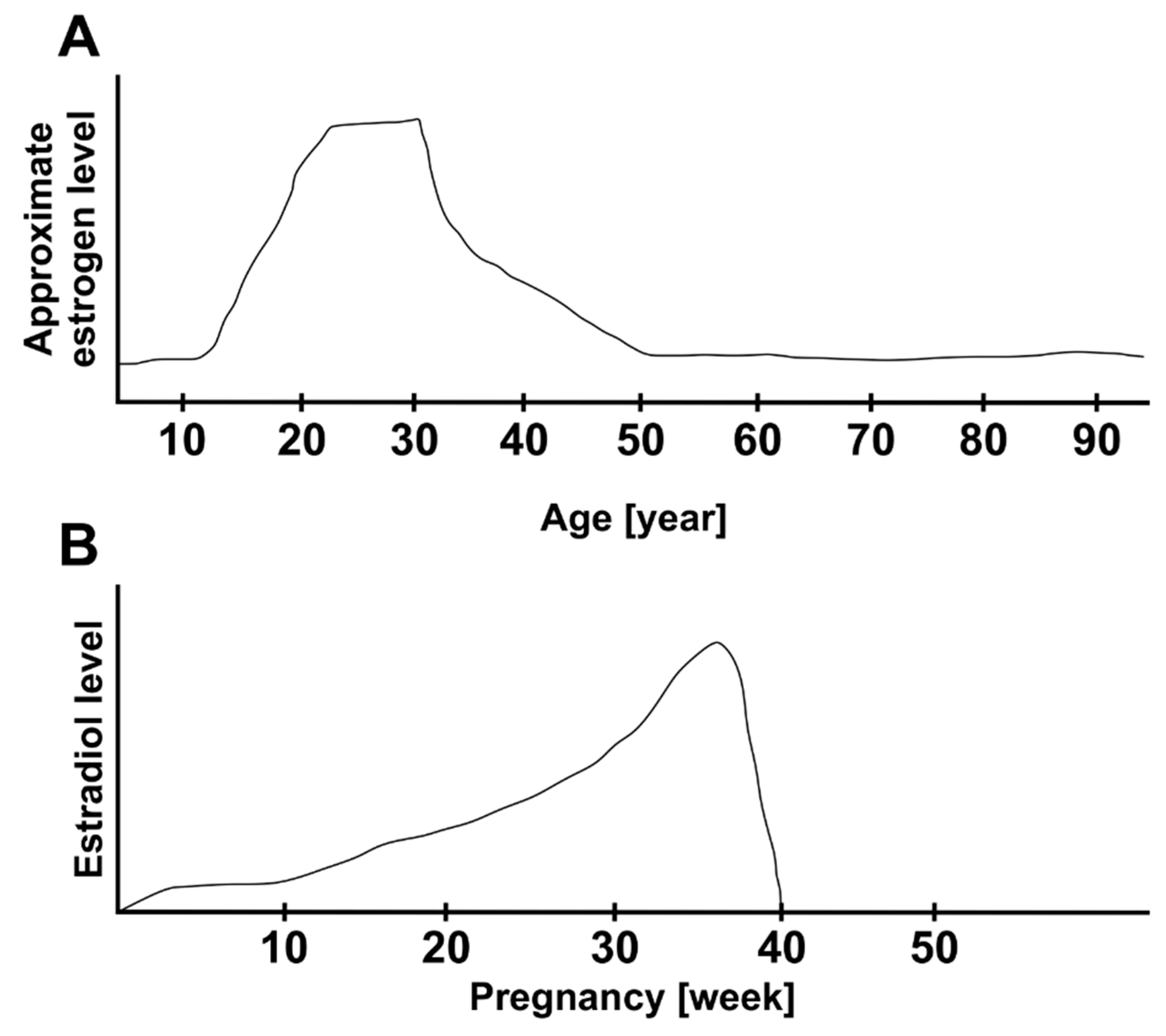
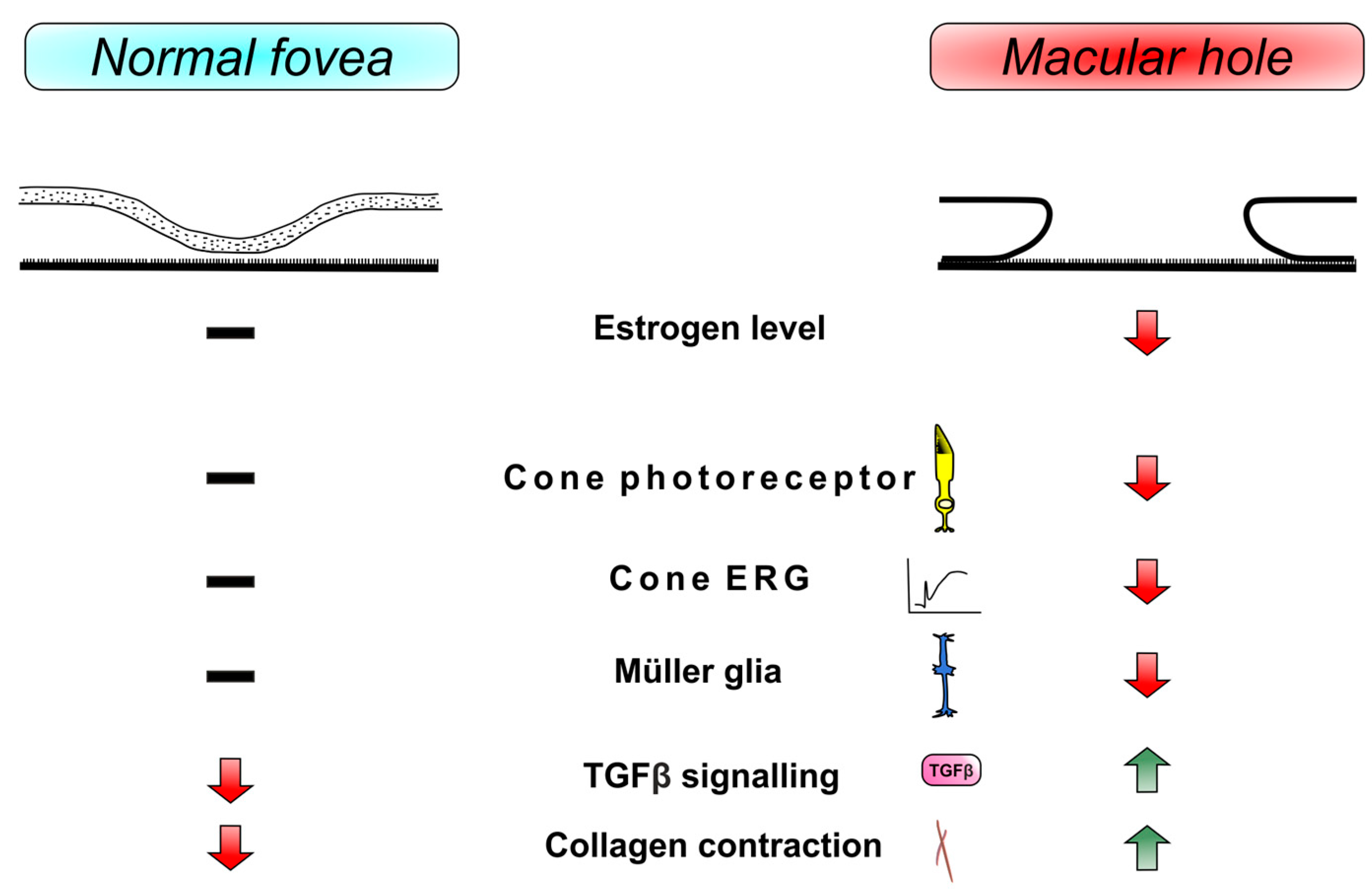
2.4. Role of Estrogen in Idiopathic Macula Hole
3. Anatomical and Morphological Effects of Estrogen on the Fovea
3.1. Müller Glia and Cone Cells in the Theory of Gass
3.2. Morphological Changes in the Retina Induced via Treatment with Tamoxifen
4. Functional Examination
4.1. Postive Effect of Estrogen on Vision
4.2. The Role of the Cone Cells in the Development of Idiopathic Macula Holes
5. New Epidemic Study
6. Discussion
7. Summary
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCannel, C.A.; Ensminger, J.L.; Diehl, N.N.; Hodge, D.N. Population-based incidence of macular holes. Ophthalmology 2009, 116, 1366–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.C.; Frimpong-Ansah, K.N. Idiopathic macular hole in a child. Eye 2012, 26, 620–621. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.S.; Fernandez-Sanz, G.; Levasseur, S.; Grigg, J.R.; Hunyor, A.P. Idiopathic full thickness macular hole in a 10-year-old girl. Int. J. Retin. Vitr. 2018, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Schwartz, S.D.; McHugh, J.D.; Thamby-Rajah, Y.; Hodgson, S.A.; Wormald, R.P.; Gregor, Z.J. Systemic risk factors for idiopathic macular holes: A case-control study. Eye 1998, 12, 256–259. [Google Scholar] [CrossRef]
- Weisz, J.; Ward, I.L. Plasma testosterone and progesterone titres of pregnant rats, their male and female fetuses, and neonatal offspring. Endocrinology 1980, 106, 306–316. [Google Scholar] [CrossRef]
- Du, M.; Mangold, C.A.; Bixler, G.V.; Brucklacher, R.M.; Masser, D.R.; Stout, M.B.; Elliott, M.H.; Freeman, W.M. Retinal gene expression responses to aging are sexually divergent. Mol. Vis. 2017, 23, 707–717. [Google Scholar]
- Chaychi, S.; Polosa, A.; Lachapelle, P. Differences in retinal structure and function between aging male and female Sprague-Dawley rats are strongly influenced by the Estrus cycle. PLoS ONE 2015, 10, e0136056. [Google Scholar] [CrossRef] [Green Version]
- Nixon, E.; Simpkins, J.W. Neuroprotective effects of nonfeminizing oestrogens in retinal photoreceptor neurons. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4739–4747. [Google Scholar] [CrossRef] [Green Version]
- Kaja, S.; Yang, S.H.; Wei, J.; Fujitani, K.; Liu, R.; Brun-Zinkernagel, A.M.; Simpkins, J.W.; Inokuchi, K.; Koulen, P. Estrogen protects the inner retina from apoptosis and ischemia-induced loss of Vesl-1L/Homer 1c immunoreactive synaptic connections. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3155–3162. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, T.; Takahashi, H.; Shimura, M. Estrogen has a neuroprotective effect on axotomized RGCs through ERK signal transduction pathway. Brain Res. 2006, 1093, 141–149. [Google Scholar] [CrossRef]
- The Eye Disease Case-Control Study Group. Risk factors for idiopathic macular holes. Am. J. Ophthalmol. 1994, 118, 754–761. [Google Scholar] [CrossRef]
- Maruichi, M.; Oku, H.; Takai, S.; Muramatsu, M.; Sugiyama, T.; Imamura, Y.; Minami, M.; Ueki, M.; Satoh, B.; Sakaguchi, M.; et al. Measurement of activities in two different angiotensin II generating systems, chymase and angiotensin-converting enzyme, in the vitreous fluid of vitreoretinal diseases: A possible involvement of chymase in the pathogenesis of macular hole patients. Curr. Eye Res. 2004, 29, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Katsumura, K.; Nakamura, K.; Kobayashi, M.; Muramatsu, M.; Maruichi, M.; Oku, H.; Takai, S.; Miyazaki, M.; Ikeda, T. Effects of chymase on the macular region in monkeys and porcine Muller cells: Probable involvement of chymase in the onset of idiopathic macular holes. Ophthalmic Res. 2006, 38, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Inokuchi, N.; Ikeda, T.; Nakamura, K.; Morishita, S.; Fukumoto, M.; Kida, T.; Oku, H. Vitreous estrogen levels in patients with an idiopathic macular hole. Clin. Ophthalmol. 2015, 9, 549–552. [Google Scholar] [PubMed] [Green Version]
- Shah, S.P.; Bunce, C.; Johnston, R.L.; Laidlaw, D.A. Are biometric parameters a risk factor for idiopathic macular hole formation? Results of a matched case-control series. Br. J. Ophthalmol. 2006, 90, 117–118. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Kobayashi, K.; Okinami, S. Macular hole and myopic refraction. Br. J. Ophthalmol. 2002, 86, 1269–1273. [Google Scholar] [CrossRef]
- Ghadiali, Q.; Zahid, S.; Dolz-Marco, R.; Tan, A.; Engelbert, M. An assessment of vitreous degeneration in eyes with vitreomacular traction and macular holes. J. Ophthalmol. 2017, 2017, 6834692. [Google Scholar] [CrossRef] [Green Version]
- Gass, J.D. Idiopathic senile macular Hole. Its early stages and pathogenesis. Arch. Ophthalmol. 1988, 106, 629–639. [Google Scholar] [CrossRef]
- Kishi, S.; Hagimura, N.; Shimizu, K. The role of the premacular liquefied pocket and premacular vitreous cortex in idiopathic macular hole development. Am. J. Ophthalmol. 1996, 122, 622–628. [Google Scholar] [CrossRef]
- Johnson, M.W.; Van Newkirk, M.R.; Meyer, K.A. Perifoveal vitreous detachment is the primary pathogenic event in idiopathic macular hole formation. Arch. Ophthalmol. 2001, 119, 215–222. [Google Scholar]
- Tornambe, P.E. Macular hole genesis: The hydration theory. Retina 2003, 23, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Theodossiadis, G.; Petrou, P.; Eleftheriadou, M.; Moustakas, A.L.; Datseris, I.; Theodossiadis, P. Focal vitreomacular traction: A prospective study of the evolution to macular hole: The mathematical approach. Eye 2014, 28, 1452–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Zhu, M.; Zhao, Y.; Qian, J.; Dufresne, C.; Turner, R.; Semba, R.D.; Solomon, S.D. A proteomic approach to understanding the pathogenesis of idiopathic macular hole formation. Clin. Proteom. 2017, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Kobayashi, H.; Ueda, M.; Honda, Y. Estrogen receptor expression in bovine and rat retinas. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2105–2110. [Google Scholar]
- Munaut, C.; Lambert, V.; Noël, A.; Frankenne, F.; Deprez, M.; Foidart, J.M.; Rakic, J.M. Presence of oestrogen receptor type beta in human retina. Br. J. Ophthalmol. 2001, 85, 877–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogueta, S.B.; Schwartz, S.D.; Yamashita, C.K.; Farber, D.B. Estrogen receptor in the human eye: Influence of gender and age on gene expression. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1906–1911. [Google Scholar]
- Hutchinson, C.V.; Walker, J.A.; Davidson, C. Oestrogen, ocular function and low-level vision: A review. J. Endocrinol. 2014, 223, R9–R18. [Google Scholar] [CrossRef]
- Gooren, L.J.; Toorians, A.W. Significance of oestrogens in male (patho)physiology. Ann. Endocrinol. 2003, 64, 126–135. [Google Scholar]
- Tsuchiya, Y.; Nakajima, M.; Yokoi, T. Cytochrome P450-mediated metabolism of estrogens and its regulation in human. Cancer Lett. 2005, 227, 115–124. [Google Scholar] [CrossRef]
- Giguère, V. Transcriptional control of energy homeostasis by the estrogen-related receptors. Endocr. Rev. 2008, 29, 677–696. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, S.N.; Emter, R.; Hock, M.B.; Knutti, D.; Cardenas, J.; Podvinec, M.; Oakeley, E.J.; Kralli, A. The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 6472–6477. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, J.; Tarttelin, E.E.; Gregory-Evans, C.Y.; Moosajee, M.; Gregory-Evans, K. Transcriptional regulation and expression of the dominant drusen gene FBLN3 (EFEMP1) in mammalian retina. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4613–4621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Wang, S.; Wang, B.; Du, F.; Hu, C.; Li, H.; Feng, Y.; Zhu, R.; Mo, M.; Cao, Y.; et al. 17β-Estradiol up-regulates Nrf2 via PI3K/AKT and estrogen receptor signaling pathways to suppress light-induced degeneration in rat retina. Neuroscience 2015, 304, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Ström, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [Green Version]
- Giddabasappa, A.; Bauler, M.; Yepuru, M.; Chaum, E.; Dalton, J.T.; Eswaraka, J. 17-β estradiol protects ARPE-19 cells from oxidative stress through oestrogen receptor-β. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5278–5287. [Google Scholar] [CrossRef] [Green Version]
- Pisano, A.; Preziuso, C.; Iommarini, L.; Perli, E.; Grazioli, P.; Campese, A.F.; Maresca, A.; Montopoli, M.; Masuelli, L.; Sadun, A.A.; et al. Targeting estrogen receptor β as preventive therapeutic strategy for Leber’s hereditary optic neuropathy. Hum. Mol. Genet. 2015, 24, 6921–6931. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Cavaliere, F.; Watanabe, C.; Nucci, C.; Bagetta, G.; Corasaniti, M.T.; Sakurada, S.; Morrone, L.A. 17Beta-estradiol prevents retinal ganglion cell loss induced by acute rise of intraocular pressure in rat. Prog. Brain Res. 2008, 173, 583–590. [Google Scholar]
- Prokai-Tatrai, K.; Xin, H.; Nguyen, V.; Szarka, S.; Blazics, B.; Prokai, L.; Koulen, P. 17β-estradiol eye drops protect the retinal ganglion cell layer and preserve visual function in an in vivo model of glaucoma. Mol. Pharm. 2013, 10, 3253–3261. [Google Scholar] [CrossRef] [Green Version]
- Honig, M.G.; Del Mar, N.A.; Henderson, D.L.; O’Neal, D.; Doty, J.B.; Cox, R.; Li, C.; Perry, A.M.; Moore, B.M.; Reiner, A. Raloxifene Modulates Microglia and Rescues Visual Deficits and Pathology After Impact Traumatic Brain Injury. Front. Neurosci. 2021, 15, 701317. [Google Scholar] [CrossRef]
- Kimura, K.; Orita, T.; Fujitsu, Y.; Liu, Y.; Wakuta, M.; Morishige, N.; Suzuki, K.; Sonoda, K.H. Inhibition by female sex hormones of collagen gel contraction mediated by retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- Hao, M.; Li, Y.; Lin, W.; Xu, Q.; Shao, N.; Zhang, Y.; Kuang, H. Estrogen prevents high-glucose-induced damage of retinal ganglion cells via mitochondrial pathway. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 83–90. [Google Scholar] [CrossRef]
- Farber, D.B.; Theendakara, V.P.; Akhmedov, N.B.; Saghizadeh, M. ZBED4, a novel retinal protein expressed in cones and Müller cells. Adv. Exp. Med. Biol. 2010, 664, 79–87. [Google Scholar] [PubMed]
- Masuda, T.; Wada, Y.; Kawamura, S. ES1 is a mitochondrial enlarging factor contributing to form megamitochondria in zebrafish cones. Sci. Rep. 2016, 6, 22360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.H.; Chen, Z.Y.; Yin, L.L.; Zheng, Z.; Wu, X.W. Effects of estrogen on collagen gel contraction by human retinal glial cells. Chin. Med. J. 2012, 125, 4098–4103. [Google Scholar]
- James, M.; Feman, S.S. Macular holes. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 1980, 215, 59–63. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, P.J.; Fine, S.L.; Hillis, A.I. Clinical features of idiopathic macular cysts and holes. Am. J. Ophthalmol. 1982, 93, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, Y.; Morishita, S.; Horie, T.; Fukumoto, M.; Sato, T.; Kida, T.; Oku, H.; Sugasawa, J.; Ikeda, T.; Nakamura, K. A comparison of sex steroid concentration levels in the vitreous and serum of patients with vitreoretinal diseases. PLoS ONE 2017, 12, e0180933. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.E.; Kim, S.W.; Chung, H.W.; Kang, S.W. Estrogen antagonist and development of macular hole. Korean J. Ophthalmol. 2010, 24, 306–309. [Google Scholar] [CrossRef] [Green Version]
- Cronin, B.G.; Lekich, C.K.; Bourke, R.D. Tamoxifen therapy conveys increased risk of developing a macular hole. Int. Ophthalmol. 2005, 26, 101–105. [Google Scholar] [CrossRef]
- Torrell Belzach, N.; Vela Segarra, J.I.; Crespí Vilimelis, J.; Alhayek, M. Bilateral Macular Hole Related to Tamoxifen Low-Dose Toxicity. Rep. Ophthalmol. 2020, 11, 528–533. [Google Scholar] [CrossRef]
- Goldman, D. Müller glial cell reprogramming and retina regeneration. Nat. Rev. Neurosci. 2014, 15, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäenpää, H.; Mannerström, M.; Toimela, T.; Salminen, L.; Saransaari, P.; Tähti, H. Glutamate uptake is inhibited by tamoxifen and toremifene in cultured retinal pigment epithelial cells. Pharmacol. Toxicol. 2002, 91, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, A.; Saito, W.; Hashimoto, Y.; Ishida, S. Improvements in visual acuity and macular morphology following cessation of anti-estrogen drugs in a patient with anti-estrogen maculopathy resembling macular telangiectasia type 2: A pathogenic hypothesis. BMC Ophthalmol. 2019, 19, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, E. Some structural features of the fovea centralis in the human retina. Arch. Ophthalmol. 1969, 82, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Gass, J.D. Müller cell cone, an overlooked part of the anatomy of the fovea centralis: Hypotheses concerning its role in the pathogenesis of macular hole and foveomacular retinoschisis. Arch. Ophthalmol. 1999, 117, 821–823. [Google Scholar] [CrossRef] [Green Version]
- Ezra, E. Idiopathic full thickness macular hole: Natural history and pathogenesis. Br. J. Ophthalmol. 2001, 85, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Baget-Bernaldiz, M.; Soler Lluis, N.; Romero-Aroca, P.; Traveset-Maeso, A. Optical coherence tomography study in tamoxifen maculopathy. Arch. Soc. Esp. Oftalmol. 2008, 83, 615–618. [Google Scholar] [PubMed]
- Bicer, T.; Imamoglu, G.I.; Dogan, A.S.; Avarisli, N.A.; Kabatas, N.; Bicer, B.K.; Gurdal, C. The effects of adjuvant hormonotherapy on tear functions in patients with breast cancer. Int. Ophthalmol. 2020, 40, 2077–2083. [Google Scholar] [CrossRef]
- Gualino, V.; Cohen, S.Y.; Delyfer, M.N.; Sahel, J.A.; Gaudric, A. Optical coherence tomography findings in tamoxifen retinopathy. Am. J. Ophthalmol. 2005, 140, 757–758. [Google Scholar] [CrossRef]
- Wagner-Schuman, M.; Dubis, A.M.; Nordgren, R.N.; Lei, Y.; Odell, D.; Chiao, H.; Weh, E.; Fischer, W.; Sulai, Y.; Dubra, A.; et al. Race- and sex-related differences in retinal thickness and foveal pit morphology. Investig. Ophthalmol. Vis. Sci. 2011, 52, 625–634. [Google Scholar] [CrossRef]
- Bafiq, R.; Mathew, R.; Pearce, E.; Abdel-Hey, A.; Richardson, M.; Bailey, T.; Sivaprasad, S. Age, sex, and ethnic variations in inner and outer retinal and choroidal thickness on spectral-domain optical coherence tomography. Am. J. Ophthalmol. 2015, 160, 1034–1043.e1. [Google Scholar] [CrossRef] [PubMed]
- Eisner, A.; Burke, S.N.; Toomey, M.D. Visual sensitivity across the menstrual cycle. Vis. Neurosci. 2004, 21, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, G.Y.; Bearse MAJr Harrison, W.W.; Bronson-Castain, K.W.; Schneck, M.E.; Barez, S.; Adams, A.J. Differences in neuroretinal function between adult males and females. Optom. Vis. Sci. 2014, 91, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, H.; Erkin, E.F.; Mavioğlu, H.; Sungurtekin, U. Changes in pattern reversal evoked potentials during menstrual cycle. Int. Ophthalmol. 1998, 22, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Okawa, H.; Sampath, A.P.; Laughlin, S.B.; Fain, G.L. ATP consumption by mammalian rod photoreceptors in darkness and in light. Curr. Biol. 2008, 18, 1917–1921. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, S.; Ghosh, F. Cone implicit time as a predictor of visual outcome in macular hole surgery. Graefes Arch. Clin. Exp. Ophthalmol. 2014, 252, 1903–1909. [Google Scholar] [CrossRef] [PubMed]
- Ştefănescu-Dima, A.Ş.; Corîci, C.A.; Mănescu, M.R.; Sas, T.N.; Iancău, M.; Mocanu, C.L. Posterior vitreous detachment and macular anatomical changes—A tomographic-electroretinographic study. Rom. J. Morphol. Embryol. 2016, 57, 751–758. [Google Scholar]
- Karatepe, A.S.; Menteş, J.; Erakgün, E.T.; Afrashi, F.; Nalçacı, S.; Akkın, C.; Ateş, Y. Vitreoretinal interface characteristics in eyes with idiopathic macular holes: Qualitative and quantitative analysis. Turk. J. Ophthalmol. 2018, 48, 70–74. [Google Scholar] [CrossRef]
- Hwang, S.; Kang, S.W.; Kim, S.J.; Choi, J.; Son, K.Y.; Lim, D.H.; Shin, D.W.; Choi, D.; Chang, Y.; Ryu, S.; et al. Risk factors for the development of idiopathic macular hole: A nationwide population-based cohort study. Sci. Rep. 2022, 12, 21778. [Google Scholar] [CrossRef]
- Wulf, K.-H.; Schmidt-Matthiesen, H. Schwangerschaft I, 3rd ed.; Urban&Schwarzenberg: München, Germany, 1992. [Google Scholar]
- Wulf, K.-H.; Schmidt-Matthiesen, H. Gynäkologische Endokrinologie, Band I, 2nd ed; Urban&Schwarzenberg: München, Germany, 1987. [Google Scholar]
- Kunst, S.; Wolloscheck, T.; Grether, M.; Trunsch, P.; Wolfrum, U.; Spessert, R. Photoreceptor cells display a daily rhythm in the orphan receptor Esrrβ. Mol. Vis. 2015, 21, 173–184. [Google Scholar]



{kind=link}
{kind=link}
| Study | Species | Relevant Results | Category |
|---|---|---|---|
| Zhu C et al., 2015 | Rats | Protective mechanism of βE2 through antioxidative effects through NRF2 activation | Positive effect of estrogen on retina |
| Giddabasappa A et al., 2010 | Human | 17β-E2-mediated cytoprotection occurred through the preservation of mitochondrial function, reduction in ROS production, induction of cellular antioxidant genes | |
| Honig MG et al., 2021 | Mice | Estrogen reduced the visual system damage and rescued photoreceptor cells after ocular blast in mice | |
| Farber DB et al., 2010 | Human | ZBED4, a novel protein localized in cone cell and glial Müller cells, several putative ZBED4-interacting proteins including a co-repressor of the ORα | |
| Masuda T et al., 2016 | Zebra fish | ES1, a novel mitochondria-enlarging factor localized in cone mitochondria, contributes to the formation of mega-mitochondria in cone cells | |
| The Eye Disease Case-Control Study Group, 1994 | Human | Positive effect of estrogen against IMH | Epidemic study |
| Sungsoon H et al., 2022 | Human | IMH occurs earlier in woman than men, postmenopausal women with two or more children showed a greater risk for IMH than those who had not been pregnant | |
| Kimura K et al., 2014 | Human | Positive effect of estrogen on RPE cells, estrogen appears to inhibit TGF-β2-induced collagen contraction | Collagen contraction study |
| Qiu QH et al., 2012 | Human | Estrogen can inhibit collagen gel contraction, which is caused by cultured human retinal glial cells | |
| James M et al., 1980 | Human | Development of macular holes under estrogen treatment | Controverse study |
| McDonnell PJ et al., 1982 | Human | Development of macular holes under estrogen treatment | |
| Inokuchi N et al., 2015 | Human | Vitreous E2 level is higher than the E1 level in IMH patients | |
| Nishikawa Y et al., 2017 | Human | Production of E2 by astrocytes and Müller cells may be responsible for the suspected local production of E2 | |
| Chung SE et al., 2010 | Human | Relationship between antiestrogen treatment and IMH development | Antiestrogen study |
| Cronin BG et al., 2005 | Human | Relationship between antiestrogen treatment and IMH development | |
| Torrell Belzach N et al., 2020 | Human | Relationship between antiestrogen treatment and IMH development | |
| Mäenpää H et al., 2002 | Human | Tamoxifen: dose-dependent decrease in the glutamate uptake of RPE cells, which has been noted in porcine RPE cells as well as in human RPE cell lines | |
| Yamada E et al., 1969 | Human | Structure of fovea in regard to Müller glia and cone cells | Fovea structure |
| Gass JD, 1999 | Human | Important hypothesis, suggesting that Müller cell cones bind the receptor cells in the fovea together | |
| Ezra E, 2001 | Human | Anatomical association between cones and IMH | |
| Yilmaz H et al., 1998 | Human | Pattern reversal evoked potentials (PRVEP) latency during the ovulatory phase was significantly shorter than that of other phases | Retina function |
| Andréasson S et al., 2014 | Human | Cone IT in full-field 30 Hz flicker ERG has been suggested as a predictor of visual outcome after surgery for IMH | |
| Ştefănescu-Dima AŞ, 2016 | Human | Response of cone cells determined through full-field ERG was proposed as a marker of retinal damage in macular pathology |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wergenthaler, N.; Dick, H.B.; Tsai, T.; Joachim, S.C. Etiology of Idiopathic Macular Holes in the Light of Estrogen Hormone. Curr. Issues Mol. Biol. 2023, 45, 6339-6351. https://doi.org/10.3390/cimb45080400
Wergenthaler N, Dick HB, Tsai T, Joachim SC. Etiology of Idiopathic Macular Holes in the Light of Estrogen Hormone. Current Issues in Molecular Biology. 2023; 45(8):6339-6351. https://doi.org/10.3390/cimb45080400
Chicago/Turabian StyleWergenthaler, Nousal, H. Burkhard Dick, Teresa Tsai, and Stephanie C. Joachim. 2023. "Etiology of Idiopathic Macular Holes in the Light of Estrogen Hormone" Current Issues in Molecular Biology 45, no. 8: 6339-6351. https://doi.org/10.3390/cimb45080400
APA StyleWergenthaler, N., Dick, H. B., Tsai, T., & Joachim, S. C. (2023). Etiology of Idiopathic Macular Holes in the Light of Estrogen Hormone. Current Issues in Molecular Biology, 45(8), 6339-6351. https://doi.org/10.3390/cimb45080400